Analytical sequence to study G-CSF effect on the transcriptome of isolated spinal motoneurons from SOD1 G93A mice, an animal model for amyotrophic lateral sclerosis
Alexandre Henriques, Stefan Kastner, Oliver Wafzig, Jose-Luis Gonzalez De Aguilar, Armin Schneider

TL;DR
This study examines how G-CSF affects gene activity in spinal motor neurons of mice with a model of amyotrophic lateral sclerosis.
Contribution
The paper provides quality controls and details the experimental strategy for a microarray study on G-CSF's effects.
Findings
G-CSF re-adjusted gene expression in motoneurons of symptomatic SOD1G93A mice.
G-CSF modulates genes related to neuromuscular function in the studied animal model.
Abstract
Granulocyte-colony stimulating factor (G-CSF) has been recently identified as a neurotrophic factor able to preserve motor functions, rescue motor units and extent survival in an animal model of amyotrophic lateral sclerosis, the SOD1 G93A mice. To gain insight into the mode of action of G-CSF, we have recently performed gene expression profiling on isolated lumbar motoneurons from SOD1G93A mice, and shown that G-CSF re-adjusted gene expression in motoneurons of symptomatic SOD1G93A mice and modulates genes related to neuromuscular function (Henriques et al., 2015). Here, we provide quality controls for the microarray experiment (GO accession number GSE60856) and describe the experimental strategy.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
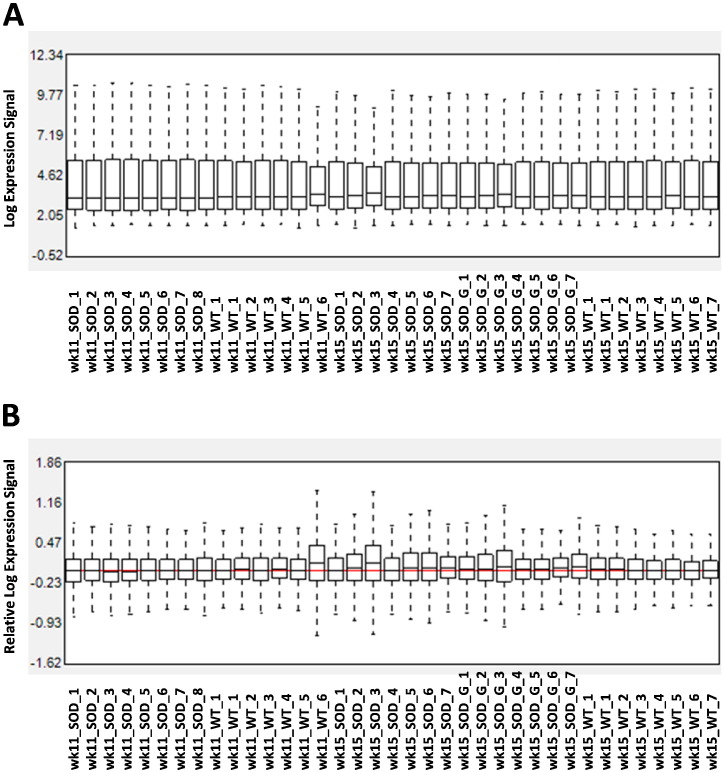 Figure 1
Figure 1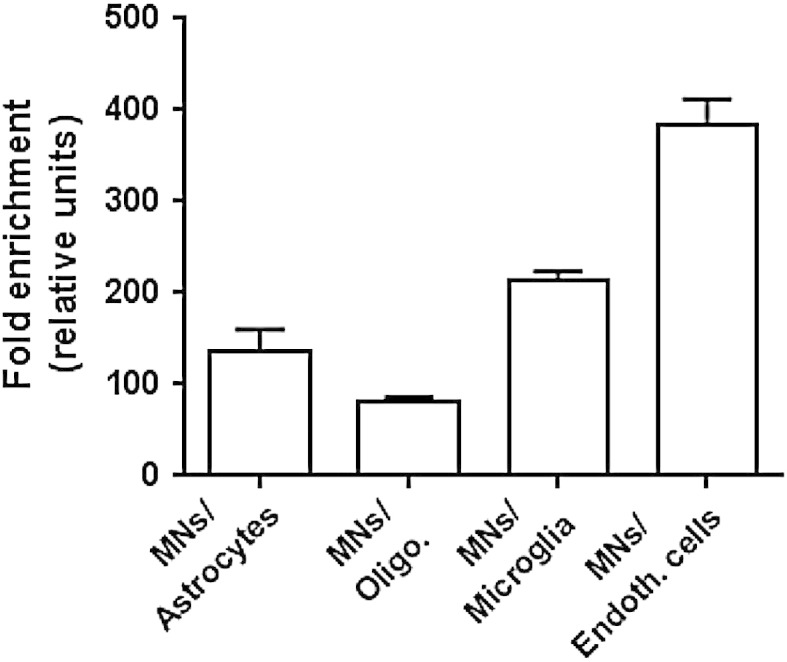 Figure 2
Figure 2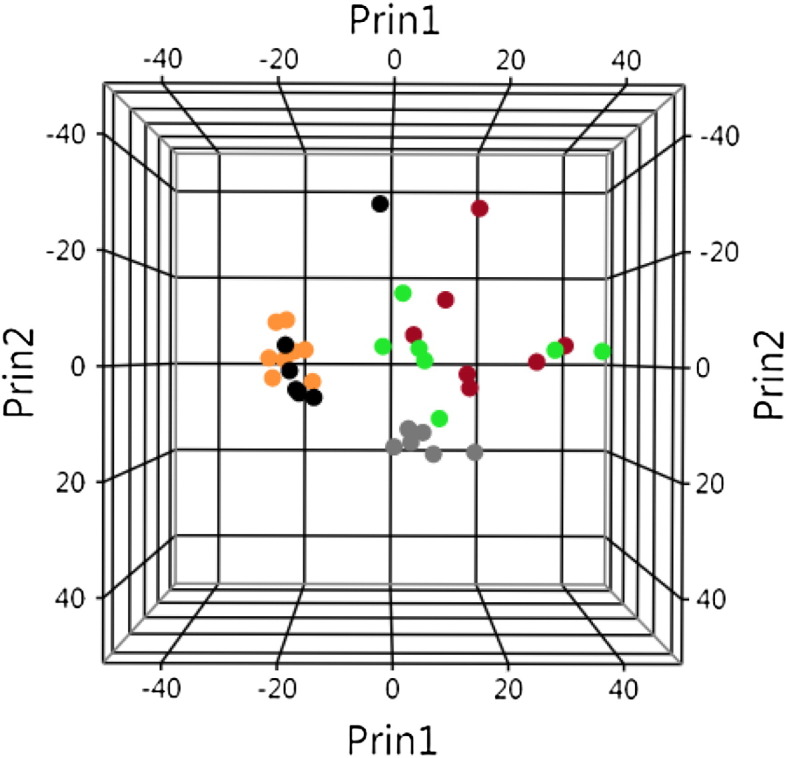 Figure 3
Figure 3Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAmyotrophic Lateral Sclerosis Research · Neurogenetic and Muscular Disorders Research · Muscle Physiology and Disorders
SpecificationsOrganism/cell line/tissueMus musculus, laser capture spinal motoneuronsSexFemalesSequencer or array typeAffymetrix, GeneChip® mouse genome 430 2.0 arrayData formatRaw data (cel files)Experimental factorsIsolated motoneurons from SOD1 G93A mice with or without pharmacological treatment based on G-CSF.Experimental featuresCombined laser microdissection, gene expression profiling to study motoneuron specific expression in an animal model of ALS, with and without neuroprotective treatment based on G-CSF.ConsentN/ASample source locationN/A
Direct link to deposited data
Deposited data can be found at: http://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE60856.
Experimental design, materials and methods
Amyotrophic lateral sclerosis (ALS) is a neurodegenerative disease characterized by the death of cortical, bulbar and spinal motoneurons. G-CSF is a hematopoietic growth factor with neuroprotective properties for motoneurons [2], [3]. Many studies have undertaken the study of the transcriptome of tissues or isolated cells from animal models or ALS patients [4]. The aim of the original study was to identify for the first time the transcriptional changes in motoneurons from SOD1 G93A mice upon G-CSF treatment [1].
Mice and treatment
Two cohorts of mice have been used within the study. A first cohort consisted of SOD1G93A mice and non-transgenic littermates of 11 weeks of age. These mice were used to detect early transcriptional response due to the SOD1 G93A transgene. The second cohort of mice consisted of SOD1 G93A and non-transgenic mice receiving the vehicle solution (250 mM sorbitol, 0.004% Tween-80 and 10 mM sodium acetate buffer (pH 4)), along with a group of SOD1 G93A mice receiving G-CSF (30 μg/kg/day). This cohort was included in order to detect transcripts deregulated by ALS and transcripts modulated by G-CSF.
The administration of vehicle or treatment was achieved by osmotic pumps, from week 11 to week 15. After the sacrifice, lumbar spinal cords were dissected and snap frozen.
Laser-microdissection and microarray
Special care was taken to avoid contamination with RNases. We have dissected 300 motor neurons per mouse using the Leica Laser Microdissection technology. Total RNA was extracted and amplified. Microarray was performed using the GeneChip® mouse genome 430 2.0 array from Affymetrix. Intensities were normalized using the Expression Console software from Affymetrix (http://www.affymetrix.com/estore/browse/level_seven_software_products_only.jsp?productId=131414).
Quality controls and analytical sequence
The efficacy of the intensity normalization was assessed with box plots showing probe cell intensity before normalization and relative probe cell intensity after normalization (Fig. 1A, B). After normalization, variability among arrays was strongly reduced, as monitored by the nearness between the median of log expression signals from each samples to the median array of all samples (red line, Fig. 1B).
Enrichment in mRNA from motoneurons was assessed by comparing the relative expression of cell specific genes [motoneurons: neurofilament-heavy chain (Nefh); astrocytes: glial fibrillary acidic protein (gfap); oligodendrocytes: SRY (sex determining region Y)-box 10(Sox10); microglia: allograft inflammatory factor 1-like (Aif1l); endothelial cells: claudin 5(Cldn5)] [5].
Based on the ratios between Nefh and the other cell specific transcripts, the laser microdissection led to an enrichment of motoneuronal mRNA estimated to a 135-fold as compared to astrocyte mRNA, to an 82-fold as compared to oligodendrocyte mRNA, to a 214-fold as compared to microglia mRNA and to a 384-fold as compared to endothelial mRNA (Fig. 2).
Principal component analysis (JMP 11.0.0, SAS Institute, Cary, North Carolina) was performed with the 1279 genes showing a standard deviation higher than 1.1 across all array to assess homogeneity among samples (Fig. 3). Age had a strong influence on the distribution of samples as shown by the segregation between samples from 11 weeks and 15 weeks old mice, regardless of the genotype.
At 15 weeks of age, samples from SOD1 G93A mice presented strong dispersion, presumably due to the variability of the pace of disease progression. It can be noted that five samples from the G-CSF treated mice regrouped together and were close to the non-transgenic or asymptomatic SOD1 G93A clusters. Samples from the placebo-treated SOD1 G93A mice presented the strongest dispersion. These data suggest that G-CSF had a substantial effect on the transcriptome of SOD1 G93A motoneurons.
Prior to supervised analysis, gene expression database was filtered for intensity signal (at least 4 arrays showed an intensity higher than centile 0.2) to remove background noise and for variability (standard deviation higher than centile 0.3) to discard transcripts with no or little changes in the expression across all arrays.
Discriminant analysis was used to determine how samples from SOD1 G93A mice treated with G-CSF relate to the other experimental groups. Only transcripts showing a standard deviation greater than 1.2 across all arrays were included in this analysis.
In an attempt to identify specific transcriptomic deregulation with biological relevance by ANOVA, we applied another filtering step, consisting in removing all transcripts with a fold-change expression lower than 1.2 when comparing non-transgenic and SOD1 G93A samples. The results generated by the discriminant analysis and by ANOVA are detailed in the original manuscript [1].
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Henriques A.Kastner S.Chatzikonstantinou E.Pitzer C.Plaas C.Kirsch F.Wafzig O.Kruger C.Spoelgen R.Gonzalez De Aguilar J.L.Gene expression changes in spinal motoneurons of the SOD 1(G 93A) transgenic model for ALS after treatment with G-CSF Front. Cell. Neurosci.820154642565359010.3389/fncel.2014.00464 PMC 4299451 · doi ↗ · pubmed ↗
- 2Henriques A.Pitzer C.Dittgen T.Klugmann M.Dupuis L.Schneider A.CNS-targeted viral delivery of G-CSF in an animal model for ALS: improved efficacy and preservation of the neuromuscular unit Mol. Ther.19220112842922113957210.1038/mt.2010.271PMC 3034860 · doi ↗ · pubmed ↗
- 3Henriques A.Pitzer C.Schneider A.Neurotrophic growth factors for the treatment of amyotrophic lateral sclerosis: where do we stand?Front. Neurosci.42010322059294810.3389/fnins.2010.00032 PMC 2902233 · doi ↗ · pubmed ↗
- 4Henriques A.Gonzalez De Aguilar JL: Can transcriptomics cut the Gordian knot of amyotrophic lateral sclerosis?Curr. Genomics 12720115065152254795710.2174/138920211797904043 PMC 3219845 · doi ↗ · pubmed ↗
- 5Cahoy J.D.Emery B.Kaushal A.Foo L.C.Zamanian J.L.Christopherson K.S.Xing Y.Lubischer J.L.Krieg P.A.Krupenko S.A.A transcriptome database for astrocytes, neurons, and oligodendrocytes: a new resource for understanding brain development and function J. Neurosci. Off. J. Soc. Neurosci.281200826427810.1523/JNEUROSCI.4178-07.2008 PMC 667114318171944 · doi ↗ · pubmed ↗