Revisiting the link between NADPH oxidase p22phox C242T polymorphism and ischemic stroke risk: an updated meta-analysis
Yan Wang, Li An

TL;DR
This study finds no significant link between a genetic variation in NADPH oxidase p22phox and the risk of ischemic stroke.
Contribution
An updated meta-analysis clarifies the inconsistent findings regarding the C242T polymorphism and ischemic stroke risk.
Findings
No significant association was found between the C242T polymorphism and ischemic stroke susceptibility.
Subgroup analyses by ethnicity and stroke type also showed no significant risk increase.
Sensitivity analysis confirmed the stability of the results.
Abstract
The relationship between the NADPH oxidase p22phox C242T and susceptibility to ischemic stroke (IS) has been extensively studied, yet the findings from these studies remain inconsistent. To clarify this association, we conducted an updated meta-analysis of case-control studies. A comprehensive literature search of PubMed, Embase, and CNKI was performed on January 31, 2025. Studies exhibiting Hardy-Weinberg equilibrium (HWE) deviations in the control population were excluded from analysis. Pooled odds ratios (ORs) with corresponding 95 % confidence intervals (CIs) were calculated to evaluate the association across various genetic models. Our meta-analysis included 10 studies, comprising 3,422 cases and 3,410 controls. We found no significant association between the C242T polymorphism and IS susceptibility under any genetic model. Subgroup analyses stratified by ethnicity and IS type…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
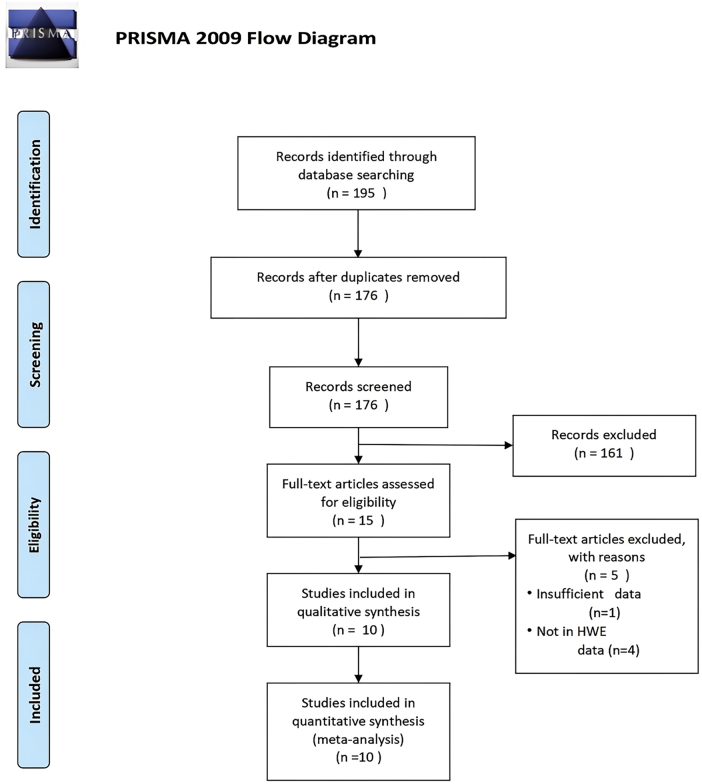 Figure 1
Figure 1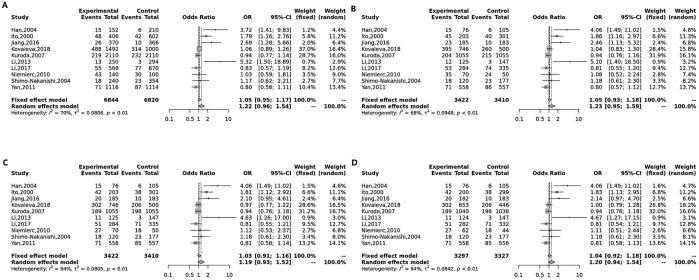 Figure 2
Figure 2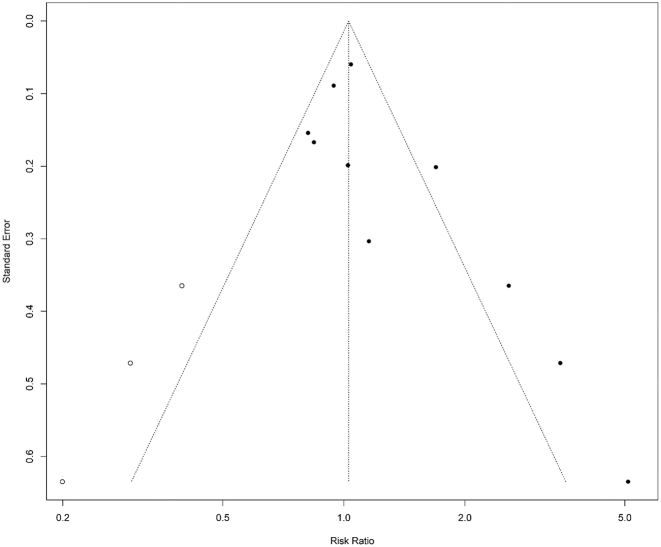 Figure 3
Figure 3| First author, Year of publication | Country | Ethnicity | Sample size | Age (Mean ± SD, y) | Subtype of IS | ||
|---|---|---|---|---|---|---|---|
| Case | Control | Case | Control | ||||
| Han [ | China | East Asian | 76 | 105 | 64.92 ± 11.1 | 63.91 ± 11.9 | SVO and LAA |
| Ito [ | Japan | East Asian | 203 | 301 | 58 ± 8 | 59 ± 4 | SVO and LAA |
| Jiang [ | China | East Asian | 185 | 183 | 66.31 ± 10.82 | 67.91 ± 12.27 | SVO and LAA |
| Kovaleva [ | Russia | Caucasian | 746 | 500 | 67.89 ± 0.42 | 54.42 ± 0.55 | SVO and LAA |
| Kuroda [ | Japan | East Asian | 1,055 | 1,055 | 70 ± 10 | 70 ± 10 | SVO, LAA, CE and SUE |
| Li [ | China | East Asian | 125 | 147 | 65.42 ± 11.65 | 65.22 ± 9.69 | SVO and LAA |
| Li [ | China | East Asian | 284 | 335 | 61.46 ± 10.52 | 60.17 ± 9.56 | SVO and LAA |
| Niemierc [ | Poland | Caucasian | 70 | 50 | 8.48 ± 5.44 | 9.0 ± 6.1 | SVO, LAA and SUE |
| Shimo-Nakanishi [ | Japan | East Asian | 120 | 177 | 61.2 ± 11.4 | 58.9 ± 9.25 | SVO and LAA |
| Yan [ | China | East Asian | 558 | 557 | 61.0 ± 9.8 | 62.2 ± 9.3 | SVO and LAA |
| Study | Distribution of p22phox C242T genotype | Frequency of p22phox C242T alleles | HWE | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| CC | CT | TT | C | T | |||||||
| Cases | Controls | Cases | Controls | Cases | Controls | Cases | Controls | Cases | Controls | ||
| Han [ | 61 | 99 | 15 | 6 | 0 | 0 | 137 | 204 | 15 | 6 | 0.763 |
| Ito [ | 158 | 261 | 42 | 38 | 3 | 2 | 358 | 560 | 48 | 42 | 0.634 |
| Jiang [ | 162 | 173 | 20 | 10 | 3 | 0 | 344 | 356 | 26 | 10 | 0.704 |
| Kovaleva [ | 351 | 240 | 302 | 206 | 93 | 54 | 1,004 | 686 | 488 | 314 | 0.329 |
| Kuroda [ | 851 | 840 | 189 | 198 | 15 | 17 | 1,891 | 1,878 | 219 | 232 | 0.181 |
| Li [ | 113 | 144 | 11 | 3 | 1 | 0 | 237 | 291 | 13 | 3 | 0.900 |
| Li [ | 231 | 261 | 51 | 71 | 2 | 3 | 513 | 593 | 55 | 77 | 0.444 |
| Niemierc [ | 35 | 26 | 27 | 18 | 8 | 6 | 97 | 70 | 43 | 30 | 0.312 |
| Shimo-Nakanishi [ | 102 | 154 | 18 | 23 | 0 | 0 | 222 | 331 | 18 | 23 | 0.355 |
| Yan [ | 487 | 471 | 71 | 85 | 0 | 1 | 1,045 | 1,027 | 71 | 87 | 0.158 |
| Population | OR | I2 (%) |
|
| ||||
|---|---|---|---|---|---|---|---|---|
| Fixed effects (95 % CI) |
| Random effects (95 % CI) |
| |||||
| Allele contrast (T vs. C) | All | 1.05 [0.95–1.17] | 0.35 | 1.22 [0.96–1.54] | 0.10 | 70.43 | 0.0004 | 0.0352 |
| Caucasian | 1.06 [0.90–1.25] | 0.49 | 1.06 [0.90–1.25] | 0.49 | 0 | 0.9298 | NA | |
| East Asian | 1.05 [0.91–1.20] | 0.51 | 1.37 [0.97–1.93] | 0.07 | 76.98 | 0.0001 | 0.0173 | |
| Japanese | 1.05 [0.89–1.25] | 0.55 | 1.22 [0.78–1.89] | 0.38 | 72.03 | 0.028 | 0.4829 | |
| Chinese | 1.03 [0.83–1.29] | 0.76 | 1.70 [0.90–3.20] | 0.10 | 82.79 | 0.0001 | 0.0023 | |
| Adult | 1.05 [0.95–1.17] | 0.35 | 1.25 [0.97–1.61] | 0.09 | 73.71 | 0.0002 | 0.0347 | |
| LAA | 1.03 [0.85–1.25] | 0.75 | 1.07 [0.80–1.44] | 0.15 | 43.83 | 0.1485 | 0.5224 | |
| SVO | 1.08 [0.88–1.31] | 0.47 | 1.11 [0.75–1.63] | 0.61 | 61.38 | 0.051 | 0.8318 | |
| Dominant model (TT + TC vs. CC) | All | 1.05 [0.93–1.18] | 0.45 | 1.23 [0.95–1.59] | 0.11 | 68.47 | 0.0008 | 0.0188 |
| Caucasian | 1.05 [0.84–1.29] | 0.71 | 1.05 [0.84–1.29] | 0.71 | 0 | 0.9139 | NA | |
| East Asian | 1.05 [0.90–1.22] | 0.51 | 1.22 [0.96–1.54] | 0.10 | 75.47 | 0.0002 | 0.0165 | |
| Japanese | 1.06 [0.88–1.28] | 0.52 | 1.23 [0.78–1.96] | 0.37 | 70.93 | 0.0321 | 0.4755 | |
| Chinese | 1.03 [0.82–1.30] | 0.79 | 1.67 [0.88–3.17] | 0.12 | 81.49 | 0.0002 | 0.0022 | |
| Adult | 1.05 [0.93–1.18] | 0.46 | 1.26 [0.95–1.65] | 0.10 | 71.97 | 0.0004 | 0.0169 | |
| LAA | 1.03 [0.83–1.27] | 0.82 | 1.10 [0.77–1.56] | 0.61 | 53.83 | 0.0897 | 0.4429 | |
| SVO | 1.09 [0.88–1.35] | 0.43 | 1.11 [0.75–1.65] | 0.60 | 57.36 | 0.0708 | 0.8637 | |
| Overdominant model (TC vs. TT + CC) | All | 1.03 [0.91–1.16] | 0.67 | 1.19 [0.93–1.52] | 0.16 | 64.06 | 0.0029 | 0.0121 |
| Caucasian | 0.98 [0.79–1.22] | 0.87 | 0.98 [0.79–1.22] | 0.87 | 0 | 0.7277 | NA | |
| East Asian | 1.05 [0.90–1.22] | 0.53 | 1.32 [0.94–1.84] | 0.11 | 71.65 | 0.0009 | 0.0163 | |
| Japanese | 1.07 [0.88–1.29] | 0.51 | 1.22 [0.79–1.88] | 0.37 | 65.88 | 0.0533 | 0.467 | |
| Chinese | 1.02 [0.81–1.30] | 0.86 | 1.57 [0.86–2.88] | 0.14 | 78.67 | 0.0009 | 0.0026 | |
| Adult | 1.03 [0.90–1.16] | 0.70 | 1.21 [0.93–1.57] | 0.16 | 67.99 | 0.0016 | 0.0115 | |
| LAA | 1.02 [0.82–1.26] | 0.88 | 1.11 [0.76–1.62] | 0.60 | 59.22 | 0.0614 | 0.3985 | |
| SVO | 1.10 [0.88–1.37] | 0.41 | 1.10 [0.77–1.58] | 0.59 | 48.46 | 0.1207 | 0.9248 | |
| Codominant model (TC vs. CC) | All | 1.04 [0.92–1.18] | 0.56 | 1.20 [0.94–1.54] | 0.15 | 68.47 | 0.0004 | 0.014 |
| Caucasian | 1.01 [0.80–1.27] | 0.92 | 1.01 [0.80–1.27] | 0.92 | 0 | 0.8002 | NA | |
| East Asian | 1.05 [0.90–1.22] | 0.53 | 1.32 [0.94–1.86] | 0.10 | 72.23 | 0.0007 | 0.0162 | |
| Japanese | 1.07 [0.88–1.29] | 0.51 | 1.22 [0.79–1.90] | 0.37 | 67.18 | 0.0475 | 0.4683 | |
| Chinese | 1.02 [0.81–1.29] | 0.86 | 1.58 [0.86–2.91] | 0.14 | 78.98 | 0.0008 | 0.0026 | |
| Adult | 1.04 [0.91–1.18] | 0.59 | 1.22 [0.96–1.54] | 0.14 | 70.43 | 0.0004 | 0.0352 | |
| LAA | 1.02 [0.82–1.26] | 0.86 | 1.11 [0.76–1.61] | 0.60 | 58.41 | 0.0654 | 0.4036 | |
| SVO | 1.10 [0.88–1.37] | 0.41 | 1.11 [0.77–1.60] | 0.59 | 50.22 | 0.1103 | 0.908 | |
- —Jiangsu Provincial Research Projects on Geriatric Health
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsNeutrophil, Myeloperoxidase and Oxidative Mechanisms · Nitric Oxide and Endothelin Effects · Renin-Angiotensin System Studies
Introduction
1
Ischemic stroke (IS) is a complex multifactorial disorder resulting from the interplay of genetic predispositions and environmental factors such as hypertension, diabetes, hyperlipidemia, and smoking [1]. As a major global public health concern, IS imposes a substantial burden on families, communities, and healthcare systems. Genetic factors are increasingly recognized as key contributors to IS susceptibility, with several gene polymorphisms implicated in its pathogenesis [2], [3], [4]. Atherosclerosis, a primary cause of ischemic cerebrovascular disease, involves accelerated lipid peroxidation driven by elevated levels of reactive oxygen species (ROS) [5], [6], [7], [8]. A major enzymatic source of ROS is the nicotinamide adenine dinucleotide phosphate (NADPH) oxidase complex [9].
The NADPH oxidase complex, which includes membrane subunits such as gp91phox (NOX2) and p22phox, as well as cytosolic components like p47phox, p67phox, p40phox, and Rac2, is a key source of ROS production [10], [11], [12]. The CYBA gene, which encodes the p22phox protein, is located on the long arm of chromosome 16 and exhibits several allelic variants [13], [14], [15]. One notable variant is the C242T polymorphism, which involves an amino acid substitution (His/Tyr) at residue 72, causing significant morphological changes in the extracellular loop of the p22phox protein. This polymorphism may disrupt the interaction between p22phox and the catalytic subunit NOX2, potentially reducing ROS production by NADPH oxidase [16].
In the past few decades, numerous studies have investigated the relationship between the p22phox C242T polymorphism and the risk of ischemic cerebrovascular disease [17], [18], [19]. However, results have been inconsistent, with some studies suggesting an association and others reporting none [20], [21], [22]. This inconsistency is further complicated by potential population stratification and deviations from Hardy-Weinberg equilibrium (HWE) in control groups. Therefore, the objective of this updated meta-analysis was to quantitatively synthesize the most recent evidence to reassess the association between the NADPH oxidase p22phox C242T polymorphism and ischemic stroke risk.
Materials and methods
2
Literature search
2.1
We conducted an extensive electronic search of relevant studies in PubMed, Embase, and the China National Knowledge Infrastructure (CNKI) up to January 2025. Search keywords included “NADPH,” “p22phox,” “C242T,” “rs4673,” “polymorphism,” and “ischemic stroke.” Our aim was to identify all eligible case-control studies exploring the association between the p22phox C242T polymorphism and ischemic stroke (IS) susceptibility.
Selection criteria and quality assessment
2.2
Studies were included if they met the following criteria: (1) case-control design evaluating the association between the p22phox C242T polymorphism and IS susceptibility; and (2) provision of sufficient data on genotype and allele frequencies. Studies were excluded if: (1) control groups deviated from Hardy-Weinberg equilibrium (HWE); or (2) they were duplicate publications. Maintaining HWE in control groups is essential for minimizing genotype errors and preventing population stratification biases. Two authors independently assessed the quality of included articles using the Newcastle-Ottawa Scale, which encompasses eight key criteria: adequate case determination, representativeness of cases, control selection, control determination, comparability between cases and controls, exposure ascertainment, uniform ascertainment methods for both groups, and non-response rate [23].
Data extraction
2.3
For each study, we extracted the following information: first author’s surname, publication year, ethnic group, country of origin, sample size, IS subtype, and genotype frequency. This detailed data extraction enabled us to compile comprehensive information for our meta-analysis.
Statistical analysis
2.4
Given that some studies reported zero frequency for the TT genotype, we applied four genetic models for analysis: the allelic model (T allele vs. C allele), the dominant model (TT + TC vs. CC), the overdominant model (TC vs. TT + CC), and the codominant model (TC vs. CC). We calculated odds ratios (ORs) and 95 % confidence intervals (CIs) from each study to assess the association between the p22phox C242T polymorphism and IS susceptibility. Heterogeneity across studies was evaluated using the chi-square test and I^2^ statistic. Meta-regression was performed using the method of moments to identify sources of heterogeneity. Both fixed-effects and random-effects models were applied and compared to ensure robustness. Subgroup analyses were conducted based on IS subtypes and ethnicity to explore potential differences. Sensitivity analysis was performed by sequentially removing individual studies to assess the stability of the pooled results. Publication bias was evaluated using funnel plots and Egger’s regression test. Statistical analyses were conducted using MetaGenyo, a web-based tool for meta-analysis of genetic association studies (https://metagenyo.genyo.es/). If publication bias was detected, the trim-and-fill method was applied using the R package ‘meta’ to assess whether this bias affected the combined effect size results.
Results
3
Literature search and study characteristics
3.1
Our comprehensive search across PubMed, Embase, and CNKI identified 195 articles. After screening titles and abstracts, we selected 15 studies for detailed evaluation [17], [18], [19, [24], [25], [26], [27], [28], [29], [30], [31], [32], [33], [34], [35]. Upon thorough review, five articles were excluded based on predefined criteria [17], [18], [19, 24], 32] (Supplementary data). Specifically, one study lacked sufficient genotype and allele frequency data [17], while four others deviated from HWE in their control groups [18], 19], 24], 32]. Ultimately, 10 studies were included in our meta-analysis, encompassing 3,422 cases and 3,410 controls [25], [26], [27], [28], [29], [30], [31, [33], [34], [35] (Figure 1). The characteristics of these studies are summarized in Table 1, and the detailed distribution of alleles and genotypes is presented in Table 2. Following rigorous quality assessment, all included studies were rated as medium or high quality, with scores ranging from 5 to 9.
Flow of studies through the review process.
Meta-analysis findings
3.2
Our meta-analysis revealed no significant association between the p22phox C242T polymorphism and ischemic stroke susceptibility in any genetic model. Specifically, the allelic model (T vs. C) showed an odds ratio (OR) of 1.22 with a 95 % confidence interval (CI) of 0.96–1.54 (p = 0.11). The dominant model (TT + TC vs. CC) yielded an OR of 1.23 (95 % CI: 0.95–1.59, p = 0.11), while the overdominant model (TC vs. TT + CC) had an OR of 1.19 (95 % CI: 0.93–1.52, p = 0.17). The codominant model (TC vs. CC) resulted in an OR of 1.20 (95 % CI: 0.94–1.54, p = 0.15). These results are visually represented in Figure 2.
Forest plot analysis evaluating the association between NADPH oxidase p22phox C242T polymorphism and ischemic stroke (IS) risk. The analysis includes four genetic models: (A) Allelic model comparing the T allele to the C allele; (B) dominant model combining TT and TC genotypes against CC genotype; (C) overdominant model comparing TC genotype to the combined TT and CC genotypes; (D) codominant model comparing TC genotype to CC genotype.
Heterogeneity and subgroup analyses
3.3
Significant heterogeneity was observed across all genetic models in the overall analysis, with I^2^ statistics exceeding 50 % and p-values below 0.05. Subsequent meta-regression analysis failed to identify any significant influence of ethnicity or IS type on IS risk. Subgroup analyses stratified by ethnicity and IS subtype similarly revealed no significant associations under any genetic model (Table 3). Notably, heterogeneity markedly decreased in studies focusing on small-vessel occlusion and large-artery atherosclerosis (LAA) and no heterogeneity was observed in Caucasians (Supplementary data), suggesting that these factors may have distinct genetic influences.
Sensitivity and publication bias analyses
3.4
Sensitivity analysis, performed by sequentially removing individual studies, indicated that our findings were robust, with no single study significantly influencing the overall results. Publication bias was assessed using funnel plots and Egger’s regression test. While publication bias was detected in all genetic models (p-values ranging from 0.012 to 0.035), it was absent in studies of small-vessel occlusion and large-artery atherosclerosis. The results remained unchanged after adjusting for publication bias using the trim-and-fill method in the allelic model (OR 1.027, 95 % CI 0.693–1.523; p = 0.892; Figure 3).
The adjusted funnel plot for the allelic model following adjustment with the trim-and-fill method. Solid circles denote the original studies, while hollow circles represent the imputed ones, respectively.
Discussion
4
Ischemic stroke (IS) is a complex pathological condition characterized by reduced blood flow and oxygen supply to brain tissue. The precise mechanisms linking NADPH oxidase to ischemic stroke remain unclear, although reactive oxygen species (ROS) have been implicated in its pathophysiology [10], 36]. NADPH oxidase is a key enzyme complex responsible for ROS production in the brain [36], 37]. Previous studies have suggested that superoxide-producing NADPH oxidases are associated with ischemia-reperfusion injury in the brain [38]. The C242T polymorphism in the p22phox subunit may disrupt the interaction between p22phox and NOX2 [16], potentially reducing ROS production [39], 40]. However, our meta-analysis did not identify a significant association between the p22phox C242T polymorphism and the risk of ischemic stroke, despite the presence of publication bias. This bias typically stems from the systematic under-reporting of small-scale studies with null or negative findings, consequently leading to an overestimation of the true effect size within meta-analyses. Therefore, the pooled ORs observed before adjustment should be interpreted as representing the probable upper bound of any potential association. Crucially, however, the robustness of our primary finding, a demonstrable lack of significant association, is robustly reinforced by both sensitivity analyses and the trim-and-fill method. The application of the trim-and-fill method, which imputes theoretically missing studies to correct funnel plot asymmetry, resulted in an adjusted effect size even closer to the null value (OR 1.027) that remained non-significant. This compellingly suggests that, even after accounting for potential publication bias, the conclusion of no association remains statistically robust.
Our null findings stand in stark contrast to earlier meta-analyses reporting a significant association between the C242T polymorphism and ischemic cerebrovascular disease [21], 22]. This discrepancy is largely ascribed to a critical methodological refinement in our study. Previous analyses incorporated primary studies where control groups significantly deviated from Hardy-Weinberg Equilibrium (HWE). Such deviations are a well-established indicator of potential genotyping errors, population stratification, or selection bias, which can significantly distort true genetic effect estimates in meta-analyses [41]. By rigorously excluding studies with HWE violations in controls, our analysis provides a more accurate and reliable pooled estimate. This methodological rigor aligns our results with the well-designed, null-finding study by Gu et al. [20] and highlights the critical importance of quality control in genetic association syntheses. Therefore, the principal contribution of this updated meta-analysis lies not merely in its recency, but in its application of a more stringent methodological standard, suggesting earlier positive associations may have been biased.
Furthermore, the relationship between the C242T polymorphism and ROS production is complex. Studies have shown that the T242 allele is associated with reduced NADPH oxidase activity and lower ROS levels in vascular cells [42], 43]. Arca et al. demonstrated that the T242 allele predicts a lower risk of recurrent cardiovascular events and is linked to reduced systemic oxidative stress [44]. In CC individuals, augmented NADPH oxidase activity was unrelated to elevated p22phox expression levels, implying that the C242T polymorphism disrupts the binding capacity of p22phox to gp91phox, thereby modulating NADPH oxidase function [42]. However, other research has reported no significant effect of the C242T polymorphism on oxidative stress levels in healthy individuals at rest [45]. These conflicting findings highlight the need for further research, particularly in larger and more diverse populations, to elucidate the role of this polymorphism in ischemic stroke.
The striking reduction in heterogeneity observed within specific subgroups, such as small-vessel occlusion (SVO) and Caucasian populations, warrants further consideration. Biologically, this may indicate a more homogeneous pathogenic role for NADPH oxidase-derived ROS in these distinct stroke etiologies or genetic backgrounds. For instance, the pathophysiology of SVO closely links to cerebral microangiopathy and endothelial dysfunction, processes where NADPH oxidase activity is profoundly implicated. The absence of significant heterogeneity in Caucasians could reflect a more uniform genetic architecture and a lower degree of population stratification compared to more diverse ethnic groups, thereby simplifying the genetic association and thus reinforcing this specific link.
Several limitations of our meta-analysis should be acknowledged. First, our findings rest primarily on unadjusted effect estimates. A notable limitation of this approach is its failure to adjust for potential confounding variables like age, hypertension, smoking status, diabetes, and lipid profiles. These factors are established independent predictors of IS and may also influence oxidative stress levels, thereby acting as confounders or effect modifiers in the genotype-phenotype relationship. The primary studies’ lack of adjustment for these variables introduces the risk that residual confounding could bias our pooled estimates. Consequently, the null association we observed might mask a subtle genuine association obscured by confounding, or conversely, bolster the evidence for a true null finding if such pervasive confounding existed yet still yielded no signal. Future meta-analyses incorporating individual-level data or adjusted estimates would be critical for providing more precise and confounder-resistant summary estimates. Second, our analysis is confined to published studies, which may introduce publication bias. The inclusion of unpublished studies with nonsignificant or negative results could alter the pooled estimate and provide a more balanced understanding of the true association. Third, the generalizability of our conclusions may be limited due to the ethnic diversity of the included studies. Only two studies focused exclusively on Caucasian populations, restricting statistical power and applicability to other ethnic groups. Fourth, while our meta-analysis found no statistically significant correlation, it is possible that a small true effect exists below the detection threshold of the available sample size, or that complex gene-environment interactions obscured a true relationship. Finally, the significant statistical heterogeneity across studies may affect the precision and robustness of our pooled estimate.
In summary, our meta-analysis suggests that the NADPH oxidase p22phox C242T polymorphism does not confer a significant risk for ischemic stroke. However, further large-scale studies with diverse populations are needed to validate these findings. Additionally, exploring potential interactions between the p22phox C242T polymorphism and other genetic or environmental factors may provide deeper insights into its role in ischemic stroke susceptibility.
Supplementary Material
Supplementary Material
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Jiang M Yin P Bai X Yang L Zhang J Xu S Proinflammatory and anti-inflammatory genes in stroke pathogenesis Curr Pharm Des 20202642203310.2174/138161282666620070121285932611297 · doi ↗ · pubmed ↗
- 2Deng XD Ke JL Chen TY Gao Q Zhao ZL Zhang W ERCC 1 polymorphism and its expression associated with ischemic stroke in Chinese population Front Neurol 20231399842810.3389/fneur.2022.99842836712419 PMC 9878395 · doi ↗ · pubmed ↗
- 3Wang M Shen Y Gao Y Chen H Duan F Li S NQO 1 polymorphism and susceptibility to ischemic stroke in a Chinese population BMC Med Genomics 20241721910.1186/s 12920-024-01992-739174970 PMC 11342592 · doi ↗ · pubmed ↗
- 4Wang P Lu J Wang M Wang G Chen H NRF 2 polymorphism and susceptibility to ischemic stroke in a Chinese population Peer J 202513 e 1974210.7717/peerj.1974240718775 PMC 12296571 · doi ↗ · pubmed ↗
- 5Sarmah D Datta A Raut S Sarkar A Shah B Bohra M The role of inflammasomes in atherosclerosis and stroke pathogenesis Curr Pharm Des 20202642344510.2174/138161282666620042708494932338208 · doi ↗ · pubmed ↗
- 6Batty M Bennett MR Yu E The role of oxidative stress in atherosclerosis Cells 202211384310.3390/cells 1123384336497101 PMC 9735601 · doi ↗ · pubmed ↗
- 7Koutsaliaris IK Moschonas IC Pechlivani LM Tsouka AN Tselepis AD Inflammation, oxidative stress, vascular aging and atherosclerotic ischemic stroke Curr Med Chem 202229549650910.2174/092986732866621092116171134547993 · doi ↗ · pubmed ↗
- 8Yan R Zhang X Xu W Li J Sun Y Cui S ROS-induced endothelial dysfunction in the pathogenesis of atherosclerosis Aging Dis 2024162506810.14336/AD.2024.030938502586 PMC 11745424 · doi ↗ · pubmed ↗