Draft genome sequence of the extremely halophilic archaeon Haladaptatus cibarius type strain D43T isolated from fermented seafood
Hae-Won Lee, Dae-Won Kim, Mi-Hwa Lee, Byung-Yong Kim, Yong-Joon Cho, Kyung June Yim, Hye Seon Song, Jin-Kyu Rhee, Myung-Ji Seo, Hak-Jong Choi, Jong-Soon Choi, Dong-Gi Lee, Changmann Yoon, Young-Do Nam, Seong Woon Roh

TL;DR
This paper presents the draft genome sequence of a salt-loving archaeon, Haladaptatus cibarius, isolated from fermented seafood.
Contribution
The study provides the first annotated genome sequence of Haladaptatus cibarius, contributing to understanding halophilic archaea.
Findings
The genome is 3,926,724 bp long with 4,092 protein-coding genes and 57 RNA genes.
The average G + C content of the genome is 57.76%.
The strain shows 98.7% 16S rRNA gene sequence similarity with Haladaptatus litoreus.
Abstract
An extremely halophilic archaeon, Haladaptatus cibarius D43T, was isolated from traditional Korean salt-rich fermented seafood. Strain D43T shows the highest 16S rRNA gene sequence similarity (98.7 %) with Haladaptatus litoreus RO1-28T, is Gram-negative staining, motile, and extremely halophilic. Despite potential industrial applications of extremely halophilic archaea, their genome characteristics remain obscure. Here, we describe the whole genome sequence and annotated features of strain D43T. The 3,926,724 bp genome includes 4,092 protein-coding and 57 RNA genes (including 6 rRNA and 49 tRNA genes) with an average G + C content of 57.76 %.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
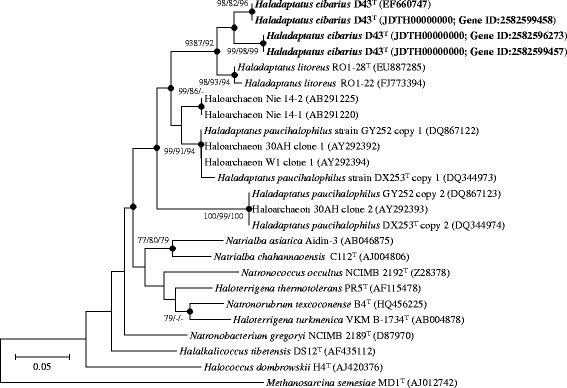 Figure 1
Figure 1 Figure 2
Figure 2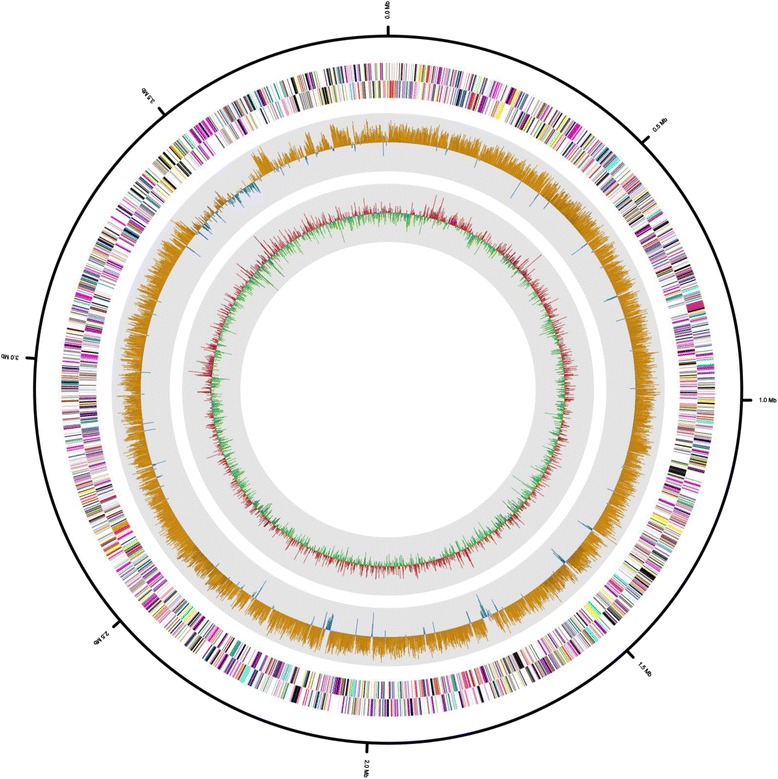 Figure 3
Figure 3Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsDevelopmental and Educational Neuropsychology · Educational theories and practices
Introduction
The extremely halophilic archaea, called haloarchaea, possess the small retinal protein halorhodopsin [1–3] and currently consists of more than 47 genera that live in hypersaline environments [4, 5]. Three members of the genus Haladaptatus—H. paucihalophilus [6], H. litoreus [7], and H. cibarius [8]—were isolated from a low-salt, sulfide-rich spring; marine solar saltern; and salt-fermented seafood, respectively. Haladaptatus comprises Gram-negative staining, non-motile haloarchaea that have polar lipids including phosphatidylglycerol, phosphatidylglycerol phosphate methyl ester, and phosphatidylglycerol sulfate [6]. The genomic analysis revealed that H. paucihalophilus survives in low salinity conditions because of trehalose synthesis with OtsAB pathway and trehalose glycosyl-transferring synthase pathway, and glycine betaine uptake [9]. However, other members in the genus Haladaptatus have not been analyzed at the genome level.
H. cibarius was isolated from the traditional Korean salt-fermented seafood, which is made with shellfish [8]. D43^T^ (= DSM 19505^T^ = JCM 15962^T^) is a representative strain and designated as the type strain of the species. It can grow in 10%–30% (w/v) NaCl (optimum, 15%), with Mg^2+^ required for growth. In addition, cells are not lysed in distilled water. The genome sequences of this genus are expected to provide fundamental information for the halotolerant features and biotechnological applications of the haloarchaea. Here, we describe the first whole genome sequence of H. cibarius along with its annotated features, and summarize the taxonomic classification.
Organism information
Classification and features
The taxonomic position for H. cibarius D43^T^ was identified with type strains obtained from the EzTaxon-e server [10]. The 16S rRNA sequences of D43^T^ and closely related strains were aligned using the ClustalW multiple sequence alignment program [11] and were subsequently used for the phylogenetic analysis. Phylogenetic trees were constructed using the neighbor-joining [12], maximum-parsimony [13], and maximum likelihood [14] algorithms with bootstrap values of 1,000 using MEGA version 5 molecular evolutionary genetics analysis program [15]. Strain D43^T^ clustered with type strains of Haladaptatus species (Fig. 1), exhibiting 16S rRNA gene sequence similarities of 98.7% and 95.1% between strain D43^T^ (EF660747) and the type strain of H. litoreus and H. paucihalophilus, respectively. Classification and general features of H. cibarius D43^T^ are shown in Table 1.Fig. 1. Phylogenetic tree constructed using the neighbor-joining method based on 16S rRNA gene sequences, showing the taxonomic position of strain D43^T^ in genus Haladaptatus. Bootstrap values (>70%) at nodes are shown as percentages calculated using the neighbor-joining/maximum parsimony/maximum likelihood probabilities based on 1000 replicates. Filled circles indicate identical branches generated using three algorithms. Methanosarcina semesiae MD1^T^ was used as an outgroup. Bar, 0.05 substitutions per nucleotide positionTable 1Classification and general features of Haladaptatus cibarius D43^T^ [18]MIGS IDPropertyTermEvidence code^a^ ClassificationDomain Archaea TAS [24]Phylum Euryarchaeota TAS [25]Class Halobacteria TAS [26]Order Halobacteriales TAS [27, 28]Family Halobacteriaceae TAS [28, 29]Genus Haladaptatus TAS [6]Species Haladaptatus cibarius TAS [8]Type strain D43^T^ (DSM 19505, JCM 15962)TAS [8]Gram stainNegativeTAS [8]Cell shapecoccus or coccobacillusTAS [8]MotilitymotileTAS [8]SporulationNot reportedTAS [8]Temperature range15–50 °CTAS [8]Optimum temperature37 °CTAS [8]pH range; Optimum6.0–8.0; 7.0TAS [8]Carbon sourceSucrose, D-fructose, D-glucose, lactose, formate, acetateTAS [8]MIGS-6HabitatSalt-fermented seafoodTAS [8]MIGS-6.3Salinity35 % NaCl (w/v)TAS [8]MIGS-22Oxygen requirementAerobicTAS [8]MIGS-15Biotic relationshipFree-livingTAS [8]MIGS-14PathogenicityNot reportedMIGS-23.1IsolationSalt-fermented foodTAS [8]MIGS-4Geographic locationRepublic of KoreaTAS [8]MIGS-5Sample collection timeNot reportedMIGS-4.1LatitudeNot reportedMIGS-4.2LongitudeNot reportedMIGS-4.3DepthNot reportedMIGS-4.4AltitudeNot reported ^a^Evidence codes - TAS: traceable author statement (i.e., a direct report exists in the literature). These evidence codes are from the Gene Ontology project [30]
Strain D43^T^ is a Gram-negative staining, coccus or coccobacillus, motile archaeon approximately 1.0 μm in diameter (Fig. 2). Catalase and oxidase tests yielded positive results, but reduction of nitrate to nitrite under aerobic conditions was negative. Cells contained the polar lipids phosphatidylglycerol, phosphatidylglycerol phosphate methyl ester, and two unidentified glycolipids. Strain D43^T^ hydrolyzed gelatin and Tween 80, utilized formate and acetate as carbon sources, and produced acid from sucrose and d-glucose. The strain was sensitive to anisomycin, aphidicolin, chloramphenicol, and rifampicin, and was resistant to ampicillin, erythromycin, kanamycin, streptomycin, and polymycin B.Fig. 2. Scanning electron micrographs of H. cibarius D43^T^ obtained by SUPRA 55VP (Carl Zeiss, Jena, Germany). Scale bars represent 200 nm
Genome sequencing and annotation
Genome project history
The genome project and sequence of the H. cibarius D43^T^ genome were deposited in the Genomes OnLine Database [16] (project ID: Gp0086819) and GenBank (accession number: JDTH00000000), respectively. The BioProject number was PRJNA236630. Sequencing and annotation were performed by Chun Lab Inc. (Seoul, Korea) and Integrated Microbial Genomes Expert Review (IMG-ER) [17].
Growth conditions and genomic DNA preparation
H. cibarius D43^T^ grew optimally on halophilic medium [6] supplemented with 15% (w/v) NaCl and 20 mM Mg^2+^ adjusted to pH 7.0, producing colonies with a pink color after incubation at 37°C as previously described [8]. Genomic DNA was extracted and purified using a G-spin DNA extraction kit (iNtRON Biotechnology Inc., Sungnam, Korea), according to the manufacturer’s instructions.
Genome sequencing and assembly
Genomic sequences of H. cibarius D43^T^ were generated from a total of 9,237,360 quality-filtered reads (710.3-fold coverage) by combining 5,074,634 reads (374.9-fold coverage) obtained from Mi-Seq 300 bp paired-end library (Illumina, San Diego, CA, USA), 4,112,798 reads (292.1-fold coverage) obtained from an Ion Torrent Personal Genome Machine 318v2 chip (Life Technologies, Carlsbad, CA, USA), and 49,928 reads (43.3-fold coverage) obtained from PacBio RS 10 kb library (Pacific Biosciences, Menlo Park, USA). Illumina and PGM data were assembled de novo with CLC Genomics Workbench 6.5.1 (CLC bio, Boston, MA, USA) and PacBio data were assembled using the HGAP2 algorithm in SMRT Analysis 2.1 (Pacific Biosciences). Resultant contigs were assembled with CodonCode Aligner 3.7 (CodonCode Corporation, Centerville, MA, USA). Sequences were assembled to 13 scaffolds with an N50 contig size of 985,075 bp; the genome sequencing project information and its associated MIGS version 2.0 compliance levels [18] are shown in Table 2.Table 2. Project informationMIGS IDPropertyTermMIGS-31Finishing qualityImproved high-quality draftMIGS-28Libraries usedIllumina PE, Ion PGM, and PacBio librariesMIGS-29Sequencing platformsIllumina Mi-seq, Ion PGM, and PacBio RS systemsMIGS-31.2Fold coverage374.92 × Illumina; 292.08 × Ion PGM; 43.25 × PacBioMIGS-30AssemblersCLC Genomics Workbench 6.5.1, SMRT Analysis 2.1MIGS-32Gene calling methodIMG-ERLocus TagHL45GenBank IDJDTH0000000GenBank Date of ReleaseJune 20, 2014GOLD IDGi0069860BIOPROJECTPRJNA236630MIGS-13Source material identifierD43^T^ Project relevanceEnvironmental and biotechnological
Genome annotation
The open reading frames of the assembled genome were predicted and annotated using IMG-ER [17], NCBI COG [19], Pfam [20], and EzTaxon-e [10] databases. The rRNA and tRNA genes were identified using RNAmmer 1.2 [21] and tRNA scan-SE 1.23 [22], respectively.
Genome properties
The draft genome sequence for H. cibarius D43^T^ contained 3,926,724 bp, with 13 scaffolds. The G + C content was 57.76 % (Fig. 3 and Table 3), and 4,092 protein-coding genes were predicted along with 57 RNA genes, including six rRNA (two 5S, three 16S, and one 23S rRNA), 49 tRNA, and two additional RNA genes. There were 2,676 protein-coding genes with predicted functions: 773 were enzymes, 98 encoded signal peptides, and 1,049 encoded transmembrane proteins. The distribution of genes in the COG functional categories is shown in Table 4. A large number of genes were associated with the COG functional categories of cell wall biogenesis (79, 3.3 %); transcription (100, 4.1 %); and transport and metabolism of amino acids (299, 12.3 %), carbohydrates (121, 5.0 %), and lipids (80, 3.3 %). Further analysis with dbCAN [23], a database for annotation of carbohydrate-active enzymes, showed that the genome contains genes encoding various enzymes for the breakdown and biosynthesis of carbohydrates such as chitinase (GH18), chitosanase (GH5), pullulanase (GH13), trehalose synthase (GT4 and 20), cellulose synthase (GT2), and alginate lyase (PL6).Fig. 3. Graphical map of the H. cibarius D43^T^ pseudochromosome. From outside to center: RNA genes (red, tRNA and blue, rRNA) and genes on the antisense and sense strands (colored according to COG categories). Inner circle shows the GC skew, with yellow and blue indicating positive and negative values, respectively. GC content is indicated in red and green. The genome map was visualized using CLgenomics 1.06 (Chun Lab Inc.)Table 3. Genome statisticsAttributeValue% of TotalGenome size (bp)3,926,724100.00DNA coding (bp)3,378,68486.04DNA G + C (bp)2,267,91557.76DNA scaffolds13100.00Total genes4,149100.00Protein-coding genes4,09298.63RNA genes571.37Genes in internal clusters3,13575.56Genes with function prediction2,67664.50Genes assigned to COGs2,18852.74Genes assigned Pfam domains2,69965.05Genes with signal peptides982.36Genes with transmembrane helices104925.28CRISPR repeats4Table 4Number of genes associated with general COG functional categoriesCodeValue% ageDescriptionJ1646.76Translation, ribosomal structure, and biogenesisA10.04RNA processing and modificationK1004.12TranscriptionL1024.20Replication, recombination, and repairB30.12Chromatin structure and dynamicsD200.82Cell cycle control, cell division, chromosome partitioningY00.00Nuclear structureV371.53Defense mechanismsT552.27Signal transduction mechanismsM793.26Cell wall/membrane biogenesisN281.15Cell motilityZ00.00CytoskeletonW00.00Extracellular structuresU281.15Intracellular trafficking and secretion, and vesicular transportO883.63Post-translational modification, protein turnover, chaperonesC1626.68Energy production and conversionG1214.99Carbohydrate transport and metabolismE29912.32Amino acid transport and metabolismF763.13Nucleotide transport and metabolismH1094.49Coenzyme transport and metabolismI803.30Lipid transport and metabolismP1737.13Inorganic ion transport and metabolismQ461.90Secondary metabolism biosynthesis, transport, and catabolismR39216.16General function prediction onlyS26310.84Function unknown-196147.26Not in COGsThe total is based on the total number of protein coding genes in the genome
Insights from the genome sequence
The genome analysis of H. cibarius D43^T^ revealed genes involved in glycine betaine synthesis—including betaine aldehyde dehydrogenase, glycine betaine demethylase, and choline-glycine betaine transporter gene—that allow H. cibarius to maintain osmotic balance in hypersaline environments. In addtion, trehalose-related genes of trehalose-6-phosphate synthase, trehalose-6-phosphatase, trehalose-6-phosphate synthase and trehalose-6-phosphate hydrolase, and trehalose-utilization protein genes were analyzed in the genome sequences of H. cibarius D43^T^. The genes related with trehalose synthesis in the genome show the possibility of trehalose production that is important in food industry.
Conclusions
The draft genome sequences of the extremely halophilic archaeon isolated from the salt-fermented seafood were analyzed. Genes related with glycine betaine and trehalose for the survival in extreme environments were identified. The extremely halophilic archaeon could be a valuable resource for biotechnological applications because hypersaline conditions minimize the risk of contamination by other microorganisms. Further characterization of halophilic enzymes of the haloarchaea based on the genomic analyses can provide more detailed information on enzyme structures and potential industrial applications.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Lanyi JK Halorhodopsin: a light-driven chloride ion pump Annu Rev Biophys Biophys Chem 198615112810.1146/annurev.bb.15.060186.0003032424472 · doi ↗ · pubmed ↗
- 2Lanyi JK Halorhodopsin, a light-driven electrogenic chloride-transport system Physiol Rev 199070319330218149810.1152/physrev.1990.70.2.319 · doi ↗ · pubmed ↗
- 3Oesterhelt D Tittor J Bamberg EA unifying concept for ion translocation by retinal proteins J Bioenerg Biomembr 19922418119110.1007/BF 007626761526960 · doi ↗ · pubmed ↗
- 4Oren A Dworkin M Falkow S Rosenberg E Shleifer KH Stakebrandt E The order Halobacteriales The prokaryotes. Volume 320063 New York Springer 113164
- 5Oren A Ventosa A International Committee on Systematics of Prokaryotes Subcommittee on the taxonomy of Halobacteriaceae and Subcommittee on the taxonomy of Halomonadaceae: minutes of the joint open meeting, 31 July 2014, Montreal, Canada Int J Syst Evol Microbiol 2014643915391810.1099/ijs.0.069930-025386005 · doi ↗ · pubmed ↗
- 6Savage KN Krumholz LR Oren A Elshahed MS Haladaptatus paucihalophilus gen. nov., sp. nov., a halophilic archaeon isolated from a low-salt, sulfide-rich spring Int J Syst Evol Microbiol 200757192410.1099/ijs.0.64464-017220434 · doi ↗ · pubmed ↗
- 7Cui HL, Sun FF, Gao X, Dong Y, Xu XW, Zhou YG, et al. Haladaptatus litoreus sp. nov., an extremely halophilic archaeon from a marine solar saltern, and emended description of the genus Haladaptatus. Int J Syst Evol Microbiol. 2010;60:1085–9.10.1099/ijs.0.015933-019666806 · doi ↗ · pubmed ↗
- 8Roh SW Lee ML Bae JW Haladaptatus cibarius sp. nov., an extremely halophilic archaeon from seafood, and emended description of the genus Haladaptatus Int J Syst Evol Microbiol 2010601187119010.1099/ijs.0.013037-019667394 · doi ↗ · pubmed ↗