High-quality permanent draft genome sequence of the Parapiptadenia rigida-nodulating Cupriavidus sp. strain UYPR2.512
Sofie E De Meyer, Elena Fabiano, Rui Tian, Peter Van Berkum, Rekha Seshadri, TBK Reddy, Victor Markowitz, Natalia N Ivanova, Amrita Pati, Tanja Woyke, John Howieson, Nikos C Kyrpides, Wayne Reeve

TL;DR
This paper presents the genome sequence of a bacteria strain that forms nodules on a Uruguayan plant, providing insights into its genetic makeup and function.
Contribution
The novel contribution is the high-quality draft genome of Cupriavidus sp. UYPR2.512, isolated from Parapiptadenia rigida root nodules.
Findings
The genome is 7,858,949 bp long and arranged in 365 scaffolds.
It contains 7,411 protein-coding genes and 76 RNA-only encoding genes.
The genome is part of the GEBA-RNB project proposal.
Abstract
Cupriavidus sp. strain UYPR2.512 is an aerobic, motile, Gram-negative, non-spore-forming rod that was isolated from a root nodule of Parapiptadenia rigida grown in soils from a native forest of Uruguay. Here we describe the features of Cupriavidus sp. strain UYPR2.512, together with sequence and annotation. The 7,858,949 bp high-quality permanent draft genome is arranged in 365 scaffolds of 369 contigs, contains 7,411 protein-coding genes and 76 RNA-only encoding genes, and is part of the GEBA-RNB project proposal.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1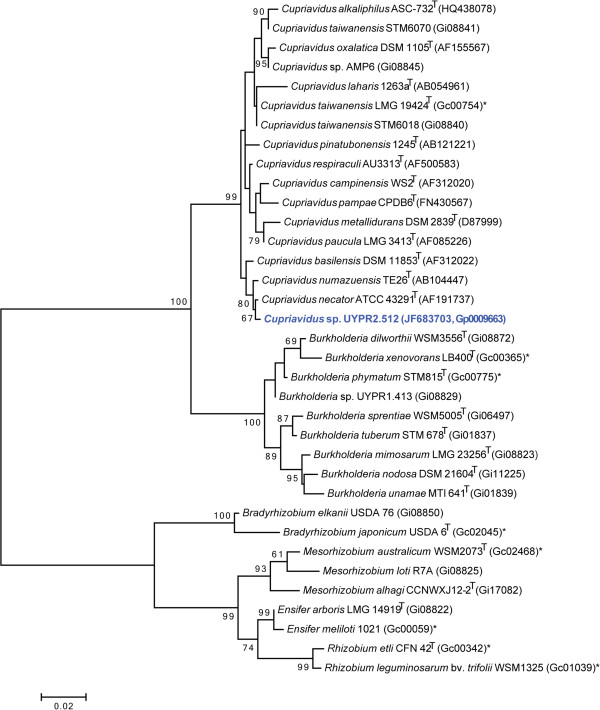 Figure 2
Figure 2 Figure 3
Figure 3| Current classification | Domain | TAS [ | |
| Phylum | TAS [ | ||
| Class | TAS [ | ||
| Order | TAS [ | ||
| Family | TAS [ | ||
| Genus | TAS [ | ||
| Species | IDA | ||
| Gram stain | Negative | IDA [ | |
| Cell shape | Rod | IDA | |
| Motility | Motile | IDA | |
| Sporulation | Non-sporulating | IDA [ | |
| Temperature range | mesophile | IDA [ | |
| Optimum temperature | 28°C | IDA | |
| pH range; Optimum | Not reported | ||
| Carbon source | Not reported | ||
| MIGS-6 | Habitat | Soil, root nodule on host | IDA |
| MIGS-6.3 | Salinity | Not reported | |
| MIGS-22 | Oxygen requirement | aerobic | IDA |
| MIGS-15 | Biotic relationship | Symbiotic | IDA |
| MIGS-14 | Pathogenicity | Non-pathogenic | NAS |
| MIGS-4 | Geographic location | Uruguay | IDA |
| MIGS-5 | Nodule collection date | 2006 | IDA |
| MIGS-4.1 | Latitude | -30.507 | IDA |
| MIGS-4.2 | Longitude | -57.71 | IDA |
| MIGS-4.4 | Altitude | 58 m | IDA |
| MIGS-31 | Finishing quality | Permanent-draft |
| MIGS-28 | Libraries used | Illumina Std PE |
| MIGS-29 | Sequencing platforms | Illumina HiSeq 2000 |
| MIGS-31.2 | Fold coverage | 106.8 X Illumina |
| MIGS-30 | Assemblers | Velvet 1.1.04, ALLPATHS-LG V.r41043 |
| MIGS-32 | Gene calling methods | Prodigal 1.4 |
| Locus Tag | A3A5 | |
| Genbank ID | ||
| Genbank Date of Release | September 16, 2013 | |
| GOLD ID | Gp0009663 [ | |
| BIOPROJECT | PRJNA165301 | |
| MIGS-13 | Source Material Identifier | UYPR2.512 |
| Project relevance | Symbiotic N2fixation, agriculture |
| Genome size (bp) | 7,858,949 | 100 |
| DNA coding (bp) | 6,709,332 | 85.37 |
| DNA G + C (bp) | 5,128,158 | 65.25 |
| DNA scaffolds | 365 | |
| Total genes | 7,487 | 100 |
| Protein coding genes | 7,411 | 98.98 |
| RNA genes | 76 | 1.02 |
| Pseudo genes | 0 | 0 |
| Genes in internal clusters | 419 | 5.6 |
| Genes with function prediction | 5,663 | 75.64 |
| Genes assigned to COGs | 4,807 | 64.20 |
| Genes with Pfam domains | 5,959 | 79.59 |
| Genes with signal peptides | 696 | 9.30 |
| Genes with transmembrane helices | 1,545 | 20.64 |
| CRISPR repeats | 1 |
| J | 183 | 3.37 | Translation, ribosomal structure and biogenesis |
| A | 1 | 0.02 | RNA processing and modification |
| K | 526 | 9.69 | Transcription |
| L | 192 | 3.54 | Replication, recombination and repair |
| B | 3 | 0.06 | Chromatin structure and dynamics |
| D | 35 | 0.65 | Cell cycle control, Cell division, chromosome partitioning |
| Y | 0 | 0.00 | Nuclear structure |
| V | 56 | 1.03 | Defense mechanisms |
| T | 210 | 3.87 | Signal transduction mechanisms |
| M | 277 | 5.11 | Cell wall/membrane/envelope biogenesis |
| N | 99 | 1.82 | Cell motility |
| Z | 0 | 0.00 | Cytoskeleton |
| W | 0 | 0.00 | Extracellular structures |
| U | 140 | 2.58 | Intracellular trafficking, secretion, and vesicular transport |
| O | 181 | 3.34 | Posttranslational modification, protein turnover, chaperones |
| C | 434 | 8.00 | Energy production and conversion |
| G | 268 | 4.94 | Carbohydrate transport and metabolism |
| E | 487 | 8.98 | Amino acid transport and metabolism |
| F | 89 | 1.64 | Nucleotide transport and metabolism |
| H | 194 | 3.58 | Coenzyme transport and metabolism |
| I | 337 | 6.21 | Lipid transport and metabolism |
| P | 272 | 5.01 | Inorganic ion transport and metabolism |
| Q | 236 | 4.35 | Secondary metabolite biosynthesis, transport and catabolism |
| R | 661 | 12.18 | General function prediction only |
| S | 545 | 10.04 | Function unknown |
| - | 2680 | 35.80 | Not in COGS |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGenomics and Phylogenetic Studies · Infections and bacterial resistance · Protist diversity and phylogeny
Introduction
Legumes establish symbiotic associations with a group of soil bacteria, rhizobia, able to fix atmospheric nitrogen (N_2_). Rhizobia elicit the formation of a symbiotic organ called a nodule comprising differentiated plant and bacterial cells. Differentiated rhizobia within nodules are termed bacteroids, and acquire the ability to fix nitrogen. Rhizobia are phylogenetically diverse including genera from the Alphaproteobacteria (Allorhizobium, Azorhizobium, Bradyrhizobium, Ensifer, Mesorhizobium, Rhizobium, etc.) as well as from the Betaproteobacteria (Burkholderia, Cupriavidus) [1,2].
The biological nitrogen fixation process significantly contributes to the development of sustainable agriculture reducing the use of supplies dependent on fuel and alleviating environmental impacts produced by the addition of chemical fertilizer [3]. Moreover, forestation with leguminous trees associated with rhizobia, “nitrogen-fixing trees”, has been successfully used for recovering degraded soils [4].
Parapiptadenia rigida (Benth.) Brenan, is a “nitrogen-fixing tree” belonging to the Piptadenia group from the Mimosoideae subfamily [5]. It is a multipurpose tree, very appreciated because of its timber and therefore used in high quality furniture and construction. It is also used for gums, tannins and essential oil extraction, has medicinal properties and is included in agroforestry and reforestation programs [4,6,7]. Taulé et al.[8] demonstrated that this species could be nodulated either by Alpha-rhizobia (Rhizobium) or by Beta-rhizobia (Burkholderia and Cupriavidus) with Burkholderia being the preferred natural symbiont of this legume. In the case of Cupriavidus sp. UYPR2.512, this strain was isolated from a nodule of a P. rigida plant grown in soils collected from Mandiyú native forest in Artigas, Uruguay. Isolated bacterial colonies of Cupriavidus sp. UYPR2.512 were able to nodulate and to promote the growth of P. rigida, as well as Mimosa pudica plants [8].
To our knowledge, the only published sequenced genome of a Beta-rhizobia belonging to the Cupriavidus genus so far is that of C. taiwanensis LMG 19424^T^[9]. Interestingly, the closest relative of Cupriavidus sp*.* UYPR2.512 is C. necator ATCC 43291^T^[8]. Here, we present the description of the Cupriavidus sp*.* UYPR2.512 high-quality permanent draft genome sequence and its annotation.
Organism information
Classification and features
Cupriavidus sp. strain UYPR2.512 is a motile, Gram-negative, non-spore-forming rod (Figure 1 Left, Center) in the order Burkholderiales of the class Betaproteobacteria. The rod-shaped form varies in size with dimensions of 0.5-0.7 μm in width and 0.9-1.2 μm in length (Figure 1 Left). It is fast growing, forming 0.5-0.8 mm diameter colonies after 24 h when grown on TY [10] at 28°C. Colonies on TY are white-opaque, slightly domed, moderately mucoid with smooth margins (Figure 1 Right).
Images of Cupriavidus sp. strain UYPR2.512 using scanning (Left) and transmission (Center) electron microscopy and the appearance of colony morphology on solid media (Right).
Figure 2 shows the phylogenetic relationship of Cupriavidus sp. strain UYPR2.512 in a 16S rRNA gene sequence based tree. This strain is the most similar to Cupriavidus necator ATCC 43291^T^, Cupriavidus oxalaticus DSM 1105^T^ and Cupriavidus taiwanensis LMG 19424^T^ based on the 16S rRNA gene alignment with sequence identities of 99.32%, 98.49% and 98.42%, respectively, as determined using the EzTaxon-e server [11]. Cupriavidus necator ATCC 43291^T^ has been isolated from soil and is a non-obligate predator causing lysis of various Gram-positive and Gram-negative bacteria in the soil [12]. Cupriavidus taiwanensis LMG 19424^T^ is a plant symbiont and was isolated from root nodules of Mimosa pudica collected from three fields at Ping-Tung Country in the southern part of Taiwan [1]. Minimum Information about the Genome Sequence (MIGS) is provided in Table 1 and Additional file 1: Table S1.
Phylogenetic tree highlighting the position of Cupriavidus sp. strain UYPR2.512 (shown in blue print) relative to other type and non-type strains in the Cupriavidus genus using a 1,034 bp internal region of the 16S rRNA gene. Several Alpha-rhizobia sequences were used as an outgroup. All sites were informative and there were no gap-containing sites. Phylogenetic analyses were performed using MEGA, version 5.05 [13]. The tree was built using the maximum likelihood method with the General Time Reversible model. Bootstrap analysis with 500 replicates was performed to assess the support of the clusters. Type strains are indicated with a superscript T. Strains with a genome sequencing project registered in GOLD [14] are shown in bold and have the GOLD ID mentioned after the strain number, otherwise the NCBI accession number has been provided. Finished genomes are designated with an asterisk.
Table 1: Classification and general features of Cupriavidus sp. strain UYPR2.512 in accordance with the MIGS recommendations[15]published by the Genome Standards Consortium[16]
Symbiotaxonomy
Cupriavidus sp. strain UYPR2.512 was isolated from Parapiptadenia rigida, a Mimosoideae legume native to Uruguay [8]. This tree is native to South America, including south Brazil, Argentina, Paraguay, and Uruguay, and used by locals for timber and as a source of gums, tannins and essential oils [8]. Cupriavidus sp. strain UYPR2.512 is able to renodulate its original host and is highly efficient in fixing nitrogen with this host [8]. A selection of other host plants, including Trifolium repens, Medicago sativa, Peltophorum dubium and Mimosa pudica were tested for their ability to nodulate with UYPR2.512. Of these plants, strain UYPR2.512 was only able to nodulate and fix nitrogen effectively with M. pudica[8].
Genome sequencing information
Genome project history
This organism was selected for sequencing on the basis of its environmental and agricultural relevance to issues in global carbon cycling, alternative energy production, and biogeochemical importance, and is part of the Genomic Encyclopedia of Bacteria and Archaea, The Root Nodulating Bacteria chapter (GEBA-RNB) project at the U.S. Department of Energy, Joint Genome Institute [25]. The genome project is deposited in the Genomes OnLine Database [14] and the high-quality permanent draft genome sequence in IMG [26]. Sequencing, finishing and annotation were performed by the JGI using state of the art sequencing technology [27]. A summary of the project information is shown in Table 2.
Table 2: Genome sequencing project information for Cupriavidus sp. strain UYPR2.512
Growth conditions and DNA isolation
Cupriavidus sp. strain UYPR2.512 was grown to mid logarithmic phase in TY rich media [10] on a gyratory shaker at 28°C. DNA was isolated from 60 mL of cells using a CTAB (Cetyl trimethyl ammonium bromide) bacterial genomic DNA isolation method [29].
Genome sequencing and assembly
The draft genome of Cupriavidus sp. UYPR2.512 was generated at the DOE Joint Genome Institute [27]. An Illumina Std shotgun library was constructed and sequenced using the Illumina HiSeq 2000 platform which generated 29,312,424 reads totaling 4,396.9 Mbp [30]. All general aspects of library construction and sequencing performed at the JGI can be found at the JGI web site [31]. All raw Illumina sequence data was passed through DUK, a filtering program developed at JGI, which removes known Illumina sequencing and library preparation artifacts (Mingkun L, Copeland A, Han J. unpublished). Artifact filtered sequence data was then screened and trimmed according to the k–mers present in the dataset. High–depth k–mers, presumably derived from MDA amplification bias, cause problems in the assembly, especially if the k–mer depth varies in orders of magnitude for different regions of the genome. Reads with high k–mer coverage (>30x average k–mer depth) were normalized to an average depth of 30x. Reads with an average kmer depth of less than 2x were removed. Following steps were then performed for assembly: (1) normalized Illumina reads were assembled using Velvet version 1.1.04 [32] (2) 1–3 Kbp simulated paired end reads were created from Velvet contigs using wgsim [33] (3) normalized Illumina reads were assembled with simulated read pairs using Allpaths–LG (version r41043)[34]. Parameters for assembly steps were: 1) Velvet (velveth: 63 –shortPaired and velvetg: -very clean yes –exportFiltered yes –min contig lgth 500 –scaffolding no –cov cutoff 10) 2) wgsim (-e 0 –1 100 –2 100 –r 0 –R 0 –X 0) 3) Allpaths–LG (PrepareAllpathsInputs: PHRED 64 = 1 PLOIDY = 1 FRAG COVERAGE = 125 JUMP COVERAGE = 25 LONG JUMP COV = 50, RunAllpathsLG: THREADS = 8 RUN = std_shredpairs TARGETS = standard VAPI_WARN_ONLY = True OVERWRITE = True). The final draft assembly contained 369 contigs in 365 scaffolds. The total size of the genome is 7.9 Mbp and the final assembly is based on 839.6 Mbp of Illumina data, which provides an average of 106.8x coverage.
Genome annotation
Genes were identified using Prodigal [35], as part of the DOE-JGI genome annotation pipeline [36,37] followed by a round of manual curation using GenePRIMP [38] for finished genomes and Draft genomes in fewer than 10 scaffolds. The predicted CDSs were translated and used to search the National Center for Biotechnology Information (NCBI) non-redundant database, UniProt, TIGRFam, Pfam, KEGG, COG, and InterPro databases. The tRNAScanSE tool [39] was used to find tRNA genes, whereas ribosomal RNA genes were found by searches against models of the ribosomal RNA genes built from SILVA [40]. Other non–coding RNAs such as the RNA components of the protein secretion complex and the RNase P were identified by searching the genome for the corresponding Rfam profiles using INFERNAL [41]. Additional gene prediction analysis and manual functional annotation was performed within the Integrated Microbial Genomes-Expert Review (IMG-ER) system [42] developed by the Joint Genome Institute, Walnut Creek, CA, USA.
Genome properties
The genome is 7,858,949 nucleotides with 65.25% GC content (Table 3) and comprised of 365 scaffolds and 369 contigs (Figure 3). From a total of 7,487 genes, 7,411 were protein encoding and 76 RNA only encoding genes. The majority of genes (75.64%) were assigned a putative function whilst the remaining genes were annotated as hypothetical. The distribution of genes into COG functional categories is presented in Table 4.
Table 3: Genome statistics for Cupriavidus sp. strain UYPR2.512
Graphical map of the four largest scaffolds of the genome of Cupriavidus sp. strain UYPR2.512. From the bottom to the top of each scaffold: Genes on forward strand (color by COG categories as denoted by the IMG platform), Genes on reverse strand (color by COG categories), RNA genes (tRNAs green, sRNAs red, other RNAs black), GC content, GC skew.
Table 4: Number of protein coding genes of Cupriavidus sp. strain UYPR2.512 associated with the general COG functional categories
Conclusion
Cupriavidus sp. UYPR2.512 belongs to a group of Beta-rhizobia isolated from Parapiptadenia rigida, a native tree from Uruguay belonging to the Mimosoideae legume group [8]. This tree is also native to the south of Brazil, Argentina and Paraguay [8]. Greenhouse experiments from previous studies have shown that Cupriavidus sp. UYPR2.512 is also able to nodulate and fix nitrogen with Mimosa pudica, an invasive species in many regions around the world [8]. Phylogenetic analysis revealed that UYPR2.512 is the most closely related to Cupriavidus necator ATCC 43291^T^, Cupriavidus oxalaticus DSM 1105^T^ and Cupriavidus taiwanensis LMG 19424^T^. In contrast to the other two strains, Cupriavidus taiwanensis LMG 19424^T^ is a microsymbiont that is able to nodulate and fix nitrogen in association with Mimosa species [43]. In total five Cupriavidus strains (AMP6, LMG 19424^T^, STM6018, STM6070 and UYPR2.512), which can form a symbiotic association have now been sequenced. A comparison of these strains reveals that UYPR2.512 has the largest genome (7.9 Mbp), with the highest KOG count (1398), the lowest G + C (65.25%) and signal peptide (9.3%) percentages in this group. All of these genomes share the nitrogenase-RXN MetaCyc pathway catalyzed by a multiprotein nitrogenase complex. Out of five Cupriavidus strains (AMP6, LMG 19424^T^, STM6018, STM6070 and UYPR2.512), which contain the N-fixation pathway, only Cupriavidus sp. UYPR2.512 has been shown to nodulate and fix effectively with Parapiptadenia rigida. The genome attributes of Cupriavidus sp. UYPR2.512 will therefore be important for ongoing molecular analysis of the plant microbe interactions required for the establishment of leguminous tree symbioses with this host.
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
EF supplied the strain and background information for this project, PVB supplied DNA to JGI, TR performed all imaging, SDM and WR drafted the paper, JH provided financial support and all other authors were involved in sequencing the genome and editing the final manuscript. All authors read and approved the final manuscript.
Supplementary Material
Additional file 1Associated MIGS Record.Click here for file
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Chen WM Laevens S Lee TM Coenye T De Vos P Mergeay M Ralstonia taiwanensis sp. nov., isolated from root nodules of Mimosa species and sputum of a cystic fibrosis patient Int J Syst Evol Microbiol 2001511729173510.1099/00207713-51-5-172911594603 · doi ↗ · pubmed ↗
- 2Moulin L Munive A Dreyfus B Boivin-Masson C Nodulation of legumes by members of the beta-subclass of Proteobacteria Nature 200141194895010.1038/3508207011418858 · doi ↗ · pubmed ↗
- 3Crews TE Peoples MB Legume versus fertilizer sources of nitrogen: ecological tradeoffs and human needs Agric Ecosyst Environ 200410227929710.1016/j.agee.2003.09.018 · doi ↗
- 4Farias JA Hoppe JM Vivian JAC Comportamento de mudas de Parapiptadenia rigida (Bentham) Brenan, submetidas a diferentes índices de luminosidade e em função de diferentes dimensões de recipientes Caderno de Pesquisa Série Biologia 2005176980
- 5Jobson RW Luckow M Phylogenetic study of the genus Piptadenia (Mimosoideae : Leguminosae) using plastid trn L-F and trn K/mat K sequence data Syst Bot 20073256957510.1600/036364407782250544 · doi ↗
- 6Schmidt CA Murillo R Bruhn T Bringmann G Goettert M Heinzmann B Catechin derivatives from Parapiptadenia rigida with in vitro wound-healing properties J Nat Prod 2010732035204110.1021/np 100523 s 21080642 · doi ↗ · pubmed ↗
- 7de Souza GC Haas A Pvon Poser GL Schapoval EE Elisabetsky E Ethnopharmacological studies of antimicrobial remedies in the south of Brazil J Ethnopharmacol 20049013514310.1016/j.jep.2003.09.03914698521 · doi ↗ · pubmed ↗
- 8Taule C Zabaleta M Mareque C Platero R Sanjurjo L Sicardi M New betaproteobacterial rhizobium strains able to efficiently nodulate Parapiptadenia rigida (Benth.) Brenan Appl Environ Microbiol 2012781692170010.1128/AEM.06215-1122226956 PMC 3298154 · doi ↗ · pubmed ↗