Theileria haneyi: An Emerging Equids Hemoparasite with Milder Virulence but Major Diagnostic and Therapeutic Implications
Bassma S. M. Elsawy, Heba F. Alzan

TL;DR
Theileria haneyi is a newly identified horse parasite that is hard to detect and resistant to treatment, requiring better diagnostic tools and research.
Contribution
The paper identifies T. haneyi as a distinct parasite with unique genetic and diagnostic challenges, highlighting the need for new detection and treatment methods.
Findings
T. haneyi cannot be detected by standard T. equi cELISA due to the absence of the ema-1 gene.
T. haneyi is resistant to imidocarb dipropionate and may interfere with T. equi clearance.
Species-specific PCR and ELISA assays have been developed but may be limited by genetic variation.
Abstract
Theileria haneyi, a recently discovered tick-borne hemoparasite infecting equids globally, has significant implications for equine health. Although it is closely related to T. equi (sharing 23% genomic divergence), it establishes an asymptomatic carrier state in persistently infected horses, creating a silent transmission reservoir. Its discovery and unique genetics justify its classification as a new taxon. A critical diagnostic challenge is that the lack of the ema-1 gene in T. haneyi prevents its detection by the standard T. equi cELISA, emphasizing the need for species-specific tools. Although species-specific PCR assays, including PCR and qPCR targeting genes like chr1sco or ema-11, respectively, and an indirect ELISA targeting the EMA-11 recombinant protein, have been developed, global genetic variations may limit their serological utility. Therapeutically, T. haneyi exhibits…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
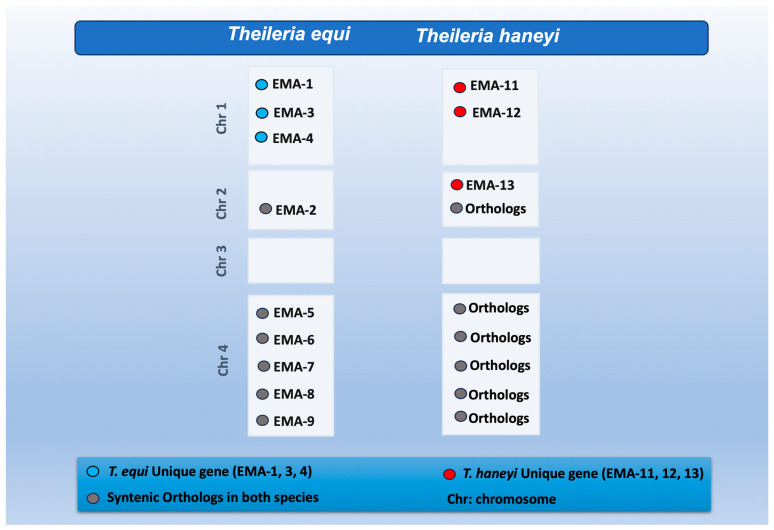 Figure 1
Figure 1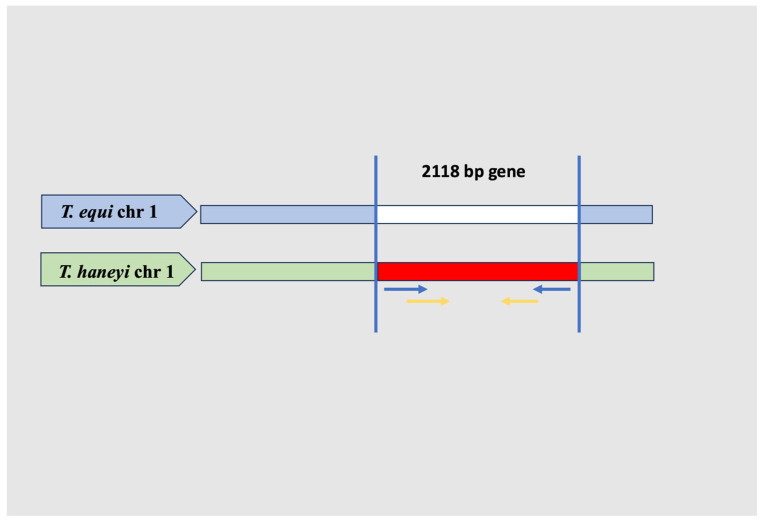 Figure 2
Figure 2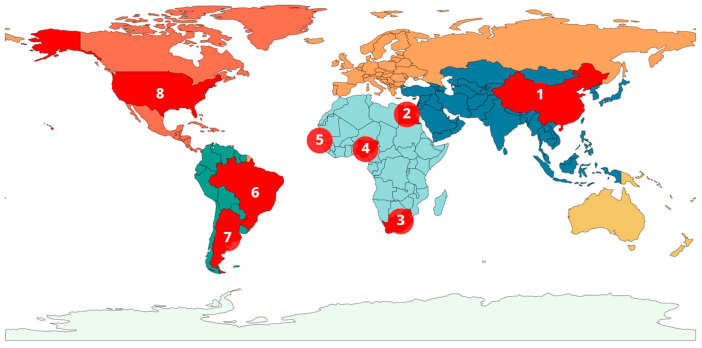 Figure 3
Figure 3- —National Research Centre, Cairo, Egypt
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsVector-borne infectious diseases · Parasitic Diseases Research and Treatment · Vector-Borne Animal Diseases
1. Introduction
Theileria (T.) haneyi is a recently discovered apicomplexan tick-borne hemoparasite that primarily infects equids, including horses [1] and donkeys [2]. It is closely related to T. equi, another causative agent of equine piroplasmosis (EP), a disease with significant global economic implications. It is morphologically smaller than T. equi in size. Theileria haneyi appears to have a global distribution, with infected equids having been identified in North and South America, Africa [1,3,4,5], China [6,7], and for the first time in donkeys and horses in Egypt by Elsawy et al. [2]. However, it remains unknown whether T. haneyi is capable of infecting vertebrate hosts other than equids as T. equi which can infect dogs, camels [8,9,10,11], cattle, sheep and goats [12,13]. In the meantime, there is a lack of knowledge regarding its geographical distribution and its prevalence worldwide.
The T. haneyi organism causes milder clinical disease (such as variable fever and anemia) than T. equi in experimentally infected horses and can be superinfected with T. equi [14]. Horses remain persistently infected following the acute stage of the disease, and these asymptomatic horses are presumed to be reservoirs of infectious organisms for competent tick vectors [14].
The global importance of T. haneyi to equine health was recently shown by its resistance to imidocarb dipropionate (ID) and its interference with T. equi clearance by ID in some co-infected horses. This review shows findings from various academic papers to provide a comprehensive overview of T. haneyi, discovery and genomic diversity, virulence and clinical progression, global geographical distribution, treatment efficacy, diagnostic methods progress, and transmission dynamics.
2. Data Collection
This study employed a systematic literature search to identify research on T. haneyi infections in equids worldwide. The primary search was conducted across PubMed, Scopus, and ScienceDirect, focusing on English-language publications from January 2018 to July 2026.
The search strategy utilized a combination of keywords covering the parasite, tick-borne diseases, virulence, transmission, treatment, diagnosis, and competent vectors. These terms were integrated with animal species (equids, horses, donkeys) and detection methods (microscopical, serological, and molecular examinations), along with geographical filters for global coverage. Boolean operators (“AND” “OR”) were used to construct precise search strings. Supplementary searches were performed on the Egyptian Knowledge Bank website for local journals and Google Scholar to access full-text publications.
3. Discovery and Genomic Diversity
Theileria haneyi was accidentally discovered in horses at the United States–Mexico border [1]. Phylogenetic analysis based on 18S rDNA revealed that this erythrocyte infective parasite is related to, yet distinct from, other Theileria species found in Africa, with its closest relatives being Theileria spp. from waterbuck and mountain zebra [1]. The observed sequence variability at the 18S rDNA locus suggests the potential existence of additional cryptic species within the genus [1].
Among the described species, the genome of this novel equine Theileria parasite is most similar to that of T. equi genotype C. However, the estimated divergence time between T. haneyi and T. equi, based on genomic sequence data, is over 33 million years [1,15]. The new species, T. haneyi, shows a high degree of genetic protein divergence from T. equi. This divergence (23%) is even greater than that between two other major, distinct species infecting domestic cattle and African/Asian wild buffalo (Theileria parva and Theileria annulata, respectively, “18%”) [1]. This significant genomic divergence, coupled with morphological differences, justifies the classification of T. haneyi as a new taxon [1].
Despite the overall genomic divergence, it was found that T. haneyi possesses a nine-member Equi Merozoite Antigen (EMA) superfamily, a multigene family previously thought to be exclusive to T. equi. Yet, significant sequence divergence in antigenic loci means that T. haneyi is not detectable by currently available diagnostic tests designed for T. equi [1]. Briefly, six of the nine T. equi EMA genes (ema-2, ema-5 to ema-9) have orthologs in T. haneyi, but T. equi’s ema-1, ema-3, and ema-4 (all on chromosome 1) are absent in the latter. This specific deletion of ema-1 in T. haneyi prevents its detection by the T. equi cELISA, which targets the EMA-1 protein. In contrast, T. haneyi has three unique EMA genes: ema-11 and ema-12 on chromosome 1, and ema-13 on chromosome 2 [1] (Figure 1). This highlights the challenges in apicomplexan parasite surveillance and equine piroplasmosis epidemiological studies, which consequently reflects the importance of developing specific diagnostic tools for the newly identified species T. haneyi.
Importantly, although the T. haneyi genome has been studied [1], the genome assemblies of T. haneyi still have gaps with uncertain chromosome structure, limiting biological interpretation. In a recent study, researchers generated a high-quality, gap-free genome assembly of the T. haneyi Eagle Pass isolate using a combination of long- and short-read sequencing technologies. Long-read sequencing enabled the assembly of near-complete chromosomes, while short-read data were used for error correction [16].
4. Transmission Dynamics
Understanding the transmission dynamics of T. haneyi is crucial for effective disease control. As a tick-borne hemoparasite, its transmission is primarily mediated by ticks. However, identifying competent tick vectors remains an area of ongoing research. A recent study investigated whether Haemaphysalis longicornis (H. longicornis), also known as the Asian longhorned tick, can transstadially transmit T. haneyi to horses or not [17]. This tick species is endemic to East Asia and has spread to other regions, including the USA, where it is known to transmit Theileria orientalis Ikeda genotype to cattle [17]. The study found that while H. longicornis larvae were efficient at acquiring T. haneyi from infected horses; there was no evidence of transstadial transmission to naïve horses [17]. This suggests that H. longicornis is not a competent vector for T. haneyi transmission, despite its ability to feed efficiently on horses and transmit related Theileria species [17]. This finding highlights a significant knowledge gap regarding the natural tick vectors for T. haneyi. Further research is needed to identify the specific tick species responsible for transmitting T. haneyi across different geographical regions. The ability of T. haneyi-infected horses to become asymptomatic carriers also contributes to the complexity of transmission, as these animals can serve as a silent reservoir for the parasite, facilitating its spread without overt clinical signs [14].
It is important to note that the future dissemination of T. haneyi is also driven by climate change, expanding tick habitats, and the global movement of asymptomatic equine carriers through trade. Iatrogenic transmission via contaminated veterinary equipment and complex co-infections with other parasites further complicate its dissemination and clinical impact.
5. Virulence and Clinical Progression
Initial studies demonstrated reduced clinical severity in spleen-intact horses. Furthermore, it was found that T. haneyi is less virulent than T. equi in splenectomized horses [14]. Theileria haneyi is a less virulent parasite than T. equi that can superinfect with T. equi in horses; it causes a delayed serological response and is not consistently detected by current T. equi diagnostic assays due to limited antigenic cross-reactivity [3]. In early experiments, splenectomized horses survived T. haneyi infection and progressed to an asymptomatic carrier state, in stark contrast to the high fatality rate of T. equi in splenectomized horses. Thus, it was theorized that T. haneyi is less virulent than T. equi. This was confirmed by Sears et al. [14], who evaluated clinical data from splenectomized, T. haneyi-infected horses and found that seven of the eight splenectomized, T. haneyi-infected horses survived and progressed to an asymptomatic carrier state. These data contrast with the high fatality rate observed in T. equi-infected splenectomized horses without intermediation [14]. This reduced virulence of T. haneyi is probably attributed to its genomic reduction, as its genome is approximately 2 Mbp smaller than that of T. equi [14].
In cases of acute T. haneyi infection in splenectomized horses, the symptoms included fever (102.4 to 105.8 °F), anemia through decreased packed cell volume (PCV), and variable levels of parasitemia. Parasitemia appeared in waves, with some horses maintaining low levels while others experienced intermittent increases. The ability of T. haneyi-infected horses to become asymptomatic carriers has significant implications for disease control and surveillance. These animals can serve as a continuous reservoir for transmission [14], as with T. equi, where clinical signs are often subtle without careful observation. However, clinical signs can vary significantly in uncontrolled environments, especially in working or athletic animals [14].
6. Diagnostic Methods Progress
The accurate diagnosis of T. haneyi infection is critical for effective management and control, especially given its potential to co-infect with T. equi and its resistance to common treatments. Traditional diagnostic methods for equine piroplasmosis, such as nPCR assays targeting T. equi genes, have been found to be unreliable for detecting T. haneyi due to significant sequence divergence in antigenic loci [1]. To overcome this issue, a gene coding for a protein of unknown function in the syntenic T. haneyi locus of the vacated ema1 gene was selected to detect T. haneyi by PCR assay [1,3]. Certain studies used species-specific nested PCR-targeting this gene, which is specific only for T. haneyi, to detect its prevalence in countries such as Nigeria (2.7% in horses) [5], South Africa [18] and Egypt (53.1% in horses and 38.1% in donkeys) [2]. In China, Yang et al. [7] used nPCR targeting chromosome 1 single-copy (chr1sco) open reading frame genes, which have no detectable orthologs in T. equi or B. caballi (Figure 2), for the detection of T. haneyi (prevalence 11.7%).
Related studies have developed a new real-time quantitative PCR (qPCR) method based on the chr1sco gene, which is considered a straightforward, rapid and sensitive diagnostic method to detect T. haneyi [6]. However, false-negative results and a poor correlation between the results of the nested PCR and qPCR assays were reported [6], suggesting that this qPCR assay may not be as specific or as sensitive as the nested PCR in detecting T. haneyi infections when based on the chr1sco gene. Bhoora et al. [19] targeted the equi merozoite antigen 11 (Th. ema-11) gene, which is not found in T. equi, as a target for qPCR assays. It has demonstrated efficiency, specificity, and sensitivity in detecting T. haneyi ema-11. A recent study utilized the EMA 11 gene and EMA 10 genes for the detection of T. haneyi in Brazil using conventional PCR and semi-nested PCR assays, with a prevalence of 2.1% in an imported horse from Texas, US, and kept in an equestrian center in Brazil [20]. However, the positive sample in the PCR assays targeting the ema-10 and ema-11 genes tested negative in the 18S rRNA and ema-12-based PCR, which limited a more precise characterization of the T. haneyi detected in that study [20]. Nevertheless, the first detection of T. haneyi in Argentina was reported based on 18S rDNA-based PCR [21].
Recent advancements have focused on developing more specific serological diagnostic tools to detect the presence of antibodies against T. haneyi in animal serum. An indirect (i)ELISA based on the equi merozoite antigen 11 (ThEMA11) recombinant EMA-11 protein of T. haneyi was developed and shown promise in detecting geographically diverse T. haneyi strains in the sera of infected horses from different countries around the world [22]. Since the collected samples were from the USA, Germany, Mexico, France, Ireland, Puerto Rico and the Netherlands, there is a risk of potential variation in the EMA-11 protein sequence, which might limit the effectiveness of the indirect ELISA outside of these regions [19]. The same iELISA was utilized in China and showed 16.41% seroprevalence in a cross-sectional analysis of 2627 equine samples [7].
7. Global T. haneyi Geographical Distribution Analysis
Theileria haneyi was first detected in 2018 in a horse at the United States–Mexico border, North America [1]. This initial finding was a result of surveillance efforts, specifically near Eagle Pass, Texas, which led to the parasite sometimes being referred to as the T. haneyi (EP) strain [1]. Detection began to be performed in other countries on the Africa continent, such as South Africa [18], Nigeria [5], Gambia [23] and Egypt [2]. Later, it was detected in Asian countries such as China [7], in addition to Brazil and Argentina in South America [20,21]. Current information indicates that T. haneyi is distributed across three continents: North America, Africa, and Asia. In Europe, Facile et al. [24] examined equine samples for the presence of T. haneyi, targeting the chr1sco gene in nPCR, but all samples were negative. However, there is still a lack of data regarding its distribution in other continents. Even though the presence of T. haneyi has been identified on some content, the individual countries have not yet been surveyed within the same continent. This emphasized the need for further surveillance studies in other countries to cover their global range (Table 1 and Figure 3).
8. Treatment Efficacy
There is a significant therapeutic gap in the treatment of equine piroplasmosis caused by T. haneyi, as evidenced by studies on the effectiveness of common antiparasitic drugs [17] (Table 2). It has been shown that T. haneyi is resistant to several widely used therapeutic agents, including imidocarb (ID) dipropionate, buparvaquone, tulathromycin, and diclazuril [25]. Imidocarb dipropionate is often the drug of choice for treating equine piroplasmosis and controlling clinical signs of T. equi infection worldwide [25]. However, it fails to treat T. haneyi-infected horses. Compounding the issue, T. haneyi co-infection can hinder the successful treatment of T. equi by ID dipropionate [14]. Thus, this resistance is a significant concern. Additionally, growing concerns regarding ID-resistant parasite strains and their associated toxicity have highlighted the urgent need for novel, safer, and more effective antiparasitic agents.
The in vitro efficacy of tafenoquine succinate (TFQ), a synthetic 8-aminoquinoline with broad antiparasitic activity, against T. equi and B. caballi as a potential treatment for equine piroplasmosis was tested by Cardillo et al. [26]. However, it has not yet been used for the treatment of T. haneyi. It was found that TFQ has potent in vitro activity against T. equi and moderate activity against B. caballi, along with a mild cytotoxic profile in equine Peripheral Blood Mononuclear Cells (PBMCs). Moreover, certain studies have investigated the temporary efficacy of buparvaquone against T. haneyi in chronically infected horses. It was found that the recommended dose of 2.5 mg/kg led to a rapid reduction in T. haneyi levels to undetectable levels by nested PCR (nPCR), but the recurrence occurred after a minimum of seven weeks [25]. Subsequent re-administration of buparvaquone at an increased dosage of 6 mg/kg failed to produce a theilericidal effect upon recrudescence [25]. This suggests that buparvaquone may offer only temporary suppression rather than a definitive cure for T. haneyi. Accordingly, this complicates treatment strategies for horses infected by Theileria spp., highlighting the need for new therapeutic approaches that are effective against T. haneyi and do not compromise the treatment of co-existing T. equi infections.
9. Conclusions and Recommendations
Theileria haneyi is a less virulent parasite compared to T. equi, yet it poses significant challenges to global equine health due to its drug resistance and ability to establish asymptomatic carrier states. While recent genomic advancements research provides an establishment for understanding its biology, the parasite’s distinct genetic profile from T. equi demands the global validation of specific diagnostic tools such as qPCR and nPCR targeting T. haneyi’s unique genes, such as the EMA 11 and the chr1sco open reading frame genes.
Future research must spotlight identifying its competent tick vectors through detecting the presence of this parasite in different ticks in the field that infest the farm animals to draw a wider picture for parasite spreading vectors and exploring the associated co-infections across diverse livestock like cattle, camel, sheep and goats to fully interpret its transmission dynamics. Finally, developing targeted therapeutics and vaccines is essential to relieve its impact on equine health and prevent its further spread. The insights gained from studying T. haneyi not only contribute to our understanding of equine piroplasmosis but also provide a foundation for defining virulence mechanisms within the broader Apicomplexa phylum, as well as the synergetic effect regarding drug resistance.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Knowles D.P. Kappmeyer L.S. Haney D. Herndon D.R. Fry L.M. Munro J.B. Sears K. Ueti M.W. Wise L.N. Silva M. Discovery of a Novel Species, Theileria haneyi n. Sp., Infective to Equids, Highlights Exceptional Genomic Diversity within the Genus Theileria: Implications for Apicomplexan Parasite Surveillance Int. J. Parasitol.20184867969010.1016/j.ijpara.2018.03.01029885436 · doi ↗ · pubmed ↗
- 2Elsawy B.S.M. Nassar A.M. Alzan H.F. Bhoora R.V. Ozubek S. Mahmoud M.S. Kandil O.M. Mahdy O.A. Rapid Detection of Equine Piroplasms Using Multiplex PCR and First Genetic Characterization of Theileria haneyi in Egypt Pathogens 202110141410.3390/pathogens 1011141434832570 PMC 8620363 · doi ↗ · pubmed ↗
- 3Sears K.P. Kappmeyer L.S. Wise L.N. Silva M. Ueti M.W. White S. Reif K.E. Knowles D.P. Veterinary Parasitology Infection Dynamics of Theileria equi and Theileria haneyi, a Newly Discovered Apicomplexan of the Horse Vet. Parasitol.2019271687510.1016/j.vetpar.2019.06.00931303207 · doi ↗ · pubmed ↗
- 4Bishop R.P. Kappmeyer L.S. Onzere C.K. Odongo D.O. Githaka N. Sears K.P. Knowles D.P. Fry L.M. Equid Infective Theileria Cluster in Distinct 18S RRNA Gene Clades Comprising Multiple Taxa with Unusually Broad Mammalian Host Ranges Parasites Vectors 20201326110.1186/s 13071-020-04131-032430015 PMC 7236219 · doi ↗ · pubmed ↗
- 5Mshelia P.W. Kappmeyer L. Johnson W.C. Kudi C.A. Oluyinka O.O. Balogun E.O. Richard E.E. Onoja E. Sears K.P. Ueti M.W. Molecular Detection of Theileria Species and Babesia Caballi from Horses in Nigeria Parasitol. Res.20201192955296310.1007/s 00436-020-06797-y 32647992 PMC 7431391 · doi ↗ · pubmed ↗
- 6Zhou B. Yang G. Hu Z. Chen K. Guo W. Wang X. Du C. Development of a Real-Time Quantitative PCR Based on a Taqman-MGB Probe for the Rapid Detection of Theileria haneyi Microorganisms 202311263310.3390/microorganisms 1111263338004645 PMC 10673206 · doi ↗ · pubmed ↗
- 7Yang G. Chen Y. Chen K. Hu Z. Li J. Wang J. Guo W. Wang X. Du C. Prevalence and Molecular Epidemiology of the Novel Equine Parasite Theileria haneyi in China Equine Vet. J.20255863864910.1111/evj.1450540257377 · doi ↗ · pubmed ↗
- 8Matjila P.T. Leisewitz A.L. Oosthuizen M.C. Jongejan F. Penzhorn B.L. Detection of a Theileria Species in Dogs in South Africa Vet. Parasitol.2008157344010.1016/j.vetpar.2008.06.02518687528 · doi ↗ · pubmed ↗