Maternal undernutrition reprograms reproductive and metabolic phenotypes in female offspring of the rabbit model
María Arias-Alvarez, Alfonso Gutiérrez-Adan, Eva Pericuesta, Pablo Bermejo-Alvarez, Pilar Millán, María Rodríguez, Pedro Luis Lorenzo, Pilar García Rebollar, Rosa María García-García

TL;DR
Maternal undernutrition in rabbits affects offspring's reproductive and metabolic health through gene expression changes in oocytes, even when re-feeding occurs.
Contribution
The study reveals that re-feeding after maternal undernutrition can reduce long-term effects on offspring growth and metabolism while modulating gene expression in oocytes.
Findings
Maternal food restriction increased serum aminotransferase levels in offspring, suggesting hepatic stress.
FR group oocytes showed upregulated genes related to oxidative stress and fatty acid uptake, while cumulus cells showed altered apoptosis-related gene expression.
Re-feeding after maternal undernutrition attenuated adverse effects on offspring growth and metabolism.
Abstract
Maternal undernutrition during pregnancy in mammals can induce long-term effects in offspring health through molecular programming of the gametes. Using the rabbit model, this study investigates whether maternal food restriction (FR) during the first two-thirds of gestation induces effects on ovarian follicular, oocyte, and early embryo developmental markers in F1 female offspring at the onset of reproductive life. Additionally, body composition, metabolic profile, and growth trajectory from birth to sexual maturity (16 weeks) were assessed to evaluate potential impacts on overall health. Pregnant females (F0) were fed either ad libitum (Control group) or a restricted diet covering 60% of nutritional requirements (FR group). Offspring from both groups were fed ad libitum. Maternal FR had no significant effects on birth weight and survival of progeny, growth trajectory, feed intake or…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
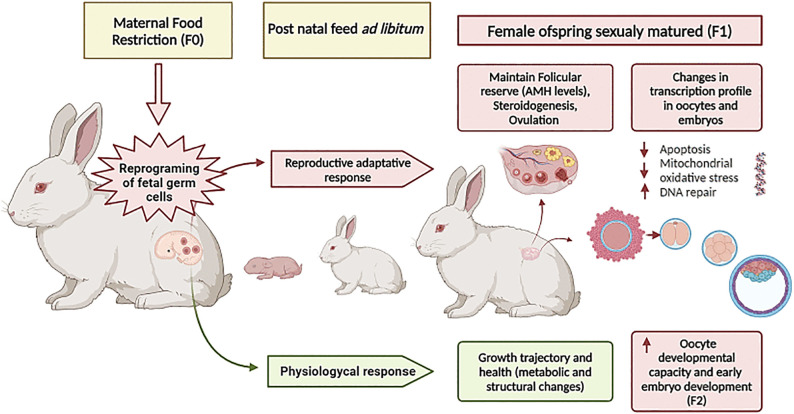 Fig 1
Fig 1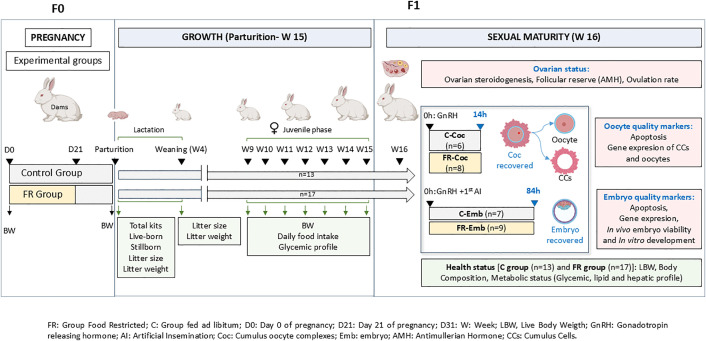 Fig 2
Fig 2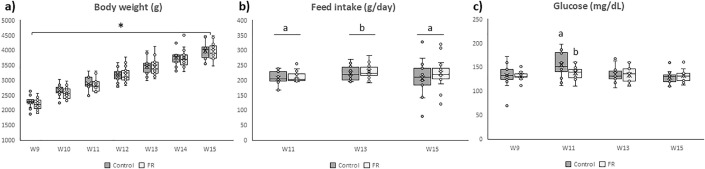 Fig 3
Fig 3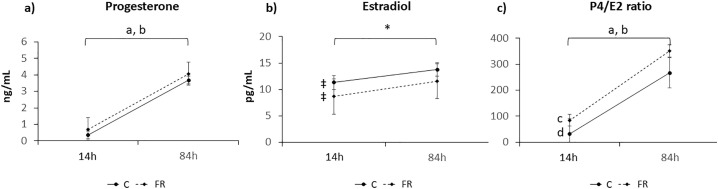 Fig 4
Fig 4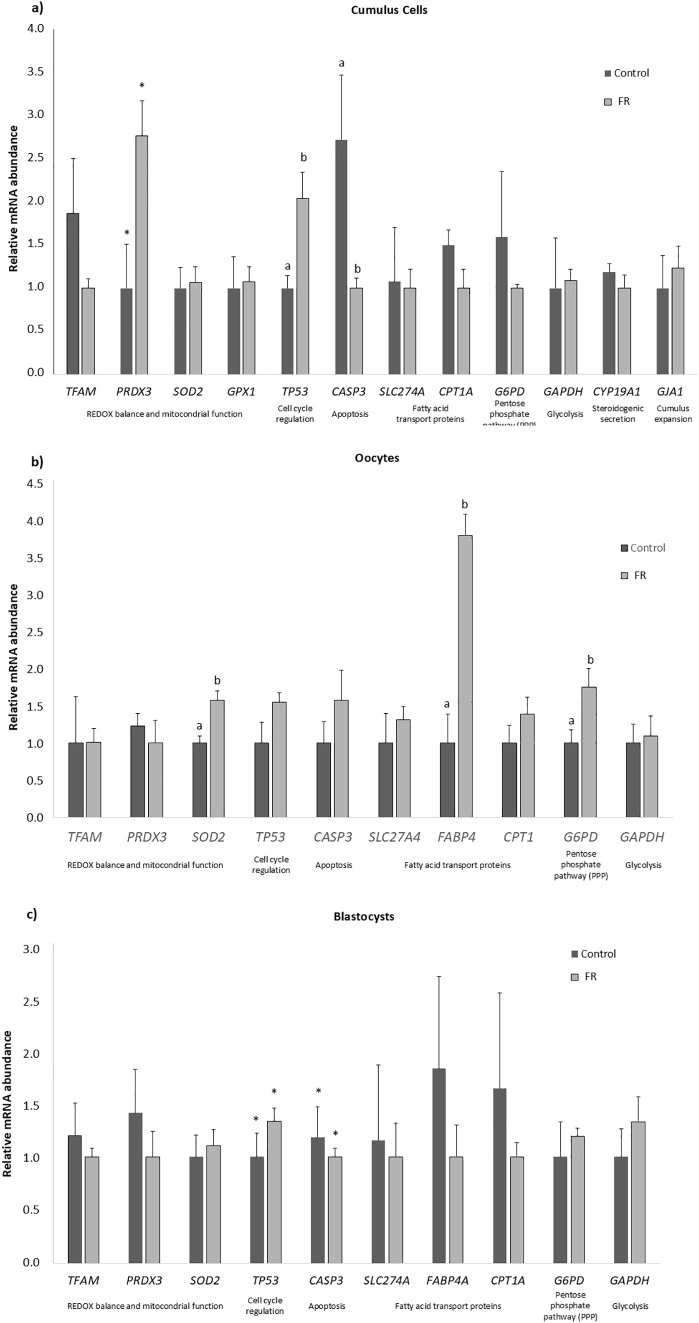 Fig 5
Fig 5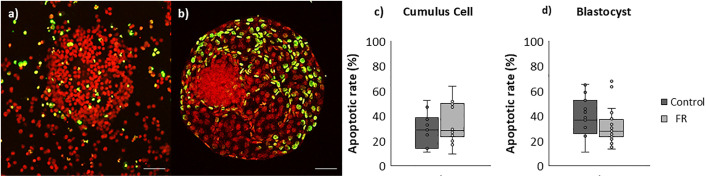 Fig 6
Fig 6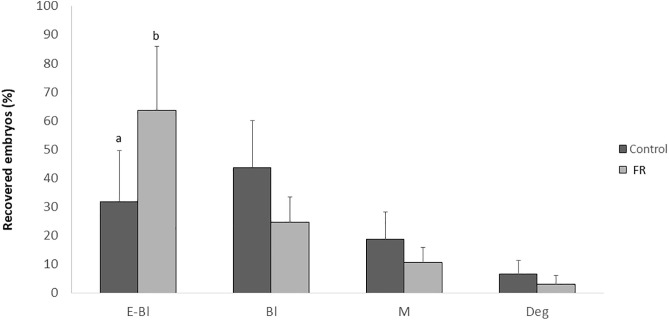 Fig 7
Fig 7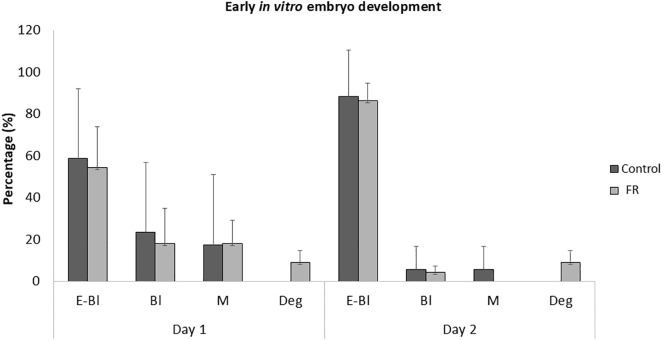 Fig 8
Fig 8- —Spanish Ministry of Science
- —Spanish Ministry of Science
- —Universidad Complutense Madrid
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBirth, Development, and Health · Gestational Diabetes Research and Management · Rabbits: Nutrition, Reproduction, Health
Introduction
According to the concept of Developmental Origins of Health and Disease (DOHaD), early life events can induce permanent structural and functional modifications in foetal organs preparing the organism for predicted postnatal conditions [1]. This framework is supported by extensive evidence showing that maternal nutrition during pregnancy plays critical role in organogenesis, and metabolic programming, thereby influencing the offspring’s long-term health and predisposing to chronic diseases such as obesity, and type 2 diabetes through epigenetic mechanisms [2–7]. The thrifty gene hypothesis proposed by James Neel [8] posits that in utero undernutrition activates adaptive genetic responses —such as genes involved in fat storage capacity, glycolysis or energy metabolism—beneficial to ensure fetal survival, but detrimental when nutrition is abundant in postnatal life [9].
The influence of the nutrition of women and adolescent girls of reproductive age on the health and reproductive potential of their progeny represents a critical public health concern [10]. Epidemiological studies of the Dutch winter famine, during which caloric intake was reduced by 60–70%, demonstrated that maternal malnutrition during pregnancy can induce long-term health consequences in the offspring, even in the absence of marked effects on birth weight [11]. Currently, an estimated 29.3% of the global population (approximately 2.3 billion people) experiences moderate to severe food insecurity [12], with an increase of 150 million people affected by hunger since the onset of the COVID-19 pandemic [13]. Calorie restriction diets -typically involving a reduction of 30–50% of daily maintenance requirements- are common among women of reproductive age in both high- and low-income countries, albeit driven by different socioeconomic and cultural factors.
Nutritional requirements vary throughout pregnancy, particularly during critical developmental periods [14], such as the formation and proliferation of fetal germ cells, which are particularly vulnerable to nutritional disturbances [5,15]. Epigenetic modifications to the genome that arise during fetal development in the germ line can persist in future generations by altering the gene expression in oocytes and therefore, affect fertility across F1 and F2 generations through their effects on oocyte competence and early embryo development [10,16]. Altered DNA methylation in F1 oocytes may transmit these metabolic disturbances to the next generation, as such epigenetic changes persist in adult F2 tissues [17]. Many studies in animal models, such as rats, mice, and humans, have suggested various epigenetic factors (e.g., DNA methylation, histone modification, and small RNAs) in germ cells may carry these memories [18,19]. The female offspring are more affected by in utero malnutrition than males [20]. As a consequence, some studies have demonstrated that acute fasting before ovulation compromises oocyte developmental potential in rabbits [21], while prolonged caloric restriction alters the transcriptomic profile in mice [22]. However, the effects of maternal food restriction during gestation on the transcriptional profile of oocytes from F1 females remain largely unexplored [23].
Prenatal maternal dietary restriction to 50% of nutrient requirements has been associated with impaired ovarian offspring development [2,24], reduced follicular reserve [25–28], altered follicular steroidogenic activity [29], disrupted ovulation, and increased follicular atresia in sheep, cow and rodents models [30]. Nevertheless, early-life adaptations leading to long-term reproductive dysfunctions may depend on several factors, including the timing and severity of the nutritional insult, as well as species-specific physiological responses [24,25,31–33]. Both the amount of food consumed, and the composition of the diet are important factors [34]. Interestingly, recent studies have challenged the prevailing notion of detrimental maternal food restriction effects, reporting no adverse consequences on ovarian and uterine function or fertility in heifers exposed to prenatal or postnatal nutritional interventions [35,36]. These findings are particularly relevant for elucidating the role of nutrition in reproductive health and may offer valuable translational models for both human and animal reproductive biology.
Ovarian oxidative stress is suggested as a possible mechanism to explain the changes in offspring reproduction in response to maternal undernutrition [25,29,37]. Oxidative stress induces a cascade of chain reactions that can ultimately lead to lipid peroxidation in membrane phospholipids, affecting the function and permeability of cell membranes, and generating DNA damage, meiotic arrest and mitochondrial dysfunction, which, in turn, activate caspase cascades and culminate in irreversible cell death [38]. Given that mitochondria are inherited exclusively through the maternal line, such alterations may compromise oocyte competence and early embryonic development [39] and negatively impact the fertility potential of female progeny [40]. Rabbits are a valuable translational model for human health research, particularly for investigating the mechanisms regulating fetal growth, developmental reprogramming, and their long-term physiological consequences [23,41–44]. Compared to rodents, rabbits exhibit higher biochemical and physiological similarity to humans [45,46], and are considered a more appropriate model for studying human lipid metabolism [47]. Furthermore, their genome is more closely related to that of humans [48], and their extensive historical use in toxicology has led to a well-characterized physiology [10,49,50]. As induced ovulators, rabbits also offer the advantage of precise control over the timing of ovulation, embryo development, and gestation. The use of artificial insemination (AI) further reduces variability by minimizing confounding paternal effects.
The main aim of this study was to test the hypothesis that 40% of food restriction (FR) during early and mid-pregnancy (covering 60% of nutritional requirements) exerts long-term effects on F1 female offspring, particularly on ovarian function (ovulation rate, follicular reserve, and steroidogenic activity), oocyte competence, and embryo development. We used a well-established rabbit model of maternal FR during the first two thirds of gestation [41–44,50], consistent with the definition of FR by the World Health Organization [13]. This period encompasses key developmental stages, including preimplantation (days 0–7), organogenesis and gonadal differentiation (days 14–16), and placental formation (days 10–17) [50]. Previous studies demonstrated that this model alters lipid metabolism during early to mid-gestation [41], while maintaining maternal energy homeostasis (glucose and insulin levels) and lipid metabolism (NEFA) at term. Compensatory feeding in late gestation mitigates maternal body weight and body reserve losses in this rabbit model [42]. To estimate ovarian reserve, serum anti-Müllerian hormone (AMH) concentrations were measured as a reliable biomarker. Specific target genes involved in metabolic processes (fatty acid uptake, transport and energy metabolism); redox balance and mitochondrial function, cell cycle regulation and apoptosis, meiotic progression, and Cumulus Cells (CCs) steroidogenesis and expansion were evaluated as potential markers of oocyte and embryo quality in F1 offspring at the onset of reproductive life (Fig 1).In addition, given the known long-term impact of maternal FR on offspring health and metabolism, we assessed postnatal and juvenile growth, feed intake, and metabolic parameters (glycemic, lipid, and hepatic profiles) and body composition in sexually mature F1 females to provide integrative insights into their reproductive performance.
Graphical abstract.Summary of the experimental design and key findings of this study.
Materials and methods
Unless otherwise stated, all chemicals were purchased from Merk (Spain).
Animals and experimental design
New Zealand White x California rabbit does (Oryctolagus cuniculus) were housed individually in flat-deck cages under conditions of constant light/darkness (16:8 h), a temperature of 20–25ºC and relative humidity of 60–75% using a forced ventilation system. Animals had free access to fresh water. This study was performed according to Spanish Policy for Animal Protection RD53/2013. All experimental procedures with animals were approved by the Animal Ethics Committee of the Madrid Polytechnic University (UPM, Spain) and approved by the competent authority in animal welfare (code of project: PROEX 302/15) and complied with Spanish guidelines for the care and use of animals in research, according to European Union Regulation 2010/63/UE.
Maternal food restriction model (F0 generation)
Multiparous females of 6 months of age (F0) were fed ad libitum a commercial diet that meet the nutritional requirements of rabbits, especially regarding calories and protein content (2400 kcal/kg of digestible energy, 16% crude protein, 37% crude fibre, and 3.7% fat, NANTA, Madrid, Spain). Diet was formulated following the nutritional recommendations for breeding does by de Blas and Mateos [51]. There was only one diet, provided in different quantities for the experimental groups. Feed intake was recorded daily and established in a mean of 175 g per animal and day. The animals did not have access to bedding as an additional nutrient source during the period of FR.
All F0 females were artificially inseminated (AI) using a pool of fresh diluted semen with more than 20 million spermatozoa per dose in 0.5 mL of a commercial diluent (commercial extender MA-24 Ovejero, León, Spain). Ovulation was induced by intramuscular injection of 20 μg gonadotropin-releasing hormone (Inducel-GnRH, Ovejero, León, Spain). At this stage, females were randomly assigned to two experimental groups: the control group (C, n = 72) which received the standard diet ad libitum throughout pregnancy, and the food restriction group (FR, n = 70), which received a quantitatively restricted diet (Fig 2). The FR females were fed 60% of their previous daily food intake (105 g/day) until day 21 of pregnancy, followed by ad libitum feeding for one week more until the end of pregnancy (gestation length is usually 31 days) according to our previous experimental design of moderate maternal FR [42–44]. A pregnancy diagnosis was made by abdominal palpation. Maternal body weight (BW) from 11 C and 15 FR pregnant does randomly chosen was recorded on the day of the AI and at the end of pregnancy.
Experimental design.Pregnant females (F0) were fed ad libitum all through the gestation period (group C) or were feed restricted (105 g/day) from Day 0 (artificial insemination, AI) to Day 21 (group FR) of gestation (gestation length is 31 days). In female offspring (F1), sampling was specially focused when does initiate their reproductive life (16 weeks). The factors examined were body weight (BW), body composition and metabolic status. After induced ovulation with GnRH analogue treatment, ovarian status and, oocyte and embryo quality markers were examined 14 h later (Coc groups) or 84 h after ovulation induction and AI (Emb groups). Also, offspring performance was assessed at parturition, weaning (week 4) and from week 9 to 15 (juvenile phase). Female offspring were fed ad libitum all their postnatal life. FR: feed restriction group; C: control ad libitum feeding group; D0: Day 0 of pregnancy; D21: Day 21 of pregnancy; D31: Day 31 of pregnancy (delivery); LBW, live body weight; GnRH: gonadotropin-releasing hormone; AI: artificial insemination; Coc: cumulus oocyte complex; Emb: embryo; AMH: antimullerian hormone; CCs: cumulus cells.
Female offspring (F1 generation)
At birth, the total number of kits, number of live-born, and stillborn were recorded from 41 C and 49 FR females. The number of kits per litter was adjusted from 7 to 12at the beginning of lactation. Litter size and total litter weight were recorded both at parturition and weaning (4 weeks of age). Offspring were continuously fed ad libitum with the same control diet as their F0 dams until reaching sexual maturity at 16 weeks of age. From 9 weeks to 15 weeks of age (juvenile stage), a total of 13 C and 17 FR daughters (F1 generation) were randomly chosen from F0 dams belonging to control and FR group. These F1 females were housed in individual cages, and their BW was monitored weekly, while feed intake and glycemic profile were assessed every two weeks. When F1 females reached sexual maturity (at 16 weeks of age), BW, body composition (BC), blood metabolic profile (glycemic, lipid and hepatic markers), ovarian steroidogenic response (estradiol and progesterone) and estimated ovarian follicular reserve by measuring serum levels of Anti-Müllerian hormone (AMH) were assessed. Blood samples were obtained from the ear vein in tubes with anticoagulant, centrifuged for 15 min at 700 g at 4 ºC, and plasma was stored at −80 ºC until analysis. All samples were collected at 09:00 h to avoid the effects of circadian variations.
F1 females of each experimental group (C and FR) were randomly divided into two subgroups: 1) Group Coc, for the study of cumulus-oocyte complex (COC) quality markers (subgroups C-Coc, n = 6, and FR-Coc, n = 8) and, 2) Group Emb, to assess embryo quality markers and in vivo and in vitro embryo developmental capacity (subgroups C-Emb, n = 7 and FR-Emb, n = 9). In Group Coc, ovulation was induced by the intramuscular injection of GnRH (20 μg; Inducel-GnRH; Ovejero) (as described before for F0 females). In the Emb groups, ovulation was also induced, and AI was performed simultaneously, as previously described for the F0 females. Reproductive tracts were obtained by mid-ventral laparotomy after sedation with 35 mg/kg of ketamine (Imalgene; Merial), and euthanasia using an intravenous bolus of barbiturate (30 mg/kg; Dolethal; Vetoquinol). Ovulated COCs were recovered by flushing the reproductive tract 14h after in both experimental groups (C-Coc and FR-Coc) [52]. Embryos were recovered 84h after GnRH administration + AI in both experimental groups (C-Emb and FR-Emb) [53]. Oviducts were flushed with phosphate-buffered saline (PBS) supplemented with 0.1% BSA (w/v). The rates recorded were ovulation rate (number of corpora lutea (CL)/ doe), COC recovery rate (COCs/ number of recorded CL x 100), and embryo recovery rate (embryos/ CL counts x 100). Embryo morphology was assessed immediately after their recovery. Expanded blastocysts were fixed in 4% paraformaldehyde for the apoptosis study, and the embryo cell counting. Early blastocysts were immediately snap-frozen in liquid nitrogen and storage at −80 °C for transcript abundance analysis by quantitative real-time polymerase chain reaction (qRT-PCR). The remaining morphologically viable embryos were in vitro cultured (IVC) to establish their developmental potential [54]. Embryos were pooled per individual F1 female and the animal was considered the experimental unit for gene expression analyses.
Estimation of body composition
The BC was determined at week 16 in F1 females by bioelectrical impedance analysis (BIA). A four-terminal body composition analyser (Model Quantum II, RJL Systems, Detroit, MI, USA) was used to determine bioelectrical resistance and reactance (both in ohms) according to [55]. Impedance, which represents the opposition encountered by the electrical current as it passes through the animal’s body, was then calculated as follows:
Impedance = (resistance² + reactance²)^½^. The regression equations described by [56] were then used to estimate water, protein, ash, fat, and energy contents in relation to BW, parity number, and physiological status of the rabbit.
Evaluation of metabolic and hormonal status
Glycemic, lipid and hepatic profiling.
We assessed serum parameters related to the offspring’s long-term health that can affect reproduction, including glycemic metabolism (glucose, insulin, and glucose/insulin ratio), lipid metabolism (total cholesterol, triglycerides (TG) and TG/cholesterol ratio), and hepatic profile (aminotransferase (AST), serum alanine transaminase (ALT) and AST/ALT ratio). All ratios between two metabolites were calculated by dividing the serum concentrations of one metabolite by that of the other for each animal, and then the mean value was calculated for each experimental group.
Carbohydrate/lipid metabolism and hepatic variables were measured using commercial kits (Biolabo, Maizy, France) in a clinical biochemical analyser (Tecom Science Co., Jiangxi, China) according to the procedures described by [42].
Plasma insulin concentrations were determined with a rabbit insulin ELISA kit (Mercodia AB, Uppsala, Sweden). The assay sensitivity of this method was 0.15 mUI/L, the intra-assay variation coefficient was 3.9% and inter-assay variance was 5.30%. Insulin resistance was identified through homeostasis model assessment (HOMA-IR) according to [33] and calculated using the equation [insulin (mU/L) × (glucose (mg/dL)/18)]/ 22.5.
Anti-müllerian hormone levels.
AMH concentrations were determined in duplicate samples using a specific rabbit AMH competitive ELISA kit (MyBioSource, San Diego, CA, USA). Absorbances were measured using an automatic plate reader (Atat Fax 3200, Awareness Technology INC) at 450 nm. Intra-assay and inter-assay coefficients of variation were below 10%. The detection limit after adjusting the standard curve to rabbit values was 0.1 ng/mL.
Ovarian steroidogenic response.
Serum estradiol (E2) and progesterone (P4) concentrations were analysed in duplicate samples using a competitive ELISA kit (DE4299 and DE1561, Demeditec Diagnostics GmbH, Kiel, Germany), as previously reported [54]. Absorbances were measured using an automatic plate reader (Atat Fax 3200, Awareness Technology INC) at 450 nm. Intra-assay and inter-assay coefficients of variation were 6.3% and 7.6% for E2, and 6.4% and 6.6% for P4, respectively. Detection limits were 1.4 pg/mL (E2) and 0.045 ng/mL (P4).
Quantitative gene expression study in cumulus oocyte complexes and blastocysts
RNA extraction and reverse transcription.
For transcript abundance analysis, we used 41 and 56 COCs (C-Coc and FR-Coc, respectively), and 32 and 40 early blastocysts (C-Emb and FR-Emb, respectively). CCs were mechanically removed by gentle repeated pipetting in phosphate buffered saline (PBS) + 1 mg/mL polyvinylpyrrolidone (PVP) medium. Groups of 10–12 blastocyst and 10–12 oocytes and their corresponding CCs were analysed separately. Pools of blastocysts, oocytes and CCs were placed in different 1.5 mL Eppendorf tubes, snap-frozen in liquid nitrogen, and stored at −80ºC until analysis. For RNA extraction and reverse transcription, poly (A) RNA was prepared as described previously [52,57,58] using the Dynabeads mRNA Direct Extraction KIT (Dynal Biotech, Thermo Fisher Scientific, Waltham, MA, USA) according to the manufacturer’s instructions with minor modifications. The reverse transcription (RT) reaction (Bioline, Thermo Fisher Scientific) was performed using poly (T) primer, random primers, and murine Moloney leukemia virus reverse transcriptase enzyme (M-MLV High-performance Reverse Transcriptase; Epicentre Biotechnologies, Thermo Fisher Scientific) in a total volume of 40 mL to generate cDNA. First, tubes were heated at 70ºC for 5 min to denature the secondary RNA structure, and then the RT was completed with the addition of 100 units of Superscript M-MLV RT enzyme. The samples were subsequently incubated at 42ºC for 60 min to allow for RT of RNA, followed by incubation at 70ºC for 10 min to denature the RT enzyme.
Quantitative real-time polymerase chain reaction.
Messenger RNA transcripts were quantified through qRT-PCR. Experiments were conducted to determine levels of each transcript relative to that of the housekeeping gene H2AFZ (histone family member z) previously validated for rabbit oocytes [52,59] and embryos [21,53,57,60]. All primers were designed using Primer–BLAST software (http://www.ncbi.nlm.nih.gov/tools/primersblast) to span exon-exon boundaries when possible. All qPCR reactions were performed in duplicate in a Rotorgene 6000 RealTime Cycler (Corbett Research Ltd, Cambridge, UK) after the addition of 2 µL samples to the PCR mix (GoTaq qPCR Master Mix; Promega Thermo Fisher Scientific) containing specific primers to amplify the housekeeping gene; genes related to important events in oocyte maturation and early embryo development in rabbits and markers of quality based on previous studies [21,52,53,57,59,60], namely TFAM, PRDX3, SOD2 (mitochondrial function and redox balance), G6PD (defence against oxidizing agents and meiotic progression), TP53 and CASP3 (cell cycle regulation and apoptosis related pathways), SLC274A, FABP4, CPT1 (fatty acid uptake and transport proteins), GAPDH (glycolysis) were used. In addition, GJA1 and CYP19A1 were quantified only in CCs because their expression is related to cell-to-cell communication for cumulus expansion in the COC, and steroidogenic secretion, respectively, and therefore they are indirect markers of oocyte competence. Primer sequences, approximate sizes of the amplified fragments, and melting temperatures for all transcripts are provided in Table 1. Cycling conditions were 94ºC for 3 min, followed by 35 cycles of 94ºC for 15 s, 56ºC for 30 s, 72ºC for 10 s, and 10 s at the corresponding melting temperature for each primer before fluorescence acquisition. Each pair of primers was previously validated [21,52,53,57,59,60] and optimized to achieve efficiencies close to 1. The comparative cycle threshold (CT) method was then used to quantify expression levels as described by [61]. To avoid primer-dimer artefacts, fluorescence was acquired in each cycle at a temperature higher than the melting temperature of primer dimers (specific for each product; 76–86ºC). Next, the threshold cycle or the cycle during the log-linear phase of the reaction at which fluorescence increased above the background was determined for each sample. Quantification was normalized against that of the endogenous control H2AFZ. According to the comparative CT method, the ΔCT value was determined by subtracting the H2AFZ CT value for each sample from each gene’s CT value. To calculate ΔΔCT, we used the highest sample ΔCT value as an arbitrary constant to subtract from all other ΔCT sample values. Fold changes in the relative gene expression of the target were determined using the formula 2^-ΔΔCT^.
Table 1: Primers used to amplify genes of interest in rabbit oocytes, cumulus cells, and blastocysts by quantitative reverse transcription–polymerase chain reaction.
Apoptosis assessment in cumulus oocyte complexes and blastocysts, and quantification of embryo cell number
The subgroups of COCs (C-Coc, n = 11; FR-Coc, n = 14) and blastocyst (C-Emb, n = 16; FR-Emb, n = 27) were pretreated with 20 mg/mL proteinase K working solution for 1 h in a humidified dark chamber at 37ºC. Strand breaks in DNA were detected by the terminal deoxyribonucleotidyl transferase-mediated dUTP–digoxigenin nick end-labelling method (TUNEL; In Situ Cell Death Detection Kit, POD; Roche Diagnostics) following a protocol adapted from [52,59]. Positive control sections were treated with DNAse I for 10 min at room temperature (RT) in a humidified chamber before incubation for the TUNEL reaction. As a negative control, samples were incubated with the label solution of the TUNEL reaction mixture without the enzyme solution. To avoid RNA interference, COCs and embryos were treated with RNAses before staining. Finally, they were counterstained with 0.25 mg/mL propidium iodide for 15 min and mounted between a coverslip and glass slide in mounting solution (ProLong Gold Antifade Reagent, Invitrogen, Waltham, MA, USA). Samples were observed under a laser scanning confocal microscope (Leica TCS SP2, Wetzlar, Germany) using a 488-nm excitation laser to visualize TUNEL-positive cells, and a 546-nm excitation laser to assess red fluorescence. Format, laser, gain, and offset were kept constants for all samples. Images were taken every 5 µm in z axis and then analysed using ImageJ software (http://rsbweb.nih.gov/ij/). The apoptosis index was calculated as the ratio of green area (TUNEL positive) divided by the red area (PI-positive) x100. Maximal projection was obtained with all sections taken in each COC to visualize the whole apoptotic area. In red nuclei (PI-positive) in embryos were quantified using the FindStackMaxima macro, which automates the counting process. Embryo quality was determined as the total number of cells per blastocysts within each experimental group.
In vivo and in vitro embryo developmental competence
Embryo development was assessed by examining morphology immediately after recovery following the International Embryo Transfer Society (IETS) guidelines and according to their developmental stage (https://www.iets.org/) using a stereomicroscope (SMZ 800, Nikon, Japan). Early and expanded blastocyst were morphologically classified based on blastocoel expansion and following the guidelines described by Bo and Mapletof [62]. Rates of recovered blastocysts and degenerated or retarded embryos in vivo (embryos whose stage of development did not correspond to time after ovulation, i.e.,: zygotes, 2–16 cell embryos, and early morulae) were calculated relative to the total number of recovered embryos.
The ability of recovered embryos to develop in vitro was assessed by in vitro culture (IVC). Only morphologically good quality embryos recovered (n = 17, in C group and n = 22 in FR group) were cultured in groups (5–10 embryos/group) in TCM-199 supplemented with 10% foetal calf serum (FCS) at 38ºC under a 5% O_2_ and 5% CO_2_ atmosphere in air at maximum humidity according to a procedure described elsewhere [52,59]. Embryo developmental competence was determined by assessing their development to the expanded blastocyst stage in 48 h under a stereomicroscope (SMZ800; Nikon, Tokyo, Japan), as previously reported [54]. The embryo viability rate after IVC was obtained by dividing the number of blastocysts by the number of cultured embryos.
Statistical analysis
Data were analysed using SAS software (9.4, Enterprise edition, SAS) and SPSS Version 19 (IBM SPSS Statistics). The Shapiro–Wilks test was used to assess normality and homogeneity of variance.
In dams (F0), evolution of body weight was assessed by repeated measures analysis (MIXED procedure). The model included the time (AI and birth) and their interaction as main effects. The dam was considered the experimental unit.
In litters (F1), total number of kits at parturition (live + stillborn), number of live-born kits, number of stillborn kits, litter weight at birth, as well as litter size and weight at weaning (number of kits), litter weight at weaning, were analysed by GLM procedure with FR as the main source of variation. The number of kits per litter, ranging from 7 to 12, was included as a covariate at the beginning of lactation. The litter was considered the experimental unit.
In the juvenile stage (F1), the evolution of BW in F1 offspring, feed intake and blood glucose concentrations were assessed by repeated measures analysis (MIXED procedure). The model included the FR, the age of the rabbits and their interaction as main effects. Each daughter (F1) was considered the experimental unit.
At 16 weeks old (F1), BW, estimated BC, serum metabolic and endocrine parameters, and number of CL (ovulation rate) were analysed as a completely randomized design, with FR as the main source of variation, using the GLM procedure. To compare serum steroidogenic hormones between experimental groups and at both time points (14 h and 84 h after ovulation), a Wilcoxon test for non-parametric samples was used. Each daughter (F1) was considered the experimental unit.
For apoptosis rate, embryo cell count, and mRNA transcript expression, an analysis of variance was performed considering only the FR as the main effect. Morphological categorization of recovered embryos between experimental groups and in vitro early embryo development was compared using Wilcoxon’s rank sums test. To assess in vitro embryo development over time, a Friedman test was performed.
Grubbs’ test was used to determine if the most extreme values were significant outliers (GraphPad). All means were compared using a protected t-test. Differences were considered significant at P < 0.05 and a trend when P < 0.10. Results are presented as least squared mean (ls means S.E.M.).
Results
Prenatal fed restriction did not affect the body weight of F0 females
Similar BW was recorded between control and FR F0 females at the time of AI (3.90 ± 0.12 kg and 4.04 ± 0.08 kg for C and FR groups, respectively) and parturition (4.62 ± 0.12 kg and 4.60 ± 0.08 kg for C and FR groups, respectively), although significant differences were observed from AI to parturition in both experimental groups (P < 0.05).
Prenatal feed restriction did not impact offspring performance at parturition, weaning and juvenile period
No significant differences were observed between the C and FR groups in the total number of live kits at parturition (11.4 ± 0.7 and 11.1 ± 0.6) and number of live-born kits (10.5 ± 0.8 and 10.7 ± 0.7). However, a trend toward a higher number of stillborns was observed in the C group (0.9 ± 0.3 and 0.3 ± 0.3; P = 0.0542). Litter weight at birth tended to be lower in the C group compared to FR (584.9 ± 21.5 g and 636.3 ± 18.8 g), although this difference was not statistically significant (P = 0.07).
At weaning, both litter weight (6218.3 ± 195.0 g and 5899.0 ± 166.4 g) and litter size (9.9 ± 0.2 and 9.8 ± 0.1 kits) were comparable between the C and FR groups. From week 9 to week 15 (juvenile phase) the BW of F1 females increased significantly as expected in both groups (P < 0.0001) without significant differences between them (Fig. 3a). Average daily feed intake was comparable between groups (Fig 3b). However, a significant effect of time was detected (P < 0.05), with increased intake observed at week 13 in both groups. The interaction between group and time was not statistically significant.
Body weight, feed intake, and blood glucose levels during the juvenile phase of female offspring (F1).Data correspond to daughters of dams fed ad libitum (C group, n = 13) or subjected to moderate food restriction during the first two-thirds of gestation (FR group, n = 17). Body weight (a) and feed intake (b) were recorded weekly from 9 to 15 weeks of age, and blood glucose (c) was measured biweekly. Each dot represents one animal; lines indicate least squares means ± SEM. Different superscripts indicate significant differences between groups [a, b (P < 0.05)] or among time points [# (P < 0.0001)].
Glycemic profile did not differ significantly between groups (Fig 3c). Nevertheless, a significant increase in glucose levels was observed in the C group at week 11 compared to FR one (P < 0.001).
Prenatal fed restriction altered hepatic enzyme levels in F1 females at the onset of their reproductive life, without affecting body composition, or glycemic and lipid profiles
At 16 weeks, females of both experimental groups reached similar mean BW and estimated BC values (Table 2).
Table 2: Body weight (BW) and estimated body composition recorded in F1 sexually mature rabbit females (16 weeks old) born to dams subjected to moderate feed restriction (FR group) for 3 weeks or fed ad libitum (C group) throughout pregnancy. After birth, all offspring were fed ad libitum.
No significant effect of maternal FR was observed on serum parameters related to glucose and lipid metabolism. However, hepatic enzyme ALT was significantly higher in F1 females from restricted mothers (P < 0.02). The ratio AST/ALT tended to be higher in FR group (P = 0.09) (Table 3).
Table 3: Metabolic serum parameters and hepatic enzyme levels of sexually matured F1 rabbit females (16 weeks old) born to dams subjected to moderate feed restriction (FR group) for 3 weeks or fed ad libitum (C group) during pregnancy at ovulation time point.
Prenatal feed restriction did not alter AMH levels, ovulation rate, or the overall ovarian steroidogenic response, except for an increased P4/E2 ratio at ovulation in F1 females
Mean serum AMH concentrations were similar in the C and FR groups at 14 h post-GnRH (0.50 ± 0.15 and 0.76 ± 0.53 ng/mL, respectively) and 84 h post-AI (0.54 ± 0.28 and 0.55 ± 0.20 ng/mL, respectively).
Mean number of CL per animal was similar in C and FR (13.00 ± 2.82 and 11.5 ± 3.81, respectively). As depicted in Fig 4a, serum P4 concentrations significantly increased (P < 0.001) from 14 h to 84 h post-GnRH, but no significant differences were observed between the two experimental groups at both time points examined (when COCs or embryos were recovered). E2 tended to be higher in C than FR group (P = 0.06, Fig 4b) at 14 h yet was similar in both groups at 84 h post-GnRH. However, the ratio P4/E2 was significantly higher in FR compared to C (P < 0.05) at 14 h (ovulation time), but no differences were found at the time of embryo recovery (84h) (Fig 4c). In both experimental groups, this ratio increased from 14 h to 84 h after ovulation (P < 0.001).
Steroid hormone concentrations in female offspring (F1).Serum levels of (a) progesterone (P4), (b) estradiol (E2), and (c) the progesterone-to-estradiol ratio (P4/E2) were measured in 16-week-old female offspring of dams subjected to moderate food restriction during the first two-thirds of gestation (FR group) or fed ad libitum throughout pregnancy (C group). Number of does: C group, 14 h (n = 6) and 84 h (n = 7); FR group, 14 h (n = 8) and at 84 h (n = 9) after ovulation. Data are presented as mean ± SEM. Different superscripts indicate significant differences [a,b (P < 0.001)] or a trend [() P = 0.07] between time points (14 h vs. 84 h after ovulation), and [c,d (P < 0.05)], or a trend [‡ (P = 0.06)] between experimental groups.*
Prenatal fed restriction modulates transcript abundance of oxidative stress and apoptosis-related genes in cumulus cells, oocytes and embryos in F1 females
Transcript abundances in oocytes, their corresponding CCs, and blastocysts are shown in Figs 5a- 5c, respectively. In oocytes, SOD2, FABP4, and G6PD poly(A) mRNA expression levels were significantly upregulated in the FR group (P < 0.05). No significant differences were observed in the other transcripts analysed. In CCs, TP53 mRNA expression was significantly upregulated in FR (P < 0.05), whereas CASP3 mRNA expression was significantly lower in FR-CCs compared to those from group C (P < 0.05). PRDX3 mRNA expression tended to be higher in FR compared to C (P = 0.06). No significant differences were observed in the other transcripts analysed. Blastocyst-mRNA abundances of the examined transcripts were similar in the two experimental groups. In blastocysts derived from FR group, TP53 gene expression tended to be upregulated (P = 0.09), and CASP3 gene expression downregulated (P = 0.09).
Relative mRNA abundance of target genes in (a) oocytes, (b) cumulus cells (CCs), and (c) blastocysts from female offspring (F1).Samples were obtained from 16-week-old female offspring (F1) of dams subjected to moderate food restriction during the first two-thirds of gestation (FR group) or fed ad libitum throughout pregnancy (C group). Group C: n = 41 oocytes with their corresponding CCs, and 32 blastocysts; Group FR: n = 56 oocytes with their corresponding CCs, and 40 blastocysts. Data are presented as the mean ± SEM. Bars with different superscripts indicate significant differences between C and FR groups for each gen [a, b (P < 0.05)] or a trend [() P = 0.07].*
Prenatal fed restriction improved in vivo blastocyst expansion rates and embryo cell count in F1 females
The COC recovery rate at 14 h post-GnRH was comparable between groups (94.87 ± 25.39% and 86.95 ± 24.73%, in group C and FR, respectively). Apoptosis percentages were similar in CCs from C and FR oocytes at ovulation (32.96 ± 14.70% and 28.80 ± 14.30%, respectively) (Fig 6).
Representative images of apoptosis in cumulus-oocyte complexes (COCs),and expanded blastocysts from female offspring.(a) COCs and (b) expanded blastocysts from F1 females. Apoptosis was evaluated by the TUNEL assay. Confocal images (x40) show maximal projection of a complete COCs (a) and blastocysts (b) illustrating the overlap between nuclei stained with propidium iodide (red) and TUNEL-positive apoptotic cells (green). Graphs (c) and (d) show the apoptotic rate in cumulus cells and blastocysts, respectively, from daughters of dams fed ad libitum (C group) or subjected to moderate food restriction during the first two-thirds of gestation (FR group). COCs: C (n = 11), FR (n = 14); embryos: C (n = 16), FR (n = 27). Each dot represents one COC or embryo; lines indicate least squares means ± SEM.
The embryo recovery rate did not differ significantly between treatments (81.69 ± 28.62% and 83.87 ± 20.07%, in C and FR, respectively). Likewise, percentages of recovered blastocysts, morulae, unfertilized oocytes, and degenerated embryos were comparable across groups. However, group FR showed a significantly higher number of expanded blastocysts compared to group C (P < 0.005; Fig 7). Embryo cell counts were significantly higher in FR than C group (3218.5 ± 640.28 and 1101.0 ± 270.17 cells, P < 0.05). Apoptosis rate did not significantly vary between the groups (39.49 ± 15.9% and 32.40 ± 17.9%, groups C and FR, respectively) (Fig 6).
In vivo embryo development.Embryos were recovered 84h after GnRH administration and artificial insemination in 16-week-old female offspring (F1) of dams subjected to moderate food restriction during the first two-thirds of gestation (FR, group; n = 9) or fed ad libitum throughout pregnancy (C group, n = 7). The total number of embryos analysed was 58 in the C group and 78 in the FR group. Developmental stages were classified as expanded blastocysts (E-Bl), early blastocysts (Bl), morulae (M), or retarded/degenerate embryos (Deg). Data presented as mean ± SEM. Different superscripts indicate significant differences between experimental groups [a, b (P < 0.005)].
As shown in Fig 8, embryo development following 48 h of in vitro culture was similar in C and FR groups.
In vitro embryo development.Rabbit embryos recovered from 16-week-old female offspring (F1) of dams subjected to moderate food restriction during the first two-thirds of gestation (FR, group) or fed ad libitum throughout pregnancy (C group) were cultured in vitro for 48h. The number of embryos cultured was 17 in the C group and 22 in the FR group. Developmental stages were classified as morulae (M), early blastocysts (Bl), expanded blastocysts (E-Bl), or degenerated embryos (Deg). Data are presented as mean ± SEM.
Discussion
In high-income countries, women frequently adopt dietary restriction strategies to lose weight, which may inadvertently continue into early pregnancy. Such in utero nutritional imbalances -particularly maternal FR- can adversely affect the reproductive potential of the offspring. Emerging evidence highlights the influence of maternal nutrition on oocyte quality in F1 offspring, providing insights into the mechanisms that govern F2 oocyte development and their potential consequences for subsequent reproductive performance. In the present study, animals were subjected to moderate FR during the first two-thirds of gestation, at levels comparable to those observed in both human and animal dietary regimens [63]. In rabbits, this gestational window encompasses critical developmental stages, including preimplantation embryogenesis (0–7 days) and organogenesis (7–18/19 days) [64]. Specifically, first germ cells can be found from day 9 of pregnancy and the gonads are evident from day 14 [50]. Our experimental model demonstrates that moderate maternal FR during this period modulates the expression of genes associated with oxidative stress, apoptosis and fatty acid uptake in oocytes of the F1 female offspring. These findings suggest that FR more than induced adverse effects exerts a potential adaptative response by the activation of compensatory intracellular responses that may support early embryonic development in juvenile females at the onset of their reproductive life.
Although both maternal overnutrition and undernutrition during gestation have been shown to adversely affect offspring growth and metabolic function [65,66], the level of FR applied in this study did not result in in overt detrimental effects on birth weight, survival or postnatal growth. Previous studies have shown that maternal FR during early gestation exerts a limited impact on these parameters in cows [28], although it may still induce metabolic alterations in the offspring [67]. In contrast, nutrient restriction during late gestation has been consistently linked to reduced birth weight in several species, including rats [68], cattle [69,70], and sheep [71]. This increased vulnerability is likely related to the exponential phase of fetal growth during late gestation, when nutrient demands are maximal and sensitivity to dietary restriction perturbations is heightened [72].
In the present study, the restoration of adequate maternal intake during the final third of gestation may may have enabled compensatory fetal growth in the FR group, resulting in birth and weaning outcomes comparable to those of control offspring [42]. In rabbits,moderate and transient FR (30%), during pregnancy has been shown to alter hormonal and metabolic parameters associated with adult disease risk, such as hyperglycaemia, insulin resistance, and increase in thyroxine levels, mirroring alterations described in humans [33]. Notably, when FR is limited to early or mid-gestation, these parameters return to physiological levels by late pregnancy, suggesting the activation of adaptive maternal and fetal responses.
Similarly, studies using a comparable FR model,as applied in the current work, have reported normalized maternal glycaemic and lipid profiles at term, despite the presence of hyperinsulinemia, insulin resistance, and hypertriglyceridemia in near-term fetuses [42]. These fetal metabolic changes may represent early adaptive responses to a nutrient-limited intrauterine environment, although they could also confer increased susceptibility to metabolic disorders such as fatty liver disease, type 2 diabetes, and obesity later in life [9]. In the present study, no significant alterations were observed in growth, feed intake, or glycemic profile of female offspring during juvenile period. Likewise, at the onset of reproductive life, BW, BC, and glycemic and lipid profiles, remained comparable between groups.
These findings are consistent with previous studies in juvenile sheep exposed to maternal undernutrition during early to mid-gestation [73], cows nutritionally restricted during the first trimester [28], mice exposed to a periconceptional protein-restricted environment [9], and rabbits undergoing 50% dietary restriction during gestation [74], all of which described preserved postnatal growth and general health in offspring. Collectively, these data support the notion that adequate postnatal nutrition may mitigate or compensate for prenatal nutritional challenges, thereby facilitating the normalization of key physiological functions [35,43,74,75].
Nevertheless, the detection of elevated serum levels of ALT in F1 female offspring of FR dams suggests the presence of subtle but persistent hepatic alterations [76–79] that were not accompanied by overt growth or metabolic disturbances during early and juvenile life.The prenatal environment also plays a key role in shaping adult ovarian folliculogenesis and steroidogenesis. AMH is widely recognized as a reliable indirect marker of ovarian reserve, as its levels correlate with the number of growing follicles across species, including rodents [80], humans [81], cows [28,82], dogs, guinea pigs, and horses [83]. In rabbits, AMH levels have been shown to correlate particularly well with the number of secondary and antral follicles [84]. In our study, AMH levels were comparable in the offspring of control and fed-restricted mothers, consistent with previous reports [85]. By contrast, studies in other species have demonstrated that maternal FR to 50% during gestation can reduce AMH levels in young adult offspring [30], indicating a compromised ovarian follicular reserve [25–27,37,86]. These interspecies discrepancies may be attributed to differences in the timing of ovarian reserve establishment. Unlike species such as the mice, where primordial follicle formation begins around days 12–13 of gestation, rabbits initiate folliculogenesis exclusively after birth [87]. Thus, maternal FR limited to early and mid-gestation is unlikely to directly impair ovarian reserve formation of F1 female rabbits. In contrast, in species such as sheep and cattle, maternal undernutrition during early gestation has been shown to delay fetal follicle formation and reduce the number of growing follicles in female offspring [24,28], despite minimal effects on prenatal or postnatal growth were evidenced [88]
Our findings also suggest that prenatal FR did not impair the ovarian response to treatment with a short-acting GnRH analogue. The number of COCs retrieved was similar between groups, and serum progesterone (P4) concentrations after ovulation and at the early post-ovulatory window (up to 84 hours) remained within physiological ranges, indicating functional corpora lutea able of supporting early embryo development [89]. Although an early rise in P4 at the time of GnRH administration can potentially shift the endometrial receptivity window and disrupt embryo-maternal synchrony [90,91] (leading to reduced implantation rates and poorer pregnancy outcomes), this effect was not evident in our model. Despite not measuring LH levels directly, the absence of a premature P4 increase 14 hours post-GnRH supports the conclusion that premature luteinization did not occur, consistent with previous observations [92]. Interestingly, an elevated P4/E2 ratio was detected at ovulation in offspring of undernourished mothers. In clinical reproductive contexts, this ratio has been proposed as a more reliable marker of pregnancy outcomes than P4 levels alone [93–95], although the supporting evidence remains inconclusive and thresholds vary depending on the GnRH protocol applied [92,96,97]. Moreover, species-specific differences must be considered, as rabbits are induced ovulators, in contrast to humans, which may limit the extrapolation of these findings. In our study, the elevated P4/E2 ratio in F1 females from FR group did not compromise luteal function nor negatively impact embryonic development. On the contrary, embryos from the FR group exhibited more advanced developmental stages, with a higher proportion reaching the expanded blastocyst stage at the time of recovery, an indicator of greater developmental competence [98]. These findings suggest the possible involvement of adaptive or compensatory mechanisms, either intrinsic to the oocyte or mediated by the oviductal microenvironment, that may confer a developmental advantage. Nevertheless, in the absence of significant differences in gene expression profiles and given the similar in vitro development observed under standardized culture conditions, the underlying mechanisms remain unclear and warrant further investigation.
Although less investigated, maternal undernutrition has also been linked to altered expression of key ovarian steroidogenic enzymes, such as P450SCC and aromatase, which are essential for sex steroid synthesis in the offspring ovary [99–101]. In our study, although serum E2 levels were slightly higher in the control group, no significant differences were observed, neither in hormone concentrations nor in CYP19A1 expression in CCs. These results are consistent with those of Maia et al. [35] who reported no significant changes in serum P4 or E2 concentrations following in utero nutritional restriction in cattle. Conversely, other studies have reported disrupted ovarian steroidogenesis [29], reduced CL number [24,30], and decreased plasma P4 levels in adult F1 females [2,102]. Collectively, these outcomes may reflect differences in the severity, duration, and type of maternal nutritional restriction, particularly in models involving protein deprivation rather than global caloric restriction [26,30,37].
At the molecular level, maternal FR induced transcriptional changes in genes associated with cell cycle regulation, apoptosis, and oxidative stress within the COCs of female offspring. TP53, often referred to as “the guardian of the genome,” orchestrates cell cycle arrest and apoptosis via mitochondrial caspases. It regulates an appropriate cellular response to various stress signals [103]. Specifically, TP53 activation in preovulatory mouse granulose cells impairs ovulation and fertilization [104] and promotes apoptosis in macaque granulosa cells [105]. In the present study, TP53 was upregulated in CCs from FR group, suggesting a potential response to prenatal stress signals. Notably, this increase was accompanied by downregulation of its effector, CASP3which may indicate a protective or compensatory adaptation rather than the activation of apoptotic pathways [106]. Together with the similar expression genes related to CCs expansion (CJA1), steroidogenic activity (CYP19A1), ovulation rate and apoptotic rate determined using the TUNEL assay in both groups, these findings suggest that maternal FR did not exert a detrimental effect on F2-COCs functionality. This observation aligns with earlier findings showing increased TP53 and CASP3 expression in in vitro matured oocytes, reflecting enhanced DNA repair and apoptotic regulation compared to in vivo matured COCs [52]. Moreover, the ancestral function of TP53 is linked to ensure fecundity and embryo viability rather than tumour suppression [103], supporting the involvement in the acquisition of developmental competence [107] and the selective elimination of damaged oocytes and embryos [108], instead of acting merely as an inductor of embryo loss [109,110].
Oxidative stress is a recognized mediator of long-term physiological changes induced by early-life nutritional insults [92,108,109]. In oocytes, SOD2 is involved in oxidative responses and mitochondrial activity, as it encodes a mitochondrial protein that catalyses the dismutation of superoxide into H_2_O_2_. Its upregulation in oocytes from the FR group indicates activation of redox homeostasis mechanisms, potentially contributing to the preservation of mitochondrial function and biogenesis. The unchanged expression of PRDX3 and TFAM mRNAs in both groups further supports this notion. Notably, mRNA levels of TP53 and CASP3 in oocytes were also similar between groups, indicating a balance between oxidative stress management and apoptotic signalling, in agreement with previous findings in rabbit oocytes [52]. PRDX3 catalyses the reduction of mitochondrial H₂O₂ and it is considered a marker of mitochondrial oxidative stress and apoptosis [110,111]. In our study, the slightly increased expression of PRDX3 in CCs from the FR group may reflect the activation of antioxidant defence. PRDX3 upregulation has previously been associated with increased mitochondrial DNA copy number, enhanced CCs survival, and improved oocyte developmental competence to the blastocyst stage in gilts [112].
Additionally, G6PD expression was upregulated in oocytes from the FR group. G6PD is a cytosolic enzyme that catalyzes the first rate-limiting step of the pentose phosphate pathway (PPP), which primarily generates NADP, a key electron donor for antioxidant defence and reductive biosynthetic reactions. Increased G6PD expression may therefore enhanced protection against oxidative damage, supporting meiotic progression and developmental potential to reach the blastocyst stage [113]. Overall, these data suggest that the antioxidant defence mechanisms in oocytes of F1 females remain functionally intact under prenatal FR conditions. Nevertheless, findings regarding G6PD expression and oocyte competence appears to be species-specific, as controversial results have been reported in rats [113], sheep [114], rabbits [52], and cattle [115]. Finally, fatty acid binding protein 4 (FABP4), a cytoplasmic protein involved in fatty acid uptake, transport, and metabolism was upregulated in oocytes from the FR group. FABP4 is found in adipocytes and granulosa cells during follicular atresia in lambs [102] and mice [116]. Its upregulation may reflect prenatal metabolic programming of lipid homeostasis. While such adaptations could be under conditions of limited nutrient availability, they may also predispose offspring to metabolic disorders later in life when nutrient supply is restored (as in the present model), potentially even affecting the F2 generation. To our knowledge, this is the first report describing altered FABP4 expression in oocytes in the context of prenatal undernutrition therefore the interpretation of this result requires prudence. The limited sample size of F1 females that achieved the onset of reproductive life and the focus on a selected set of candidate genes, although biologically relevant, do not capture the full spectrum of molecular responses to maternal undernutrition. Further studies with larger cohorts and functional analyses beyond gene expression are needed to elucidate how these genes influence reproductive outcomes and health in following generation.
Conclusions
Our results indicate that the reproductive phenotype of female offspring exposed to maternal FR during first two thirds of gestation may be modulated through changes in oocyte gene expression, particularly in pathways related to oxidative stress, apoptosis, and lipid metabolism. Importantly, the observed molecular changes are more consistent with manifestations of developmental plasticity than with impairment per se, and appear to function predominantly as compensatory mechanisms, that contribute to the preservation of oocyte competence and early embryo development in F1 females exposed to suboptimal prenatal conditions. Nevertheless, these findings should be interpreted cautiously, as the specific causal mechanism remain to be elucidated. Re-feeding at the end of pregnancy after maternal FR appears to mitigate long-term adverse effects on offspring health indicators such as growth, body composition and glycaemic and lipid metabolism but, the detection of liver damage biomarkers suggests subtle physiological impacts. Overall, these results underscore the intrinsic plasticity of female offspring to prenatal nutritional challenges and highlight the need for further research to elucidate how early-life molecular programming influences fertility and health outcomes in subsequent generations.
Supporting information
S1 DataRaw data 1.(XLSX)
S2 DataRaw data 2.(XLSX)
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Barker DJP. The origins of the developmental origins theory. J Intern Med. 2007;261(5):412–7. doi: 10.1111/j.1365-2796.2007.01809.x 17444880 · doi ↗ · pubmed ↗
- 2Sloboda DM, Howie GJ, Pleasants A, Gluckman PD, Vickers MH. Pre- and postnatal nutritional histories influence reproductive maturation and ovarian function in the rat. P Lo S One. 2009;4(8):e 6744. doi: 10.1371/journal.pone.0006744 19707592 PMC 2727050 · doi ↗ · pubmed ↗
- 3Godfrey KM, Gluckman PD, Hanson MA. Developmental origins of metabolic disease: life course and intergenerational perspectives. Trends Endocrinol Metab. 2010;21(4):199–205. doi: 10.1016/j.tem.2009.12.008 20080045 · doi ↗ · pubmed ↗
- 4Gluckman PD, Hanson MA, Cooper C, Thornburg KL. Effect of in utero and early-life conditions on adult health and disease. N Engl J Med. 2008;359(1):61–73. doi: 10.1056/NEJ Mra 0708473 18596274 PMC 3923653 · doi ↗ · pubmed ↗
- 5Gluckman PD, Hanson MA, Pinal C. The developmental origins of adult disease. Matern Child Nutr. 2005;1(3):130–41. doi: 10.1111/j.1740-8709.2005.00020.x 16881892 PMC 6860944 · doi ↗ · pubmed ↗
- 6Roseboom T, de Rooij S, Painter R. The Dutch famine and its long-term consequences for adult health. Early Hum Dev. 2006;82(8):485–91. doi: 10.1016/j.earlhumdev.2006.07.001 16876341 · doi ↗ · pubmed ↗
- 7Tobi EW, Slieker RC, Luijk R, Dekkers KF, Stein AD, Xu KM, et al. DNA methylation as a mediator of the association between prenatal adversity and risk factors for metabolic disease in adulthood. Sci Adv. 2018;4:eaao 4364. doi: 10.1126/sciadv.aao 4364 PMC 579222329399631 · doi ↗ · pubmed ↗
- 8Neel JV. Diabetes mellitus: a “thrifty” genotype rendered detrimental by “progress”?. Am J Hum Genet. 1962;14(4):353–62. 13937884 PMC 1932342 · pubmed ↗