Lifestyle Intervention Therapy Modulates Global DNA Methylation and Adipogenic Gene Expression in Severely Obese Hypogonadal Men
Siresha Bathina, Virginia Fuenmayor Lopez, Mia Prado, Salina Biene Teo, Dennis T. Villareal, Rui Chen, Clifford Qualls, Reina Armamento-Villareal

TL;DR
Lifestyle changes like diet and exercise can alter DNA methylation and gene expression in severely obese men with low testosterone.
Contribution
This study shows lifestyle interventions can reduce DNA methylation and specific gene expression in hypogonadal obese men.
Findings
LSI therapy significantly reduced global DNA methylation and 5-methylcytosine levels.
LSI decreased mRNA levels of DNMT1, DNMT3A, and DNMT3B.
LSI markedly decreased CEBPα, FTO, and PPARγ mRNA expression.
Abstract
Background/Objectives: Previous studies have suggested that lifestyle intervention (LSI) therapies involving diet and exercise can modulate DNA methylation; however, whether this occurs in severely obese hypogonadal men undergoing weight loss from diet and exercise remains unclear. Methods: In this study, we investigated the effects of weight loss from diet and exercise on global DNA methylation as well as on the mRNA expression of specific demethylation enzymes, DNMT1, DNMT3A, and DNMT3B—in peripheral blood mononuclear cells (PBMCs) and DNA methylation markers in DNA of severely obese hypogonadal men. This is a secondary analysis of samples of severely obese (body mass index of ≥35 kg/m2) hypogonadal men undergoing weight loss from diet and exercise in addition to an aromatase inhibitor (anastrozole) or placebo for a total of 12 months. Results: LSI therapy significantly reduced global…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
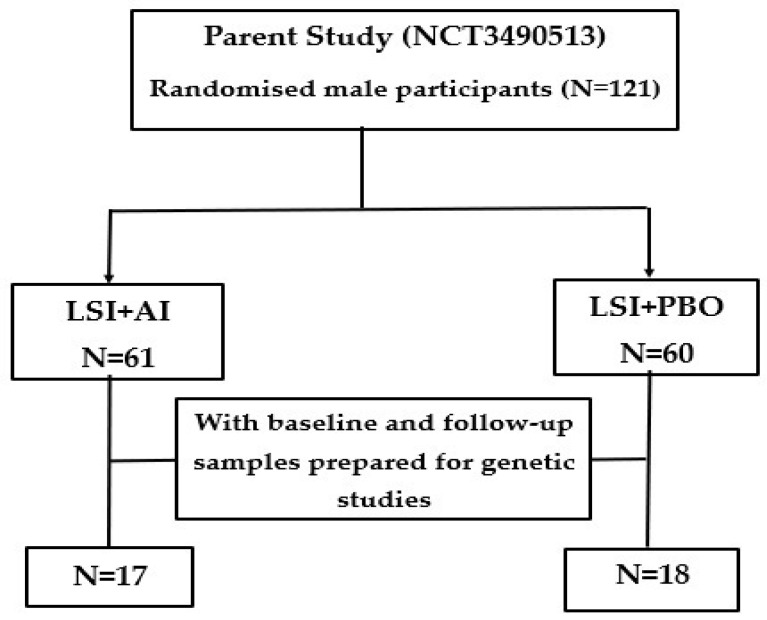 Figure 1
Figure 1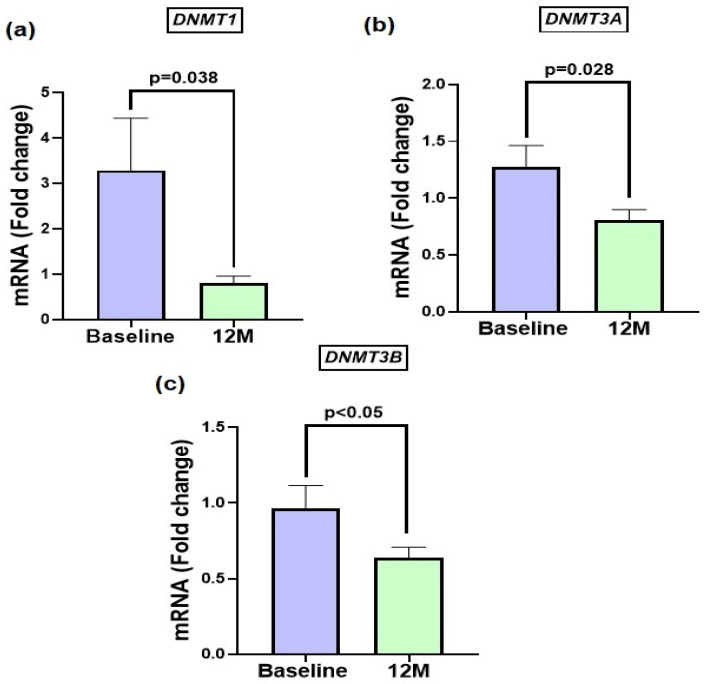 Figure 2
Figure 2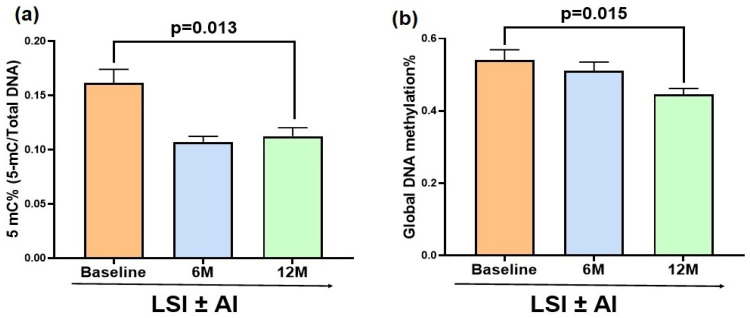 Figure 3
Figure 3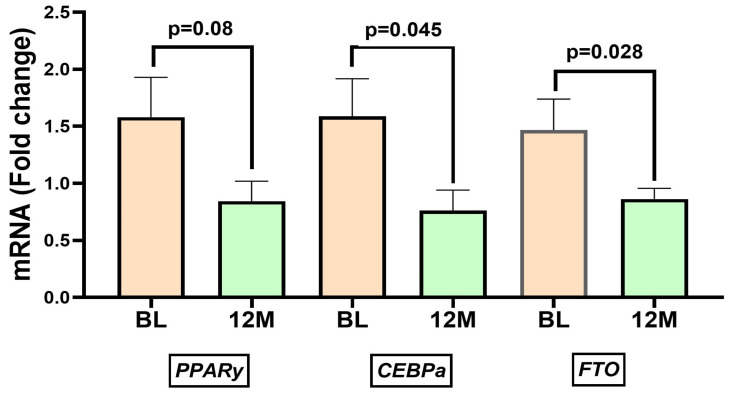 Figure 4
Figure 4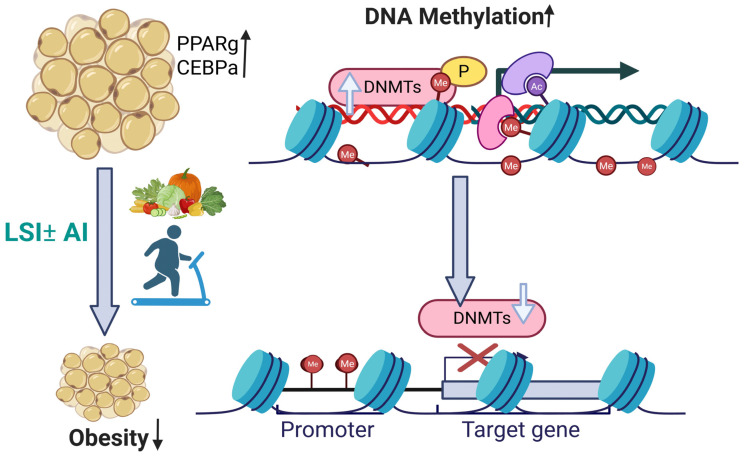 Figure 5
Figure 5- —NIH
- —VA Merit Review
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsEpigenetics and DNA Methylation · Dietary Effects on Health · Nutrition, Genetics, and Disease
1. Introduction
Obesity has become a global epidemic, contributing to a wide spectrum of metabolic and endocrine disorders including type 2 diabetes mellitus, cardiovascular disease, and male hypogonadism [1,2,3]. The recent literature suggests that obesity and related metabolic dysfunctions are accompanied by alterations in epigenetic regulation, particularly DNA methylation [4], which may mediate the interaction between environmental and genetic factors [5,6].
DNA methylation, the covalent addition of a methyl group to cytosine residues within CpG dinucleotides, is one of the most studied epigenetic modifications regulating gene expression [7]. This process is catalyzed by DNA methyltransferases (DNMTs), including DNMT1, DNMT3A, and DNMT3B, which play distinct roles in maintaining and establishing methylation patterns [8]. Aberrant DNA methylation has been implicated in several obesity-related pathologies, influencing genes involved in lipid metabolism, adipogenesis, and insulin sensitivity [9]. Specifically, the peroxisome proliferator-activated receptor gamma (PPARγ) and CCAAT/enhancer-binding protein alpha (CEBPα) are critical transcription factors governing adipocyte differentiation and glucose homeostasis, and their epigenetic regulation has been associated with obesity and metabolic dysfunction [10,11]. Our group showed that combined diet and exercise resulted in greater improvement in physical function [12] and improvement in metabolic profile [13] than diet or exercise alone. Beyond improving body composition and metabolic health, Keller et al.’s studies on 120 adults (BMI: ~30 kg/m^2^) in an 18-month diet +/− physical activity lifestyle randomized trial found that successful weight loss was associated with specific genome-wide DNA methylation changes in blood [14]. Aerobic exercise has been reported to modify DNA methylation in skeletal muscle, adipose tissue, and leukocytes, leading to altered expression of metabolic and inflammatory genes [15]. Dietary factors such as caloric restriction and nutrient composition can also influence methylation status by modulating one-carbon metabolism and methyl donor availability [16]. Despite these insights, the combined effects of diet and exercise on DNA methylation in severely obese hypogonadal men remain largely unexplored.
A prior study in obese individuals found that increased global DNA methylation and elevated DNMT1 expression have been associated with systemic inflammation and insulin resistance [17]. Conversely, in a rat study, lifestyle modification has been linked to reduced Dnmt activity and demethylation of genes regulating metabolism and inflammation [18]. However, most of these findings are derived from pre-clinical and general obese patients, but specific epigenetic mechanisms through which lifestyle interventions (LSIs) influence metabolic outcomes in severely hypogonadal men have not been clearly elucidated. Given these gaps in knowledge, the present study aimed to investigate the effects of LSIs by diet and exercise to promote weight loss on global DNA methylation in peripheral blood mononuclear cells (PBMCs) of obese hypogonadal men. We focused on the methylation and expression of PPARγ and CEBPα as representative metabolic regulators, and the expression of DNMT1, DNMT3A, and DNMT3B as key enzymes of DNA methylation machinery. By integrating molecular and physiological data, this study seeks to provide mechanistic insights into how LSIs modulate epigenetic pathways which may ultimately contribute to improved metabolic and endocrine function in obese hypogonadal men.
2. Materials and Methods
2.1. Study Design and Patient Population
This was a secondary analysis of DNA and RNA samples obtained from participants of the study, titled “Aromatase inhibitors and weight loss in severely obese men with hypogonadism” (NCT03490513). This study was a randomized double-blind placebo-controlled trial on the effect of weight loss from LSI in combination with an aromatase inhibitor (LSI + AI), anastrozole 1 mg daily, versus placebo (LSI + PBO) over 1 year on hormonal profile and symptoms of hypogonadism in men with severe obesity and hypogonadism. Inclusion/exclusion criteria were as previously published [19], but briefly, the study recruited severely obese men (BMI of ≥35 kg/m^2^), 35–65 years old, with an average fasting total T done twice between 8 a.m. and 10 a.m. on 2 separate days within 1 month of <300 ng/dL, with luteinizing hormone (LH) of <9.0 mIU/L and estradiol (E2) of ≥14 pg/mL, and with symptoms consistent with hypogonadism. The exclusion criteria included the following: (1) clinical/biochemical evidence of hypothalamic/pituitary disease; (2) drugs affecting gonadal hormone levels, production and action, or bone metabolism (bisphosphonates, teriparatide, denosumab, glucocorticoids, and phenytoin); (3) diseases affecting bone metabolism (e.g., hyperparathyroidism, untreated hyperthyroidism, osteomalacia, chronic liver disease, significant renal failure, hypercortisolism, malabsorption, immobilization, and Paget’s dis.); (4) prostate carcinoma or elevated serum PSA > 4 ng/mL; (5) hematocrit (HCT) more than 50%; (6) untreated severe obstructive sleep apnea; (7) cardiopulmonary disease (e.g., myocardial infarction within 6 months, unstable angina, and stroke) or unstable disease (e.g., NYHA Class III or IV congestive heart failure, severe pulmonary disease requiring steroid pills or the use of supplemental oxygen that would contraindicate exercise or dietary restriction); (8) unstable weight (i.e., ±2 kg) in the last 3 months; (9) BMD T-score of less than −2.0 at the spine, femoral neck, or total femur; (10) T2DM with fasting blood glucose of >160 mg/dL or A1C of >9.5%.
This study was conducted at Michael E. DeBakey VA Medical Center and the protocol was approved by the Institutional Review Board of Baylor College of Medicine. All participants provided written informed consent in accordance with the guidelines in the Declaration of Helsinki for the ethical treatment of human subjects. This study started in May 2018 and ended in June 2025.
This specific study focused on the methylation changes over time on a subset of subjects with DNA and RNA taken at least at 2 timepoints during the intervention (please see Figure 1 below).
2.2. Diet and Exercise Intervention
Dietary intervention was managed with the aid of a dietitian. Participants were instructed to consume a balanced diet to provide a deficit of 500–750 kcal/day from daily energy requirement. Follow-up visits with dietitian were weekly for the first 3 months and then every 2 weeks thereafter.
Exercise training was supervised by an exercise physiologist and consisted of aerobic training for ~45 min in duration and involved walking on a treadmill and stationary cycling. The resistance training involved nine upper-extremity and lower-extremity exercises with the use of weight-lifting machines. Initially, participants exercised in-person at our facility twice a week. The participants were also instructed to perform home-based exercises involving ground walking, treadmill walking, or stationary biking if available as well as resistance exercise using body weight (e.g., abdominal crunches), resistance bands, and ankle weights. However, because of COVID-19 pandemic restrictions, patients were later allowed to perform exercises in a gym of their choice paid for the study or just do home exercises using as much as possible the same routine.
2.3. Body Mass Index (BMI)
Body weight and height were measured by a standard weighing scale and stadiometer, respectively. BMI (kg/m^2^) was calculated by dividing the weight (in kilograms) by height (in meters) squared.
2.4. Gene Expression Studies
Blood samples were collected early in the morning after an overnight fast and processed, and then, the samples were stored at −80 °C until analysis. PBMCs were isolated from whole blood using Ficoll density gradient centrifugation. The PBMC fraction contained lymphocytes (T cells, B cells, and NK cells) and monocytes but excluded granulocytes such as neutrophils. Gene expression of PPARγ, CEBPa, FTO, DNMT1, DNMT3A, and DNMT3B in PBMCs was performed by real-time quantitative polymerase chain reaction at baseline (BL) and at 12 months (12 M).
2.4.1. RNA Extraction and qPCR Studies
RNA was extracted from PBMCs using RiboPure Blood (Invitrogen, Carlsbad, CA, USA #AM1928). A total of 200 ng of RNA was used for retro transcription into cDNA and performed using Superscript VILO Master Mix (Invitrogen, Carlsbad, CA, USA) in triplicates following protocol instructions. FAM-labeled TaqMan gene expression assays (Applied Biosystem, College Station, TX, USA) were used for PPARγ (Assay ID:), CEBPα (Hs00269972_s1:), FTO (Hs01057145_m1), DNMT1 (Hs00945875_m1), DNMT3A (Hs01027162_m1), and DNMT3B (Hs00171876_m1), and a VIC-labeled TaqMan gene expression assay for housekeeping 18S (assay ID: Hs03928990_g1). TaqMan Universal Master Mix was used following the manufacturer’s protocol. Please see the supplementary details of kits and chemicals used in this study.
Relative quantification: Relative quantification of gene expression of our samples was compared with that of human control total RNA, analyzed by TaqMan-based real-time PCR analysis (Applied Biosystems, #4307281, Carlsbad, CA, USA) was calculated using the DDCT method and adjusted for housekeeping gene expression. Data analysis was performed using a real-time PCR system QuantStudio5 and Quant Studio Design & Analysis Software 1.3.1.
2.4.2. DNA Extraction and Methylation Studies
Genomic DNA was isolated from peripheral leukocytes [20] according to the manufacturer’s instructions. DNA quality and quantity were assessed by spectrophotometry (NanoDrop) Bioanalyzer 2100 (Agilent Technologies, Santa Clara, CA, USA). and samples were normalized to 200 ng/µL for methylation analysis. Global DNA methylation (5-methylcytosine, 5-mC) was quantified using the Global DNA Methylation Assay Kit (5-Methyl Cytosine, Colorimetric; Abcam, Cambridge, UK; Cat. No. ab233486) following the manufacturer’s instructions.
Global DNA methylation was expressed as the percentage of 5-methylcytosine (5-mC%) relative to total cytosine content and calculated according to the manufacturer’s instructions. Values are expressed as the fraction of total methylated cytosine. Because the primary objective of this secondary analysis was to evaluate sustained epigenetic changes following long-term lifestyle intervention, we analyzed specimens collected at baseline (BL), 6 months, and 12 months (12 M), representing the full duration of the intervention for global methylation studies.
2.4.3. Statistical Analysis
Overall longitudinal analysis of 5 mC% or global DNA methylation (GM) were performed by repeated measures ANOVA with visit (baseline, 6 months, and 12 months) as the repeated factor adjusted for the baseline outcome value and treatment group (lifestyle with AI or placebo); change in testosterone, estradiol, or weight; or metformin use as covariates. Comparisons of 6- and 12-month values with baseline values for the different methylation parameters were performed by Student’s t-test. Data were presented as the means ± SD in tables and text, and the means ± SE in the figures. Data in graphs were analyzed using Prism 9.0 (GraphPad, San Diego, CA, USA) and tables were managed using Excel 2013 (Microsoft, Redmond, WA, USA), analyzed by Statgraphics Centurion XVI X64 (Statgraphics Technologies, Inc., The Plains, VA, USA) and confirmed using SAS version 9.3 (SAS Institute, Inc., Cary, NC, USA) A p-value of < 0.05 was considered statistically significant.
3. Results
Thirty-five participants with RNA and DNA samples taken at least at 2 timepoints were included in this study (Figure 1). The average baseline weight of these subjects was 135.9 ± 20.5 kg, and the baseline BMI was 43.8 ± 5.3 kg/m^2^. Table 1 below shows the baseline characteristics of the participants included in the study. There were no significant differences in clinical characteristics between those randomized to LSI + AI compared to the LSI + PBO at baseline. In addition, there were no significant differences in baseline characteristics of the entire population and the subjects included in this study (see Supplementary Table S2).
3.1. Body Composition
At the end of the study, the subjects lost an average of −3.7 ±3.6% (−6.7 ± 7.1 kg). Testosterone significantly increased in the LSI + AI group compared to the LSI + PBO group (+174.6+/−134.3 ng/dL vs. +43.8+/−103.5 ng/dL; respectively, p = 0.01), and estradiol significantly decreased in the LSI + AI group compared to LSI + PBO group (−13.9+/−19.0 pg/mL vs. +1.5+/−7.7 pg/mL; respectively, p = 0.01) at the end of 12 months. Table 2 below shows the changes in body composition of the participants compared to baseline at 6 and 12 months. Body fat, visceral adipose tissue, and lean mass (total and appendicular) decreased from baseline. However, these changes were not statistically significant.
3.2. DNMTs Expression Is Downregulated Following Diet and Exercise Intervention
Lifestyle with or without aromatase inhibitors (LSI ± AI) significantly modulated the expression of DNA methylation-related genes (Figure 2). Reduction in mRNA expression was observed at 12 months compared to baseline for DNMT1 (BL: 3.29 ± 1.1 vs. 12 M: 0.78 ± 0.16, p = 0.038), DNMT3A (BL: 1.28 ± 0.18 vs. 12 M: 0.81 ± 0.09, p = 0.028), and DNMT3B (BL: 0.97 ± 0.15 vs. 12 M: 0.64 ± 0.07, p = 0.05) following the intervention (Figure 2a–c). Collectively, these findings demonstrate a consistent downregulation of key components of DNA methylation machinery after the 12-month lifestyle program.
3.3. Lifestyle Modification Reduces Global and 5-mC% DNA Methylation
To validate whether the transcriptional changes were accompanied by epigenetic alterations at the DNA level, global DNA methylation markers were assessed in the PBMC samples collected at baseline (BL), 6 months (6 M), and 12 months (12 M). LSI ± AI significantly reduced the percentage of 5-mC%, with levels decreasing from BL (0.18 ± 0.03) to 6 M (0.103 ± 0.005; p = 0.052) and 12 M (0.081 ± 0.004; p = 0.013) (Figure 3a). Similarly, global DNA methylation showed a downward trajectory, declining from BL (0.85 ± 0.18) to 6 M (0.49 ± 0.03; p = 0.053) and 12 M (0.39 ± 0.014; p = 0.015) (Figure 3b). Collectively, these data indicate that LSI ± AI promotes gradual genomic DNA demethylation in PBMCs over the 12-month intervention period.
To examine the influence of AI, as half of the subjects on this study were on it and hormone studies showed significant differences in testosterone and estradiol levels between those who were and were not on AI, we compared the change in 5-mC% and global methylation at 6 and 12 months between the two groups. We found significant differences over time (6 months and 12 months; p = 0.002 for 5-mC% and p < 0.001 for global methylation by repeated measures ANOVA) but no differences due to treatment with AI or PBO (p = 0.12 for 5-mC% and p = 0.06 for global methylation), testosterone (p = 0.62 for 5-mC% and p = 0.21 for global methylation), or estradiol (p = 0.35 for 5-mC% and p = 0.92 for global methylation); and an independent difference due to weight (p = 0.02 for 5-mC% and p = 0.03 for global methylation). Since some of our patients were on metformin which has been known to affect methylation, we adjusted for the intake of the drug which did not show any difference between those on LSI + AI and LSI + PBO (Table 3).
3.4. Lifestyle Intervention Alters Expression of Adipogenic and Metabolic Genes
LSI ± AI significantly modulated the expression of genes associated with adipogenesis and metabolism in PBMCs in men with severe obesity and hypogonadism. LSI ± AI also decreased mRNA levels of adipogenic markers (Figure 4), PPARγ (BL: 1.53 ± 0.37 vs. 12 M: 0.83 ± 0.18; p = 0.08), and CEBPα (BL: 1.59 ± 0.32 vs. 12 M: 0.76 ± 0.18; p = 0.045). Similarly, FTO mRNA expression was significantly decreased after 12 months of LSI ± AI (BL: 1.47 ± 0.27 vs. 12 M: 0.86 ± 0.09; p = 0.028) (Figure 4). Notably, the direction of change was consistent across participants, and the coordinated downregulation of PPARγ, CEBPα, and FTO suggests a broad transcriptional response to the LSI ± AI.
4. Discussion
This study provides novel evidence that combined diet and exercise elicit significant epigenetic and transcriptional adaptations in severely obese hypogonadal men. We observed reductions in global DNA methylation, decreased expression of DNA methyltransferases (DNMT1, DNMT3A, and DNMT3B), and downregulation of key metabolic genes (PPARγ, CEBPα, and FTO) following 12 months of LSI ± AI. Our analysis also indicated that treatment with AI or PBO showed no influence on the changes in methylation. Thus, our findings suggest that the combination of diet and exercise reprograms the leukocyte methylome, which likely accompanies the metabolic improvement associated with LSI.
DNA methylation, catalyzed by DNMTs, regulates gene expression, genomic stability, and imprinting [7]. Hyperactivation of DNMTs has been linked to obesity, insulin resistance, and chronic inflammation [17]. The human genome encodes five DNMTs—DNMT1, DNMT2, DNMT3A, DNMT3B, and DNMT3L. Among these, DNMT1, DNMT3A, and DNMT3B are canonical cytosine-5 methyltransferases which contain an N-terminal regulatory region and a C-terminal catalytic domain that uses S-adenosylmethionine (SAM) as a methyl donor and a base-flipping mechanism to generate 5-methylcytosine [21], representing global methylation. DNMT1 preferentially methylates hemi-methylated DNA and is therefore considered the principal maintenance methyltransferase [22], whereas DNMT3A and DNMT3B primarily act on unmethylated DNA to establish de novo methylation; dysregulation of these enzymes affects adipose biology [23].
A prior study by Yang et al. suggested adipocyte differentiation involves large epigenomic changes (histone modifications, DNA methylation) in precursor cells (e.g., 3T3-L1) entering adipogenesis in lean murine adipocytes [24]. While other studies showed elevated DNMT3A expression in adipose tissue of transgenic mice [23]; these findings underscore the close association between DNMT activity and adipose tissue biology. Although not significantly different from baseline, our subjects experienced a reduction in body fat (total and visceral) with weight loss. The observed reduction in DNMT1, DNMT3A, and DNMT3B expression (Figure 2) after weight loss from diet and exercise suggests that LSI can downregulate methylation machinery, promoting a more transcriptionally permissive chromatin state. These findings are consistent with earlier studies showing exercise-induced hypomethylation [25] and decreased DNMT expression which may also involve other tissues and cells such as skeletal muscle and leukocytes [15].
Although human intervention studies have not yet directly demonstrated suppression of DNMT1, DNMT3A, or DNMT3B in metabolic tissues following lifestyle or exercise interventions, mechanistic evidence from animal and cellular models shows that reduced DNMT1 can drive passive demethylation [26], while decreased DNMT3A/3B limits de novo methylation [27]. Exercise-regulated DNMT dynamics in muscle and metabolic tissues have been observed in preclinical models [28], supporting our findings. Such changes can activate transcription of genes involved in mitochondrial biogenesis, oxidative metabolism, and anti-inflammatory responses—pathways central to the metabolic improvements associated with sustained physical activity [29].
Despite the emergence of effective weight loss drugs, exercise and dietary modifications remain important components in the management of obesity [30]. LSI not only improves body composition and glucose homeostasis but also remodels the epigenome [31]. Our data indicate that both global 5-mC and total DNA methylation levels were significantly reduced after LSI over 12 months, which was demonstrated only in the PBMCs in our study as we have no other tissues available. These changes occurred independent of AI use. Our observations agree with the results from the studies of Ronn et al, who analyzed subcutaneous adipose tissue biopsies from 23 previously sedentary, healthy men before and after 6 months of endurance exercise training. In this study, the authors observed widespread genome-wide methylation changes, with 17,975 CpG sites (across 7663 genes) significantly altered after training. Moreover, several loci with methylation changes also showed corresponding mRNA expression shifts, including key obesity and type 2 diabetes-related genes TCF7L2 and KCNQ1, demonstrating coordinated epigenetic and transcriptional remodeling in metabolic tissues [5]. Nitert et al. also showed the impact of an exercise intervention on genome-wide DNA methylation in human skeletal muscle of patients with diabetes [32].
Based on our findings, we hypothesize that the observed suppression of DNMT1, the primary maintenance methyltransferase, may promote passive loss of DNA methylation during cell turnover and concurrent reductions in DNMT3A and DNMT3B activity, enzymes responsible for de novo methylation, thus further limiting methylation as illustrated in Figure 5. Results from the genome-wide methylation studies of Benton et al. in human adipose tissue also showed differential methylation of obesity-associated genes including DNMT3A in obese women before and after gastric bypass [33].
Weight-loss interventions and reductions in inflammation were often accompanied by partial “reversal” of methylation changes, suggesting a link between inflammation driven by adiposity and epigenetic regulation [34]. In our study, we observed transcriptional downregulation of PPARγ, CEBPα, and FTO, highlighting the metabolic reprogramming induced by LSI. PPARγ and CEBPα are key adipogenic transcription factors that promote lipid accumulation and adipocyte differentiation [35] This suppression following 12 months of LSI indicates reduced adipogenic signaling which may enhance metabolic efficiency. Similar findings have been reported in exercise interventions, where PPARγ promoter demethylation correlated with metabolic adaptations in skeletal muscle [25]. The decline in FTO mRNA suggests normalization of the obesity-linked RNA demethylase pathway. FTO regulates energy expenditure and appetite through m6A RNA demethylation; its overexpression is associated with obesity and insulin resistance [36]. Downregulation of FTO following lifestyle modification may contribute to improved metabolic control and energy balance.
5. Limitations
This study is limited to using PBMCs as a surrogate for systemic epigenetic profiling. Although peripheral methylation patterns often reflect whole-body metabolic adaptations, tissue-specific analysis (e.g., skeletal muscle or adipose tissue) would provide stronger mechanistic evidence. However, these tissues are not easily obtainable in humans. This is the reason why some epigenetic studies [37,38] have relied on easily accessible surrogate tissues such as PBMCs. Accordingly, cellular heterogeneity within PBMC samples represents a limitation, and the observed methylation changes should be interpreted with caution and considered primarily as signals warranting further investigation using approaches that control for immune cell composition (e.g., cell sorting), rather than definitive conclusion. Nevertheless, our data provide supporting evidence for reduced methylation activity, as we observed decreased expression of DNMTs. In addition, this study included only severely obese hypogonadal men; therefore, our findings may not be generalizable to women, eugonadal or less obese populations. Moreover, our cohort size was small and not powered for subgroup analysis; thus, future studies should include larger populations to confirm differential responses. In addition, a locus-specific methylation analysis would have strengthened our data but was not performed in our study. Finally, half of our subjects were on aromatase inhibitors. However, there was no difference in methylation between those on aromatase inhibitors and those on placebo, suggesting that the changes in methylation patterns in the epigenome of our subjects are primarily due to LSI with contribution from the accompanying weight change.
6. Conclusions
In summary, weight loss from combined lifestyle therapy induces favorable epigenetic reprogramming in obese hypogonadal men, characterized by reduced global DNA methylation, downregulation of DNMTs, and suppression of adipogenic genes. Given the small number of subjects in our study, a study with a larger sample size, a longitudinal analysis beyond 12 months, and assessment of locus-specific methylation to assess the stability of these methylation changes and their relationship to long-term clinical outcomes is necessary to confirm our findings. Furthermore, future work integrating genome-wide methylation, transcriptomic, and metabolomic analyses could unravel specific pathways linking methylation to sustainable metabolic improvement.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bhupathiraju S.N. Hu F.B. Epidemiology of Obesity and Diabetes and Their Cardiovascular Complications Circ. Res.20161181723173510.1161/CIRCRESAHA.115.30682527230638 PMC 4887150 · doi ↗ · pubmed ↗
- 2Yuen M.M.A. Health Complications of Obesity: 224 Obesity-Associated Comorbidities from a Mechanistic Perspective Gastroenterol. Clin. N. Am.20235236338010.1016/j.gtc.2023.03.00637197879 · doi ↗ · pubmed ↗
- 3Ahmed F. Hetty S. Laterveer R. Surucu E.B. Mathioudaki A. Hornbrinck E. Patsoukaki V. Olausson J. Sundbom M. Svensson M.K. Altered Expression of Aromatase and Estrogen Receptors in Adipose Tissue from Men with Obesity or Type 2 Diabetes J. Clin. Endocrinol. Metab.2025110 e 3410 e 342410.1210/clinem/dgaf 03839833659 PMC 12448628 · doi ↗ · pubmed ↗
- 4Sandovici I. Morais T. Constância M. Monteiro M.P. Epigenetic Changes Associated with Obesity-related Metabolic Comorbidities J. Endocr. Soc.20259 bvaf 12910.1210/jendso/bvaf 12940862085 PMC 12375920 · doi ↗ · pubmed ↗
- 5Rönn T. Volkov P. Davegårdh C. Dayeh T. Hall E. Olsson A.H. Eriksson K.F. Groop L. Ling C. A six months exercise intervention influences the genome-wide DNA methylation pattern in human adipose tissue P Lo S Genet.20139 e 100357210.1371/journal.pgen.100357223825961 PMC 3694844 · doi ↗ · pubmed ↗
- 6Ling C. Rönn T. Epigenetics in Human Obesity and Type 2 Diabetes Cell Metab.2019291028104410.1016/j.cmet.2019.03.00930982733 PMC 6509280 · doi ↗ · pubmed ↗
- 7Moore L.D. Le T. Fan G. DNA Methylation and Its Basic Function Neuropsychopharmacology 201338233810.1038/npp.2012.11222781841 PMC 3521964 · doi ↗ · pubmed ↗
- 8Lyko F. The DNA methyltransferase family: A versatile toolkit for epigenetic regulation Nat. Rev. Genet.201819819210.1038/nrg.2017.8029033456 · doi ↗ · pubmed ↗