Crystal structure of the Legionella pneumophila effector SidL (Lpg0437) in complex with its metaeffector LegA11 (Lpg0436)
Dominik A. Machtens, Carissa A. Hutchison, Ashley M. Stein, Jan Eberhage, Jonas M. Willerding, Susanne Eschenburg, Stephanie R. Shames, Thomas F. Reubold

TL;DR
This study reveals how the protein LegA11 suppresses the activity of SidL in Legionella pneumophila, helping the bacteria avoid host defenses.
Contribution
The crystal structure of the SidL-LegA11 complex and functional validation of their interaction is newly presented.
Findings
The SidL-LegA11 complex forms a high-affinity 1:1 interaction with a 2.4 Å crystal structure.
LegA11 suppresses SidL's ability to inhibit host mRNA translation through direct binding.
Mutagenesis of key residues in LegA11 disrupts its metaeffector function.
Abstract
Legionella pneumophila is an opportunistic human pathogen that causes atypical pneumonia called Legionnaires’ Disease. To replicate within host cells, L. pneumophila injects up to 330 effector proteins into the host cytosol via a Dot/Icm type IV secretion system. Several effectors, called metaeffectors, regulate the activity of other effectors within infected host cells through direct protein-protein interactions. LegA11 (AnkJ/Lpg0436) has been identified as a metaeffector of SidL (Ceg14/Lpg0437), one of eight L. pneumophila effectors that inhibit host mRNA translation. LegA11 binds and suppresses SidL enzymatic activity, but the molecular basis of this regulation and impact on mRNA translation are unknown. Here, we present the crystal structure of SidL in complex with LegA11 to a resolution of 2.4 Å, revealing a high-affinity 1:1 complex with an extensive interaction interface of ~…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
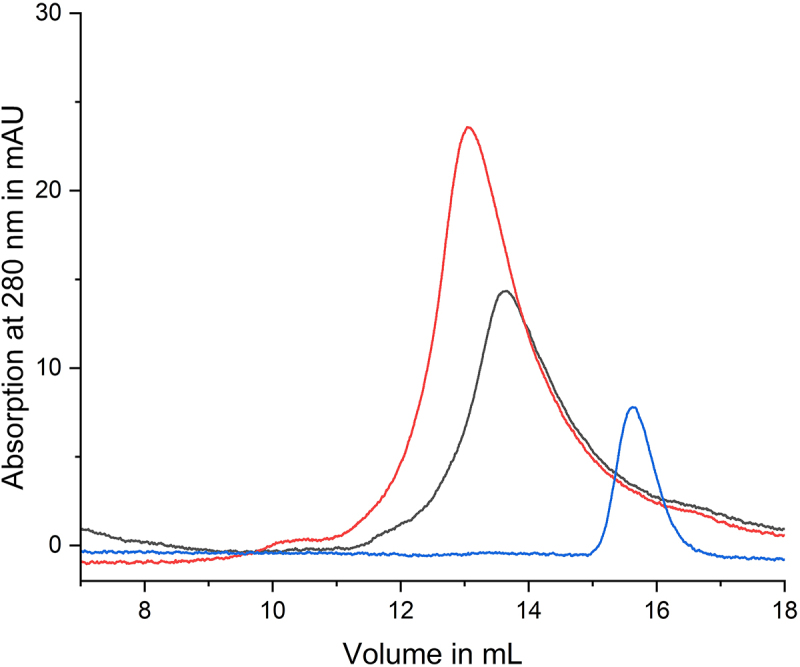 Figure 1
Figure 1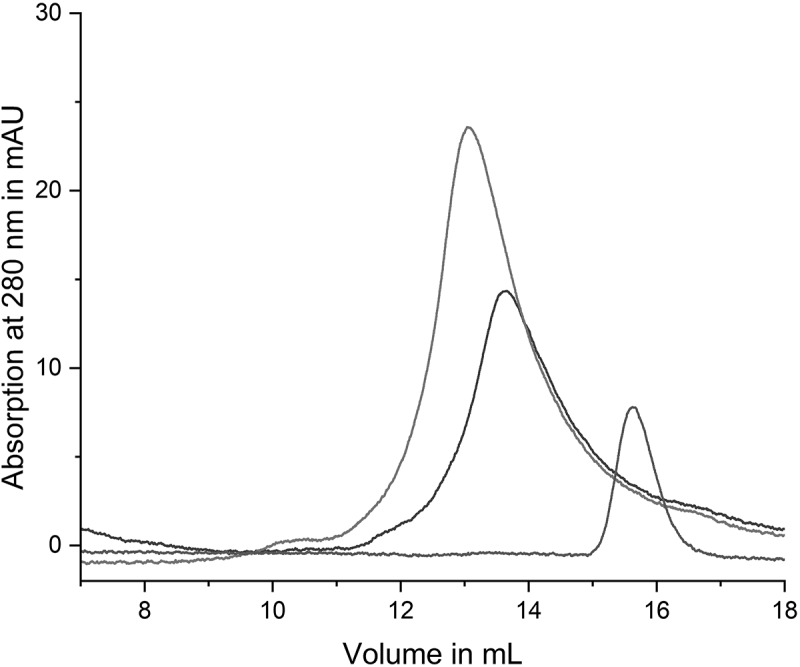 Figure 2
Figure 2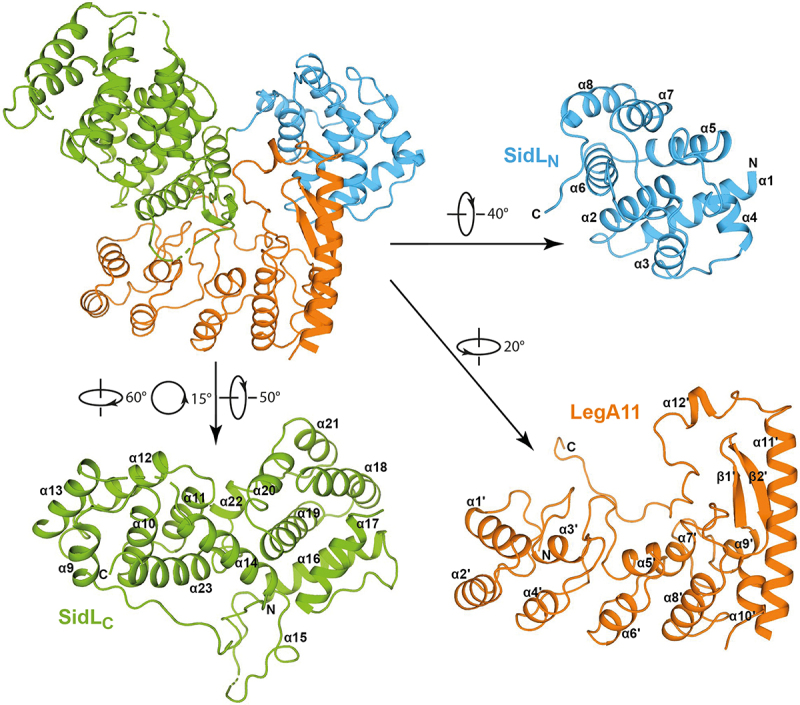 Figure 3
Figure 3 Figure 4
Figure 4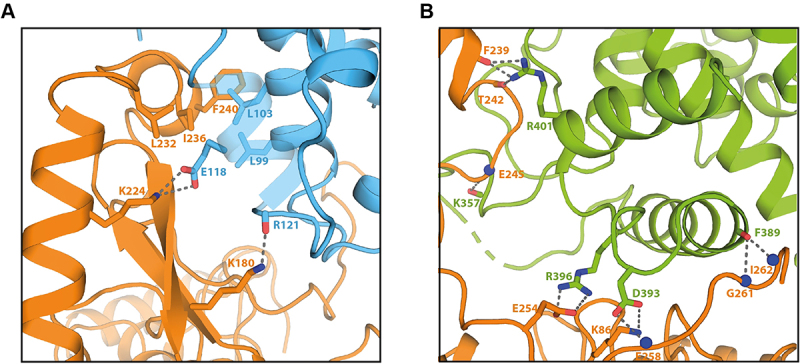 Figure 5
Figure 5 Figure 6
Figure 6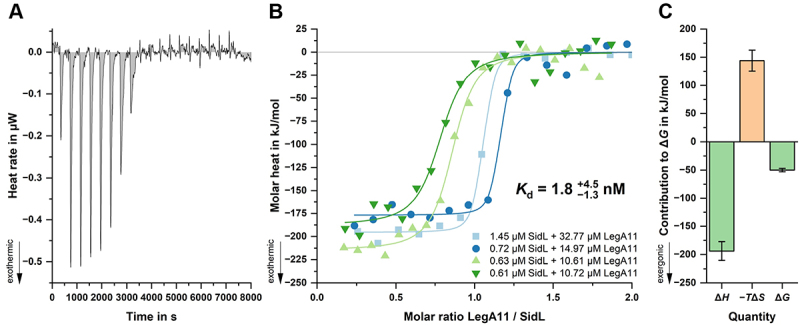 Figure 7
Figure 7 Figure 8
Figure 8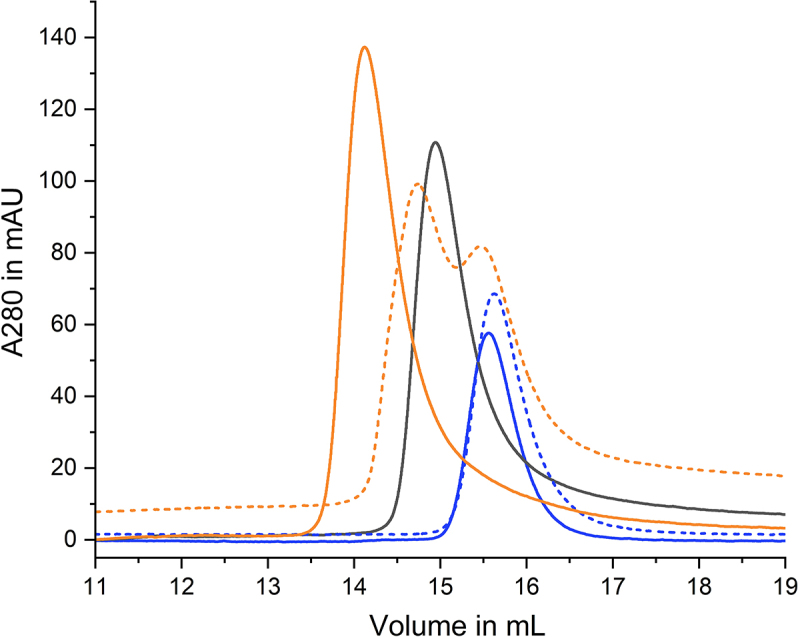 Figure 9
Figure 9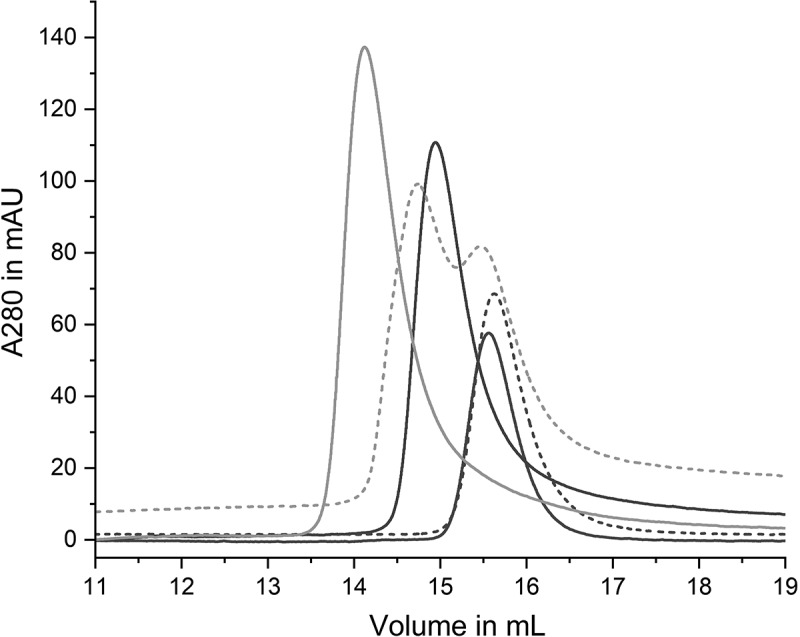 Figure 10
Figure 10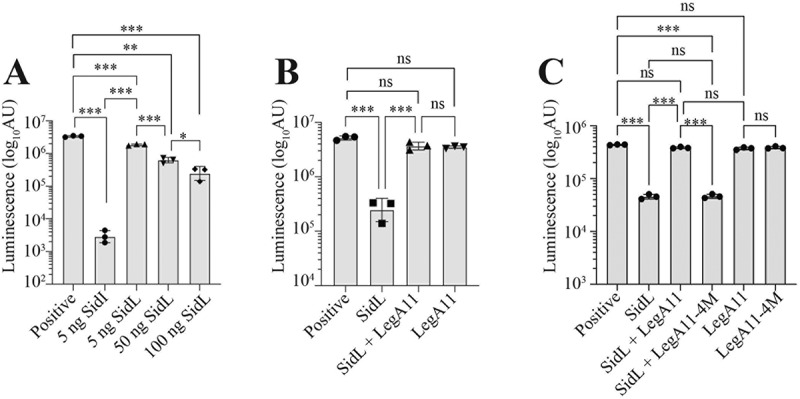 Figure 11
Figure 11- —National Institutes of Health10.13039/100000002
- —Kansas State University10.13039/100007765
- —Michigan State University10.13039/100007709
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsLegionella and Acanthamoeba research · Escherichia coli research studies · Tuberculosis Research and Epidemiology
Introduction
Legionella pneumophila is an opportunistic pathogen that is ubiquitous in freshwater environments where it parasitizes free-living amoebae [1]. L. pneumophila can also replicate within alveolar macrophages, leading to a potentially life-threatening atypical pneumonia called Legionnaires’ Disease. To replicate within host cells, L. pneumophila remodels the phagosome into a replication-permissive endoplasmic reticulum-like compartment called the Legionella-containing vacuole (LCV). To achieve this, L. pneumophila injects up to 330 effector proteins into the cytosol of the host cell through a Dot/Icm type IV secretion system (T4SS) [2]. L. pneumophila effectors interfere with and subvert key host cellular processes including ubiquitin-dependent proteasomal degradation, cell death, phagocytosis, vesicular trafficking, autophagy, and mRNA translation [3].
Several cellular processes are targeted by multiple effectors, reflecting L. pneumophila’s co-evolution with phylogenetically diverse hosts [4]. So far, eight different effectors have been identified in L. pneumophila that inhibit host mRNA translation. Lgt1 (Lpg1368), Lgt2 (Lpg2862), and Lgt3 (Lpg1488) are glycosyl transferases that transfer a glucose moiety to a specific serine residue in the eukaryotic translation elongation factor 1A [5,6]. The serine/threonine kinase LegK4 (Lpg0208) blocks translation by phosphorylation of a threonine residue in Hsp70 proteins [7], whereas VipF (Lpg0103) is an acetyl transferase that targets eukaryotic initiation factor 3 complex [8]. SidI (Lpg2504) transfers a mannosyl group to several ribosomal proteins [9]. The mechanism of action of RavX (Lpg1489) is currently unknown. SidL (Ceg14/Lpg0437) blocks translation via an unknown mechanism but is known to bind actin and has recently been shown to function as an ATPase that hydrolyzes ATP to AMP and pyrophosphate in the presence of host actin [10,11]. The dedication of multiple effectors to subversion of host translation machinery underscores its importance for L. pneumophila virulence.
Work over the past 15 years has shown that a subset of L. pneumophila effectors, termed metaeffectors, do not exclusively target host proteins but instead serve to regulate the function of other effectors through a direct protein-protein interaction [12,13], distinguishing them from antagonistic effectors that exert opposing biochemical activities on the same host factor(s) [13]. Both SidI and SidL are regulated by metaeffectors, suggesting a role for this regulatory mechanism in L. pneumophila’s subversion of host mRNA translation. SidI is regulated by MesI (Metaeffector of SidI), which forms a high-affinity 1:1 complex with SidI and fully suppresses SidI-mediated translation inhibition and toxicity [14,15]. The effector LegA11 (AnkJ/Lpg0437) binds SidL and alleviates the toxic effect of SidL in yeast cells [11,16], but the molecular and biophysical underpinnings of the SidL-LegA11 interaction and impact of LegA11 on SidL-mediated translation inhibition are unknown.
Here, we present the crystal structure of SidL in complex with its cognate metaeffector LegA11 to a resolution of 2.4 Å. Using an in vitro translation assay, we show that SidL inhibits mRNA translation by an order of magnitude and that this activity is suppressed in the presence of LegA11. Based on the crystal structure, we identified residues in LegA11 that upon mutation disrupt the binding interface and abolish the metaeffector activity of LegA11. Thus, LegA11 is a metaeffector that contributes to L. pneumophila’s subversion of host mRNA translation via a direct protein-protein interaction with its cognate effector, SidL [17].
Material and Methods
Bacterial strains, cell culture, and growth conditions
Escherichia coli strains used for cloning (Top10Invitrogen; DH5αλpir; a gift from Dr. Craig Roy, Yale University) and protein expression [BL21 (DE3); Invitrogen] were grown at 37°C in lysogeny broth (LB) medium supplemented with antibiotics for plasmid selection [100 µg/mL ampicillin(GoldBio, A-301), 50 µg/mL kanamycin (GoldBio K-120), 25 µg/mL chloramphenicol (GoldBio, C-105)]. L. pneumophilaSRS43 (Lp02 thyA+;a gift from Dr. Craig Roy, Yale University [18]) strains were cultured on charcoal N (2-acetamido)-2-aminoethanesulfonic acid (ACES)-buffered yeast extract [10 g/L ACES (Sigma, A9758), 10 g/L yeast extract (BD, 212,750), 2 g/L activated charcoal (Sigma, C5510), pH 6.9] freshly supplemented with filter sterilized L-cysteine (0.4 g/L; Sigma, C7352) and Fe(NO_3_)3 (0.135 g/L; Sigma, 216,828) at 37°C [19]. Liquid ACES-buffered yeast extract (AYE) was prepared as described without charcoal and agar. When necessary, media were supplemented with 10 µg/mL chloramphenicolfor plasmid selection. Plasmid-based gene expression in L. pneumophila was induced with 1 mM of filter sterilized isopropyl β-D-1-thiogalactopyranoside (IPTG; GoldBio, I2481C).
Acanthamoeba castellanii strain Neff was acquired from the American Type Culture Collection (ATCC #30010) and maintained in PYG broth [7.5 g/L proteose peptone (BD, 211,693), 7.5 g/L yeast extract (BD, 212,750, 15 g/LD(+)-glucose (Thermo, A1682836), 0.4 mM CaCl_2_ (Sigma, C4901), 4 mM MgSO_4_ (Sigma, M7506), 3.4 mM Na citrate (Sigma, W302600), 50 µM F_3_(NH_4_)2(SO_4_)2, (Sigma, 215,406), 2.5 mM Na_2_HPO_4_ (Sigma, S9763), 2.5 mM KH_2_PO_4_ (Sigma, P5655), pH 6.5] at 25°C [20].
Molecular cloning and strain construction
Oligonucleotide primersequences are listed in Suppl. Table S1. For expression of His_6_-Myc fusion proteins, the respective L. pneumophila genes were cloned into pT7HMT, a gift from Dr. Brian Geisbrecht [21]. Wild-type sidL and legA11 were amplified from L. pneumophila SRS43 genomic DNA (gDNA) using SidLBamHI-F/SidLNotI-R and LegA11BamHI-F/LegA11NotI-R primer pairs. legA11K195A/F239R/G240R/F243R (legA11-4 M) was amplified from cDNA on a high copy cloning vector purchased from Twist Biosciences (San Francisco, CA) using LegA11BamHI-F/LegA11NotI-R. BamHI/NotI-digested fragments were purified and ligated into BamHI/NotI-digested pT7HMT [21]. Sequence-confirmed constructs (Genewiz) were transformed into chemically competent E. coli Top10 and BL21 (DE3) for maintenance and protein expression, respectively. For genetic complementation of ∆sidL and ∆legA11 mutations, sidL and legA11 were amplified from L. pneumophila SRS43 gDNA using SidLJBBamHI-F/SidLSalI-R and LegA11JBBamHI-F/LegA11SalI-R primer pairs and ligated as BamHI/SalI fragments into BamHI/SalI-digested pSN85, a gift from Dr. Craig Roy [22]. Sequence confirmed pSN85:legA11 and pSN85:sidL plasmids were transformed into electrocompetent L. pneumophila ∆legA11 and ∆sidL using a BioRad Gene Pulser (2400 V, 25 µF, 2 mm) [14].
L. pneumophila SRS43 ∆legA11 and ∆sidL were generated by allelic exchange [18]. Deletion constructs were generated by cloning 5” and 3” flanking regions of each gene into pSR47S. To generate pSR47S:∆legA11, 5” and 3” flanking regions were amplified using LegA11KO1-F/LegA11KO1-R and LegA11KO2-F/LegA11KO2-R primer pairs, digested with SacI/NotI and NotI/SalI, respectively, and ligated into SacI/SalI-digested pSR47S (a gift from Dr. Craig Roy). To generate pSR47S:∆sidL, 5“ and 3” flanking regions were amplified using SidLKO1-F/SidLKO1-R and SidLKO2-F/SidLKO2-R primer pairs, digested with SacI/NotI and NotI/SalI, respectively, and ligated into SacI/SalI-digested pSR47S. Ligation reactions were transformed into chemically competent E. coli DH5αλpir. Deletion constructs were validated by colony PCR and Sanger Sequencing (Genewiz) and conjugated into L. pneumophila SRS43 followed by selection of sucrose-resistant, kanamycin-sensitive colonies were screened by PCR and Sanger sequencing to verify gene deletion [23].
Protein expression and purification
The coding sequences of sidL (lpg0437) (UniProt accession number Q5ZYD5) in full-length (SidL_1s_BamHI/SidL_666as_SalI) or in N- and C-terminally truncated form (SidL_23s_BamHI/SdiL_640as_SalI; SidL_76s_BamH/SidL_645as_SalI) and of legA11(lpg0436) in full-length (LegA11_1s_BamHI/LegA11_269as_SalI) were amplified from Legionella pneumophila strain Philadelphia 1 (DSM-7513, DSMZ, Braunschweig, Germany) gDNA using primer pairs and cloned into either pGEX-4T1-TEV or pMEX-4T1-TEV [24]. Point mutations were introduced by overlap-PCR. The correctness of the resulting constructs was verified by sequencing (Seqlab, Göttingen, Germany). The constructs were expressed in Escherichia coli BL21 (DE3) cells grown to an OD of 1.0 in terrific broth. Expression was induced with 0.2 mM IPTG (Thermo Scientific, R0392) overnight at 293 K. The bacteria were harvested by centrifugation for 15 min at 5000 × g. Pellets were resuspended in 50 mM HEPES-NaOH (Roth, HN78, T135) pH 7.5, 200 mM NaCl (Th. Geyer, 1367), 5 mM 2-mercaptoethanol (β-ME) (SERVA 28,625), 1 mM phenylmethyl sulfonyl fluoride (PMSF) (Roth, 6367), 0.5% (v/v) Nonidet^TM^ P-40 substitute (Sigma-Aldrich, 74,385), and 0.3 mg/mL hen egg-white lysozyme (Applichem, A3711) and incubated on ice for 30 min. The cells were lysed by sonication and cellular debris was removed by centrifugation at 32,000 × g for 30 min. The supernatant was loaded onto either an amylose sepharose (NEB, E8201) or a glutathione Superflow agarose (Thermo Scientific, 25,237) column, depending on the respective purification tag, equilibrated with a buffer containing 50 mM HEPES pH 7.5, 300 mM NaCl, and 5 mM 2-mercaptoethanol. After washing with 10 column volumes of buffer the bound fusion protein was digested on column with tobacco etch virus (TEV) protease [25] overnight. The cleaved protein was eluted with column buffer, concentrated to a volume of 2 mL, and applied to a S200 16/600 size exclusion column (GE Healthcare) equilibrated with a buffer containing 20 mM HEPES-NaOH pH 7.5, 150 mM NaCl, and 2 mM DTT (Applichem, A2948). The peak fractions were pooled, concentrated and flash-frozen in liquid nitrogen. For preparation of seleno-methionine (SeMet) (Calbiochem, 561,505) substituted SidL and LegA11, proteins were expressed in E. coli BL21(DE3) following a protocol using PSM-5052 auto-induction medium [26].
Analytical size exclusion chromatography
Equimolar amounts of SidL and/or LegA11 in a total volume of 100 µL were applied to an S200 10/300 size exclusion column (GE Healthcare) equilibrated with buffer containing 20 mM HEPES pH 7.5, 150 mM NaCl, and 2 mM DTT at a flow rate of 0.5 ml/min. The concentrations used are given in the respective figure legends. The figures representing the chromatographic data were prepared using Origin (Version 2022b. OriginLab Corporation, Northampton, MA, USA).
Crystallization
Crystallization screening using various commercially available screens was performed in the 96-well format by mixing 200 nL of protein solution containing 12 mg/mL SidL_23–640_, 5.2 mg/mL LegA11, and chymotrypsin, trypsin, or thermolysin (Sigma-Aldrich, C3142, T5266, P1512) in ratios between 1:500 to 1:2000 with 200 nL of reservoir solution in MRC plates using a Phoenix crystallization robot (Art Robbins Instruments, Sunnyvale, USA). The droplets were equilibrated against 50 µL reservoir solution at 291 K. Plate-shaped crystals grew in condition F1 (100 mM HEPES-NaOH pH 7.5 (Sigma-Aldrich, H3375, S7653), 200 mM L-proline (Sigma-Aldrich, 81,709), 100 g/L PEG3350 (Sigma-Aldrich, P4338)) of the Classics 2 screen (Qiagen, Hilden, Germany) in the presence of trypsin at 17 µg/mL. SeMet-derivatized crystals used for cryogenic data collection were grown in droplets of 1 µL protein (12 mg/mL SidL_23–640_, 5.2 mg/mL LegA11, trypsin at 34 µg/mL) and 1 µL reservoir containing 100 mM HEPES-NaOH pH 7.5, 200 mM L-proline, and 200 g/L PEG3350 equilibrated against 200 µl reservoir solution. Crystals were cryoprotected by brief immersion in reservoir solution containing 140 g/L PEG3350 and 23% (v/v) ethylene glycol (Sigma-Aldrich, 324,558) and flash-cooled in liquid nitrogen. Native crystals used for cryogenic data collection were grown in droplets of 1 µL protein (12 mg/mL SidL_23–640_, 5.2 mg/mL LegA11, trypsin at 340 µg/mL) and 1 µL reservoir containing 100 mM HEPES-NaOH pH 7.5, 200 mM L-proline, and 140 g/L PEG3350 equilibrated against 200 µl reservoir solution. Crystals were cryoprotected by serial immersion in reservoir solutions containing 140 g/L PEG3350 and 0, 2, 4, 7, 10, 13, 16, 19, or 23% (v/v) glycerol (Sigma-Aldrich, 49,767) and flash-cooled in liquid nitrogen.
Data collection and processing
A dataset from a single native crystal was collected to a resolution of 2.35 Å at beamline PROXIMA 2A at the synchrotron SOLEIL (Gif-sur-Yvette Cedex, France) using a wavelength of 0.9762 Å. Four datasets were collected from a crystal grown from SeMet-substituted protein at a wavelength of 0.9795 Å at beamline P13 at the PETRA III storage ring (Hamburg, Germany) and merged to yield a complete highly redundant dataset to 3.3 Å. Data were processed with XDS and scaled with XSCALE [27,28]. For both crystals space group P2_1_2_1_2_1_ could be assigned with unit cell dimensions of a = 68.5 Å, b = 93.4 Å, and c = 133.7 Å for the native crystal and of a = 657 Å, b = 974 Å, and c = 1307 Å for the SeMet-crystal, respectively
Structure solution and refinement
The structure was solved by the single anomalous dispersion (SAD) method employing the CRANK2 pipeline [29]. The initial electron density map was of sufficient quality for automatic building of 614 residues. The structure was iteratively completed and refined against the native data using Phenix.refine [30] followed by visual inspection and adjustment of the model in COOT [31] after each refinement cycle. Images showing structural data were prepared using Pymol (The PyMOL Molecular Graphics System, Version 1.8.6.2, Schrödinger, LLC). The data statistics are summarized in Table 1. The atomic coordinates and structure factors of SidL/LegA11 have been deposited in the Protein Data Bank (PDB entry 9SXC).Table 1.Data collection and refinement statistics. SidL/LegA11 nativeSidL/LegA11 SeMetData collection Wavelength (Å)0.9785650.97950Space groupP 21 21 21P 21 21 21Unit cell a, b, c (Å)68.53, 93.38, 133.6769.65, 97.38, 130.30Resolution range (Å)47.85 - 2.4 (2.486 - 2.4)^a^78.003 – 3.3 (3.4 – 3.3)CC_1/2_0.999 (0.665)0.999 (0.892)R_meas_^b^0.1425 (1.411)0.304 (2.02)Total reflections288,234 (37736)696,282 (62675)Unique reflections41,606 (5384)25,754 (2217)Completeness (%)99.70 (100.00)100.00 (100.00)Redundancy6.9 (7.0)27.0 (28.3)<I/σ(I)>23.42 (2.04)16.13 (3.87)Structure refinement Resolution range (Å)47.85 - 2.4 (2.47 – 2.40) R_work_^c^0.2244 (0.2995) R_free_^d^0.2672 (0.3812) No. of atoms5658 No. of protein atoms5472 No. of water molecules186 B-factors protein/water66.71/59.32 R.m.s.d. Bond lengths (Å)0.003 R.m.s.d. Bond angles (°)0.56^e^^d^ Protein Data Bank entry### ^a^The values in parentheses refer to statistics in the highest bin.^b^R_meas_ = Σ_hkl_ (n_hkl_/n_hkl_ − 1)^1/2^ Σ_i_|<I_hkl_ > −I_hkl,i_|/Σ_hkl_ Σ_i_ I_hkl,i_^c^R_work_ = Σ_hkl_|F_o_(hkl)-F_c_(hkl)|/Σ_hkl_F_o_(hkl), where F_o_ and F_c_ are the observed and calculated structure-factor amplitudes, respectively.^d^R_free_ was calculated with 5% of the data excluded from the refinement.^e^R.m.s.d. = root mean square deviation.
Isothermal titration calorimetry (ITC)
Binding between SidL (residues 76–645) and LegA11 (residues 1–269) was measured using a NanoITC 5302 microcalorimeter (Waters/TA Instruments) at 25°C. Proteins were prepared in 20 mM HEPES-NaOH (pH 7.5), 150 mM NaCl, degassed for 15 min at 30 mbar, and centrifuged at 20,000 × g for 5 min before titration. The sample cell (971 µL) contained SidL at 0.6 to 1.5 µM, with concentrations determined photometrically at 280 nm for each experiment. The syringe (100 µL) was loaded with 10 to 30 µM LegA11, also quantified by UV absorbance. Each experiment consisted of 20 injections of 4.91 µL ligand, delivered at 200 rpm stirring speed with 400 s spacing. Buffer-into-protein control titrations were performed, and the resulting heats were subtracted from binding data. Thermograms were integrated using the NanoITC software, and binding isotherms were fitted in Origin (OriginLab) with an independent binding model. Parameters obtained directly from the fit included the stoichiometry (n), dissociation constant (Kd), and enthalpy change (ΔH); Gibbs free energy (ΔG), entropy (ΔS), and −TΔS were calculated subsequently. Errors were estimated by error propagation, with asymmetric standard deviations reported for Kd. Data are representative of four independent experiments.
In vitro translation assay
Recombinant His_6_-Myc-SidL and -LegA11 were expressed in E. coli BL21 (DE3) [14]. Overnight cultures were subcultured into LB (1%) and grown for 3 h at 37°C with shaking. Cultures were supplemented with 1 mM IPTG, grown overnight at 16°C, pelleted by centrifugation at 2100 × g for 10 min at 4°C, and washed in 5 mL of ice-cold PBS. Bacterial pellets were resuspended in ice-cold lysis buffer [His Binding buffer (Zymo His-Spin Protein Miniprep Kit),10 µg/mL lysozyme (Sigma, L6876), and 10 mM 2-mercaptoethanol (Sigma, M6250)] and incubated on ice for 2–4 h followed by sonication using Kontes Micro Ultrasonic Cell Disrupter (40 amplitude, 30 s, five times). Lysates were clarified via centrifugation at 9600 × g for 15 min at 4°C. Supernatants were transferred to clean microcentrifuge tubes and His-tagged proteins were then purified using the Zymo His-Spin Miniprep kit via manufacturer’s instructions (Zymo Research). Purity was validated by SDS-PAGE and Coomassie Blue staining. Purified recombinant proteins were dialyzed overnight in PBS pH 7.4 at 4°C and quantified using the Coomassie Plus Bradford Assay (ThermoFisher, 23,238) reagent on a BioTek Epoch2 microplate reader (Agilent).
Translation of Firefly Luciferase mRNA in Rabbit Reticulocyte Lysates was quantified using the Flexi Rabbit Reticulocyte Kit (Promega, Madison, WI) [14]. Purified recombinant His_6_-Myc-SidL (50 ng) and/or His_6_-Myc-LegA11 (23.3 ng) were added to lysates in equimolar amounts (65 nM). Reactions were run in triplicate for 90 min at 30°C followed by addition of Luciferase assay reagent (Promega, E1483) and luminescence (arbitrary units) was quantified using a BioTek Synergy HTX multimode plate reader (Agilent).
L. pneumophila growth curves
L. pneumophila was cultured on CYE agar at 37°C and single colonies were used to generate 2-day heavy patches. One day prior to infection, A. castellanii cultures were split 1:5 into 75 cm^2^ tissue culture flasks and incubated in PYG media at 25°C. Cells were seeded in 24-well tissue culture-treated dishes (2.5 ×10^5^ cells/well) in 1 mL PYG and incubated at 37°C for 2 h. Adhered cells were washed in pre-warmed Ac buffer (0.4 mM CaCl_2_, 4 mM MgSO_4_, 3.4 mM Na citrate, 0.05 mM F_3_(NH_4_)2(SO_4_)2, 2.5 mM Na_2_HPO_4_, 2.5 mM KH_2_PO_4_, pH 6.5) and infected with heavy-patch grown L. pneumophila [multiplicity of infection (MOI) of 0.1] in 1 mL Ac buffer in triplicate wells. Cells were incubated for 45 min and monolayers were washed three times with PBS to remove extracellular bacteria. Cells were either lysed (bacterial uptake) or incubated in 1 mL Ac buffer for up to 72 h at 37°C. Cells were lysed hypotonically in sterile deionized water and lysates were plated on CYE agar for enumeration of colony forming units (CFU) at 45 min, 24 h, 48 h, and 72 h post-infection [18].
To quantify L. pneumophila growth in vitro, 2-day heavy patches grown on CYE agar were resuspended in supplemented AYE broth and subcultured to an OD_600_ of 0.2. Cultures were dispensed into sterile 96-well round bottom plates (n = 6/strain) and incubated with continuous orbital shaking in a BioTek Epoch2 microplate reader. Data were collected every 2 h for up to 46 h.
Statistical analyses
Data are shown as mean ± standard deviation (s.d.) or standard error (s.e.m), as indicated. Statistical analyses were performed with GraphPad Prism 10 using either Welch’s t-test, one-way ANOVA, or two-way ANOVA, as described, with statistical significance defined as p < 0.05. Unless otherwise indicated, statistical analyses were performed on biological triplicate samples (N = 3) and representative of three independent experiments.
Results
SidL directly interacts with LegA11 forming a stable 1:1 complex
LegA11 was initially identified as a metaeffector based on its ability to suppress the toxic effect of SidL in a heterologous yeast expression system and bind to SidL in yeast two-hybrid and LUMIER assays [16]. A direct protein-protein interaction between purified recombinant SidL and LegA11 was recently validated [11], but the kinetics and biophysical properties of the SidL-LegA11 interaction are unknown. To verify the direct interaction of SidL and LegA11 and assess the stoichiometry of the SidL-LegA11 complex, we subjected purified recombinant full-length SidL_1–666_, LegA11, or equimolar amounts of SidL_1–666_ and LegA11 to analytical size exclusion chromatography (SEC). In the chromatogram corresponding to the mixture, the peaks of the single proteins have merged into a single peak shifted to a lower elution volume (Figure 1). This indicates that SidL and LegA11 form a stable complex with 1:1 stoichiometry. Figure 1.Interaction of SidL and LegA11 observed in analytical size exclusion chromatography. 300 µg SidL_1–666_ and/or 125 µg LegA11 in 100 µL buffer were applied to an S200 10/300 increase size exclusion column. The absorption curve of SidL_1–666_ is shown in black, the absorption curve of LegA11 is shown in blue, the absorption curve of the mixed proteins is shown in red.A line graph showing absorption at 280 nanometers in milliabsorbance units versus volume in milliliters for protein interactions.
We employed X-ray crystallography to visualize the SidL/LegA11 complex. Full-length SidL could not be purified in amounts large enough for an extended crystallization campaign; thus, we used the truncated variant SidL_23–640_ for overexpression as it could be produced in higher yield and displayed higher stability than the full-length protein. We set up crystallization trials using various commercial screens with an equimolar mixture of SidL and LegA11 but did not obtain any crystals. We then used in situ proteolysis [32] by adding various proteases to the crystallization setups (see Materials and Methods). We obtained crystals in several conditions, the best of which diffracted X-rays to a resolution limit of 2.4 Å after optimization. We solved the structure by SAD using data from a crystal composed of selenomethionine-substituted protein. Crystallographic details are given in Table 1.
The crystal structure contains residues 76–313, 334–413, and 525–628 of SidL and residues 7–192, and 202–261 of LegA11, as well as 186 water molecules. SidL is divided into two subdomains comprising residues 76–222 (SidL_N_) and residues 228–628 (SidL_C_), respectively (Figure 2). SidL_N_ is connected to SidL_C_ via a straight elongated stretch of five residues (223–227). The two subdomains are separated by a C-terminal portion of LegA11 and do not interact with each other directly. Presumably, in the absence of LegA11 the domains display a high degree of conformational flexibility. Figure 2.Overall structure of SidL/LegA11. Cartoon representation of the three-dimensional structure of the complex formed by SidL and LegA11. The model in the upper left corner represents the complex. The two subdomains of SidL are shown in blue and green, LegA11 is shown in orange. For better visibility, the complex components have been rotated individually, the respective degrees of rotation relative to the orientation of the complex are shown next to the arrows. Secondary structure elements as well as the n- and C-termini of the individual components are labeled.The image shows the three-dimensional structure of the SidL/LegA11 complex. The complex is divided into three main components: SidLN, SidLC, and LegA11. SidLN and SidLC subdomains are depicted in blue and green, respectively, and LegA11 in orange. The SidLN subdomain includes alpha helices labeled from alpha 1 to alpha 8, with the N-terminus and C-terminus indicated. SidLC features alpha helices labeled from alpha 9 to alpha 23, with the N-terminus and C-terminus marked. LegA11 contains alpha helices labeled from alpha 1 prime to alpha 12 prime and beta sheets labeled beta 1 prime and beta 2 prime, with the N-terminus and C-terminus shown. The image includes rotation angles for better visibility: SidLN is rotated 40 degrees, SidLC is rotated 60 degrees, 15 degrees and 50 degrees and LegA11 is rotated 20 degrees. The complex components are individually rotated to enhance visibility, with arrows indicating the direction of rotation.3D structure of SidL/LegA11 complex with subdomains SidLN, SidLC and LegA11.
SidL_N_ consists of eight rather short helices (α1-α8) that form pairs, which are aligned in a nearly parallel fashion except for the second pair, where α4 is tilted by about 50° (Figure 2). The order of the helix pairs is α3/α4, α1/α2, α5/α6, and α7/α8. Helices within pairs are connected via short loops, whereas the pairs are interconnected by longer loops. SidL_C_ displays an elongated shape that is roughly divided into two parts. The first part contains helices α9-α13 that are arranged in parallel layers of three (α9-α11) and two (α12 and α13). The helix axes are oriented roughly parallel to each other. This part is completed by the C-terminal helix α23 that packs against the three-helix layer in an orthogonal fashion. The second part is composed of the helix pair α16 and α17 that packs orthogonally against a helix bundle formed by helices α18-α21. Both parts are connected via helix α14 that interacts with α23 and α19 and the short helix α22 preceding the terminal helix α23. α14 and α16 are connected via a long loop of 20 residues that contain the single-turn helix α15. Between α17 and α18, about 110 residues are missing that presumably have been removed by the protease used for crystallization.
The N-terminal part of LegA11 is formed by four ankyrin repeats, of which the first three display the typical arrangement of short, paired helices (α1’/α2,’ α3’/α4,’ α5’/α6’) with the connecting loops between pairs protruding at an angle of approximately 90° [33]. The fourth repeat (α7’/α8’) deviates from the other three through longer helices and a shortened loop between the third and fourth repeat. The first three repeats correspond to a previously published crystal structure (PDB ID 4ZHB) comprising residues 11–114. Both structures can be superimposed with a root mean square deviation of 0.52 Å. The fourth repeat is immediately followed by a pair of very short helices, α9’ and α10.’ These helices are oriented roughly perpendicular to each other and are connected via a twisted two-stranded anti-parallel β-sheet formed by strands β1’ and β2.’ The β-sheet is “wrapped” by the long helix α11’ following helix α10’ on one side, the single-turn helix α12’ at the top, and an extended loop on the other side. The overall shape resembles the letter L with the ankyrin repeats forming the stalk and the C-terminal extension forming the base.
A search for structural homologs using the DALI server with SidL_N_ as search model did not yield significantly similar structures. An equivalent search using SidL_C_ as search model identified the recently published cryo-EM structure of the Legionella effector LnaB (PDB ID 8JO4) as the closest structural homolog. Superposition of SidL_C_ with the N-terminal domain of LnaB yielded an r.m.s.d. value of 2.7 Å for 201 equivalent Cα atoms (Suppl. Fig. S1). The identity of the aligned sequence stretches is 21.9%.
Characterization of the SidL/LegA11 binding interface
SidL interacts with LegA11 via an interface of about 2300 Å^2^ as determined by PISA and sppIDER [34,35]. SidL_C_ contacts LegA11 via the inside of both the stalk and the base of the L-shape with an interaction surface of about 1500 Å^2^, whereas SidL_N_ is placed on the opposite side of the base of LegA11 (Figure 2). The interaction of SidL_N_ and LegA11 is mediated via several polar side chain-main chain interactions, for instance between the side chain of K180 of LegA11 with the main chain oxygen of R121 of SidL or the side chain of E118 of SidL with the main chain nitrogen of I178 of LegA11, respectively (Figure 3(A)). The only charged side chain-side chain interaction is between K224 of LegA11 and E118 of SidL_N_. Hydrophobic interactions involve residues L99 and L103 in SidL_N_ and L232, I236, F239, and F240 in LegA11. The interaction between SidL_C_ and LegA11 is mainly mediated by polar and charged interactions, whereas distinct hydrophobic interactions are absent (Figure 3(B)). In the following description, the respective residue of SidL_C_ will be mentioned first. Salt bridges are formed between the side chains of D393 and of K86 and the side chains of R396 and of E254, respectively. Side chain-main chain interactions are visible between the side chain of D393 and the main chain nitrogen of E258, as well as of R401 and the carbonyl oxygens of F239 and T242. Main chain-main chain interactions exist between the carbonyl oxygen of K357 and the nitrogen of E245, and the carbonyl oxygen of F389 and the nitrogens of G261 and I262. Figure 3.Binding interfaces of SidL and LegA11. Cartoon representation of the binding interfaces between SidL and LegA11. The color code corresponds to that in Figure 2. Side chains of amino acid residues responsible for mediating the interactions are depicted as stick models and are labeled accordingly. Hydrogen bonds and polar interaction are shown as gray dotted lines. (a) interface between SidL_N_ and LegA11. (b) interface between SidL_C_ and LegA11.3D structural model of the LegA11-SidLN and LegA11-SidLC interfaces. Several amino acid side chains at the interface are shown in two panels. In panel A, stick models of LegA11 (orange) residues L232, I236, F240, K224, K180 and stick models of SidLN (blue) residues L103, L99, E118, R121 on the blue protein Gray dotted lines connect specific residues, indicating polar contacts or hydrogen bonds as drawn. One gray dotted line connects orange K224 to blue E118. Another gray dotted line connects orange K180 to blue R121. No axes, scales, or numeric plots are present. In Panel B, stick models of LegA11 (orange) residues F239, T242, E245, E254, K86, E258, G261, I262 and SidLC (green) residues R401, K357, R396, D393, F389. Multiple gray dotted lines indicate contacts between labeled residues. At the upper left interface region, gray dotted lines connect green R401 to orange F239 and to orange T242 and a gray dotted line connects orange E245 to green K357. At the lower interface region, gray dotted lines connect green R396 to orange E254 and gray dotted lines connect green D393 to orange K86 and to orange E258. At the right side, a gray dotted line connects green F389 to orange I262. No axes, scales, or numeric plots are present.Two images of cartoon protein protein interfaces of SidL and LegA11 with labeled amino acid interactions.
To determine the binding affinity of SidL and LegA11, we employed isothermal titration calorimetry (ITC) using purified proteins. Based on the domain boundaries of SidL seen in the crystal structure, we recombinantly produced the variant SidL_76–628_, but found the protein to be contaminated with a degradation product. Extending the C-terminus to residue K645 yielded a stable protein that could be purified with high purity and yield, which was used in subsequent ITC experiments. We performed ITC measurements with SidL in the sample cell at concentrations between 1.45 and 0.61 µM and a 20-fold molar excess of LegA11 in the syringe. Averaging the individual measurements, we obtained a dissociation constant of 1.8 nM (Figure 4). Figure 4.Affinity of SidL and LegA11 determined by ITC. (a) baseline-corrected heat rate (µW) versus time (s) for titration of LegA11 (ligand) into SidL (receptor) at 25 °C, showing exothermic binding events (negative peaks) that diminish after saturation. (b) integrated binding isotherms from four independent experiments plotted as molar heat (kJ/mol) versus molar ratio of LegA11 to SidL. The best-fit one-site model yielded a dissociation constant Kd of 1.8 ^+4.5^ −1.3 nM, with a stoichiometry of 0.92 ± 0.16. Binding was strongly exothermic (average ΔH ≈ −200 kJ/mol). Receptor and ligand concentrations were: 1.45/32.77 µM, 0.72/14.97 µM, 0.63/10.61 µM, and 0.61/10.72 µM (SidL/lega11). C. Thermodynamic signature of binding showing large enthalpic contribution (ΔH = −194 ± 17 kJ/mol) opposed by an unfavorable entropic term (−TΔS = +144 ± 16 kJ/mol), resulting in a free energy change of ΔG = −50 ± 3 kJ/mol.The image contains three graphs panels labeled A, B and C. Panel A shows a line graph of heat rate versus time. The x-axis is labeled ‘Time in s‘, ranging from 0 to 8000 seconds. The y-axis is labeled ‘heat rate in microW‘, ranging from -0.5 to 0.1 microW. The graph shows a series of negative peaks that decrease in magnitude over time. Panel B shows a scatter plot with fitted curves of molar heat versus molar ratio. The x-axis is labeled ‘Molar ratio LegA11 / SidL‘, ranging from 0.0 to 2.0. The y-axis is labeled ‘Molar heat in kJ/mol‘, ranging from -250 to 25 kJ/mol. Four datasets are plotted with different symbols and colors, each representing different concentrations of SidL and LegA11. The fitted curves show a sigmoidal shape, indicating binding saturation. The dissociation constant Kd is reported as 1.8 plus 4.5 minus 1.3 nM. Panel C shows a bar graph of thermodynamic quantities. The x-axis shows three bars labeled delta H, minus T delta S and delta G. The y-axis is labeled ‘Contribution to delta G in kJ/mol‘, ranging from -250 to 200 kJ/mol. The delta H bar shows a large negative value around -200 kJ/mol, the minus T delta S bar shows a large positive value around 150 kJ/mol and the delta G bar shows a smaller negative value around -50 kJ/mol.Three graphs showing affinity of SidL and LegA11: heat rate vs time, molar heat vs molar ratio and thermodynamic quantities.
To verify the binding interface between SidL and LegA11 seen in the crystal structure, we introduced mutations into LegA11 to disturb complex formation. Given the large size of the interface, we decided to pursue a rather drastic approach by generating the quadruple mutant K95A/F239R/F240R/G243R. The three latter residues are located near the subdomain linker of SidL. We reasoned that introducing three charged long-chain residues into this tight hydrophobic pocket would increase the chances of disrupting the binding interaction between SidL and LegA11. We purified the resulting protein (LegA11-4 M) to homogeneity, and subjected SidL_76–645_ alone, LegA11 in wild-type or mutant form alone, or in equimolar ratio with SidL_76–645_ to analytical size exclusion chromatography (SEC). As expected, wild-type LegA11 formed a stable complex with the truncated SidL variant (Figure 5). The chromatogram corresponding to SidL_76–645_ and LegA11-4 M shows two distinct peaks very close to the positions of the single proteins. The slight shift to lower elution volumes most likely reflects a very weak transient interaction between SidL_76–645_ and LegA11-4 M. These data suggest that the interface observed in the crystal structure is indeed responsible for binding of LegA11 to SidL and that the interaction is largely abrogated by introduction of the mutations. Figure 5.Influence of mutations in LegA11 on binding to SidL_76–645_. 1 mg SidL_76–645_ and/or 400 µg LegA11 in 100 µL buffer were applied to an S200 10/300 increase size exclusion column. The colors of the respective chromatograms are as follows: SidL_76–645_ – black; LegA11-WT – blue; LegA11-4 M – dotted blue; SidL_76–645_/LegA11-WT – orange; SidL_76–645_/LegA11-4 M – dotted orange.A graph showing the influence of mutations in LegA11 on binding to SidL using size exclusion chromatography data.
LegA11 restores SidL-inhibited mRNA translation via a direct protein-protein interaction
Of the eight L. pneumophila effectors that block eukaryotic mRNA translation, two are associated with metaeffectors. The translation inhibiting effects of SidI are completely abolished in the presence of its metaeffector, MesI [14]. Therefore, we evaluated the effects of LegA11 on SidL-mediated translation inhibition. Recombinant His_6_-Myc-SidL and -LegA11 were purified from E. coli (Suppl. Fig S2) and added to cell-free rabbit reticulocyte lysates to quantify luciferase (Luc) mRNA translation. SidL inhibited Luc mRNA translation in a dose-dependent manner but with less potency than the effector SidI (Figure 6(A)), validating results from a prior study [9]. LegA11 binds and abrogates SidL toxicity in a heterologous yeast expression system [16]; thus, we tested the hypothesis that LegA11 suppresses SidL-mediated translation inhibition. Translation increased significantly when equimolar amounts of LegA11 and SidL were added to reactions and did not differ from reactions containing LegA11 alone (Figure 6(B)). Thus, LegA11 binds and suppresses SidL activity, further validating its role as a metaeffector of SidL. Figure 6.LegA11 restores SidL-mediated translation inhibition via a direct protein-protein interaction. Rabbit reticulocyte lysates were incubated with firefly luciferase (Luc) mRNA alone (positive) or with (a) 5 ng SidI, 5ng SidL, 50 ng SidL, or 100 ng SidL; (b) 65 nM of SidL, SidL and LegA11, or LegA11 alone; or (c) 65 nM of SidL alone, SidL and LegA11 variants, or LegA11 variants alone. Data shown are mean ± standard deviation (s.D.) of samples in triplicates (N = 3) and are representative of three independent experiments. AU, arbitrary units. Asterisks (*) denote statistical significance (***p < 0.001, *p < 0.05, ns, not significant) by one-way ANOVA with Tukey’s post-hoc test.The image contains three bar graphs labeled A, B and C, all measuring luminescence in arbitrary units (AU) on a logarithmic scale. The image A showing a bar graph with five conditions: Positive, 5 ng SidL, 5 ng SidL, 50 ng SidL and 100 ng SidL. The y-axis ranges from 10 superscript 2 to 10 superscript 7 AU. The positive control shows the highest luminescence, while 5 ng SidL shows the lowest. There is a dose-dependent increase in luminescence with increasing SidL concentration. The image B showing a bar graph with four conditions: Positive, SidL, SidL plus LegA11 and LegA11. The y-axis ranges from 10 superscript 4 to 10 superscript 7 AU. The positive control and LegA11 alone show high luminescence, while SidL alone shows low luminescence. The combination of SidL and LegA11 restores luminescence to levels similar to LegA11 alone. The image C showing a bar graph with six conditions: Positive, SidL, SidL plus LegA11, SidL plus LegA11-4M, LegA11 and LegA11-4M. The y-axis ranges from 10 superscript 3 to 10 superscript 6 AU. The positive control, LegA11 and LegA11-4M show high luminescence. SidL alone shows low luminescence, while combinations of SidL with LegA11 or LegA11-4M show intermediate luminescence levels. Statistical significance is indicated by asterisks above brackets connecting different conditions, with ns denoting not significant, single asterisk for p less than 0.05 and triple asterisks for p less than 0.001. The detailed data points are as follows: - Under the positive condition, A is 2.0 times 10 superscript 6 end superscript log10 AU, B is 5.0 times 10 superscript 6 end superscript log10 AU and C is 5.0 times 10 superscript 5 end superscript log10 AU. - Under the SidL 5 ng condition, A is 3.0 times 10 superscript 3 end superscript log10 AU, B is 2.0 times 10 superscript 5 end superscript log10 AU and C is 5.0 times 10 superscript 4 end superscript log10 AU. - Under the SidL 5 micrograms condition, A is 2.0 times 10 superscript 6 end superscript log10 AU, with no data for B and C. - Under the SidL 50 ng condition, A is 7.0 times 10 superscript 5 end superscript log10 AU, with no data for B and C. - Under the SidL 100 ng condition, A is 3.0 times 10 superscript 5 end superscript log10 AU, with no data for B and C. - Under the SidL plus LegA11 condition, there is no data for A, B is 3.0 times 10 superscript 6 end superscript log10 AU and C is 5.0 times 10 superscript 5 end superscript log10 AU. - Under the LegA11 condition, there is no data for A, B is 3.0 times 10 superscript 6 end superscript log10 AU and C is 5.0 times 10 superscript 5 end superscript log10 AU. - Under the SidL plus LegA11-4M condition, there is no data for A and B and C is 5.0 times 10 superscript 4 end superscript log10 AU. - Under the LegA11-4M condition, there is no data for A and B and C is 5.0 times 10 superscript 5 end superscript log10 AU.Three bar graphs showing luminescence measurements for different protein combinations and concentrations.
We hypothesized that SidL-LegA11 complex formation is required for functional regulation of SidL. To test this, we quantified Luc mRNA translation in cell-free lysates containing equimolar amounts of purified recombinant SidL with either wild-type LegA11 or LegA11-4 M (Suppl. Fig. S2), the latter of which has lost the ability to bind SidL (Figure 5). SidL-mediated translation inhibition was suppressed by wild-type LegA11 but unaffected by LegA11-4 M since the extent of translation inhibition did not differ from reactions containing SidL alone (Figure 6(C)). Luc mRNA translation was not affected by either wild-type LegA11 or LegA11-4 M alone (Figure 6(C)). Together, these data show that LegA11 regulates SidL via a direct protein-protein interaction and that SidL is unable to inhibit translation when it is bound to LegA11.
Neither SidL nor LegA11 impact L. pneumophila replication
He et al. found that neither SidL nor LegA11 individually influence L. pneumophila replication within primary bone marrow-derived macrophages (BMDMs) from susceptible A/J mice and the social amoebae, Dictyostelium discoideum [11]. D. discoideum is routinely used as a model to study L. pneumophila in the laboratory [36,37]; however, a single protozoan host is unable to capture the diversity of hosts and niches experienced by L. pneumophila in the environment [38,39]. To determine whether SidL or LegA11 influence L. pneumophila replication within a natural protozoan host, L. pneumophila strains with clean unmarked chromosomal deletions of legA11 or sidL (∆legA11 or ∆sidL, respectively) were generated to assess replication within Acanthamoeba castellanii. We found no differences in L. pneumophila ∆legA11 or ∆sidL growth relative to either wild-type L. pneumophila or genetically complemented strains in A. castellanii (Suppl. Fig S3A). We also found no differences in growth in AYE broth in vitro (Suppl. Fig. S3B). These findings support prior data suggesting that SidL and LegA11, like many L. pneumophila effectors, may function redundantly with other effectors in A. castellanii and/or have a more pronounced role in other L. pneumophila host(s) [38,39].
Discussion
Over the past decade, a growing number of metaeffectors have been identified and functionally characterized [12,14–16,18,40]. Metaeffectors are effectors that regulate cognate effector(s) by direct physical interactions [13], which is distinct from effector pairs that act in an opposing manner on the same host proteins and processes [41–43]. LegA11 was recently validated as a metaeffector of SidL [11,16], but the role of the SidL-LegA11 interaction in LegA11’s metaeffector activity was not fully established. Using purified proteins, we found that SidL and LegA11 bind with high affinity to form a stable binary complex, which prompted us to attempt a crystallographic characterization of the complex. Despite extensive crystallization trials using different N- and C-terminal truncation variants of SidL, we only obtained crystals after adding trace amounts of protease. This suggests that SidL contains one or more flexible regions that counteract crystal formation and is in line with the observation that SidL in the crystal structure is further truncated both N- and C-terminally than our shortest construct used for crystallization. Furthermore, a portion of 110 amino acid residues (aa 414–524) had been removed from SidL_C_ opposite the LegA11 binding site, which may indicate that this portion is flexible in the absence of a putative second binding partner.
SidL and LegA11 interact via a rather large interface of about 2300 Å^2^ involving mostly charged or polar side chains ([Figure 2 and 3](#f0002 f0003)). Although such an interface size suggests biological relevance [44], we wanted to verify that this interface is not a crystallization artifact. Indeed, mutagenesis of four interface residues in LegA11 (LegA11-4 M) disrupted the interaction between the two proteins. We found that LegA11-4 M was unable to form a complex with SidL (Figure 5), confirming the validity of the interface seen in the crystal structure (Figure 3(A,B)). The inability of LegA11-4 M to functionally regulate SidL in vitro (Figure 6(C)) suggests that LegA11’s metaeffector activity is mediated by a direct protein-protein interaction with SidL. A very recent study included a model of the complex between SidL and LegA11 generated with AlphaFold2 (He et al. 2025). The authors of this study described fourteen polar interactions between LegA11 and SidL. We compared these interactions to those found in our crystal structure. We could confirm the interactions of R17 and K180 from LegA11 with D549 and R121 from SidL, respectively. I2 and R6 are not resolved in our structure, which indicates that these residues do not form stable interactions within the complex. The remaining ten side chains of LegA11 are in the vicinity of their respective postulated interaction partners in SidL, but they are either not in hydrogen bonding distance or in a relative geometry that precludes hydrogen bond formation. This comparison suggests that AlphaFold2 correctly identified the interaction interface, but that details of the interaction on the level of single residues require experimental structural data.
SidL is one of at least eight Legionella effectors able to inhibit host mRNA translation in vitro [45–47]. The translation-inhibiting effector SidI (Lpg2504) is also regulated by a metaeffector, MesI (Metaeffector of SidI; Lpg2505). Like SidL and LegA11, SidI and MesI also form a high-affinity 1:1 complex that renders SidI nontoxic and unable to suppress mRNA translation [14]. Moreover, LegA11 and MesI both bind their cognate effectors – SidL and SidI, respectively – within L. pneumophila. The stability of these effector-metaeffector binary complexes may explain why less SidL and SidI are translocated into host cells when their cognate metaeffectors are present [11,48]. We found that LegA11’s metaeffector activity is mediated by a direct protein-protein interaction with SidL, but whether the same binding interface is responsible formation SidL-LegA11 regulation within living cells remains to be elucidated. We are currently testing whether LegA11-4 M can bind SidL within L. pneumophila and/or regulate SidL translocation into host cells.
A very recent study describes SidL as an ATPase that hydrolyzes ATP to AMP and pyrophosphate [11]. SidL binds ATP via an S-HxxxE motif [11], which is also present in toxins from phylogenetically diverse bacterial species [49]. He et al. [11] suggested that the inhibitory effect of LegA11 is exerted via binding to the C-terminal domain of SidL, where the active center is located [11]. To examine the spatial relationship between LegA11 and the ATP-binding site, we generated an AlphaFold3 (AF3) model of SidL complexed with ATP and superposed it with the SidL from our crystal structure. From the superposition it is apparent that LegA11 does not bind in the vicinity of the active site (Suppl. Fig. S4). Thus, it is unlikely that LegA11 abolishes ATP hydrolysis by directly occluding the nucleotide binding site of SidL. A structure of the effector RavJ in complex with its metaeffector LegL1 revealed that allosteric regulation is not a universal feature of metaeffectors LegL1 binds and occludes the active site of RavJ [16]. Thus, metaeffector regulatory mechanisms are diverse and can be defined experimentally through structural studies.
Despite functional similarities, SidL and SidI are mechanistically distinct. SidI is a mannosyltransferase that directly inactivates the host mRNA translation machinery [9,14]. Moreover, unlike SidL, the enzymatic activity of SidI is independent of host co-factors and requires only its activated sugar donor, GDP-mannose, for catalysis [14]. While SidI can function within L. pneumophila by itself [50], the requirement for host actin as a co-factor provides a plausible explanation for lack of discernable intrabacterial SidL activity (Suppl. Fig S3B). The mechanism of SidL-mediated translation inhibition has not been defined. He et al. suggest that SidL functions to deplete host cytosolic ATP pools [11], which could plausibly limit mRNA translation; however, it is also tempting to speculate a mechanistic connection between impaired actin polymerization and translation since F-actin serves as a scaffold for polyribosome assembly [9,51–55]. Sustained translation on monoribosomes may explain why SidL blocks translation with less potency than SidI (Figure 6A) [9].
SidL’s requirement for globular actin as an activating co-factor is similar to actin-mediated activation of the Legionella effector LnaB [11,49,56]. He et al. found that SidL binds LegA11 and actin simultaneously [11], which we corroborated using AF3 to generate a hypothetical ternary complex by overlaying SidL from our crystal structure with the SidL from the actin complex (Suppl Figure 5). Indeed, the binding sites of LegA11 and actin do not overlap, confirming the experimental findings of He et al. [11]. Interestingly, in all 15 AF3 predictions, SidL_C_ is sufficient for binding actin, whereas SidL_N_ is able to assume a wide variety of conformations (Suppl. Fig. S6). This is in line with the binding mode of actin in several crystal structures with LnaB, which does not possess a subdomain corresponding to SidL_N_. It is evident from our crystal structure that upon binding, LegA11 abolishes the conformational flexibility of SidL_N_ and locks it in a fixed orientation relative to SidL_C_. This conformational restriction is also likely reflected in the unusually high loss of entropy (−TΔS = 144 kJ/mol) seen in our ITC measurements (Figure 4).
A recent structural study showed that actin enhances the affinity of LnaB for its target phosphoribosyl-ubiquitin (PR-Ub) [57]. According to these authors’ model, the hydrolysis incompetent binary complex formed by LnaB and actin is activated by binding of PR-Ub. However, for SidL no third protein is required, since purified SidL and actin are sufficient for efficient ATP hydrolysis [11]. Therefore, actin must play a direct role in inducing hydrolytic competence. The most likely scenario is that binding of actin orders the large flexible loop (aa 414–524) of SidL, which at its C-terminal end borders on the nucleotide binding site. This reorientation may position loop residues required for nucleotide hydrolysis in a catalytically competent fashion. We hypothesize that the fixed N-terminal domain seen in our crystal structure prevents the correct positioning of the loop, effectively rendering SidL catalytically inactive despite concomitant binding of actin. However, elucidation of the precise mechanisms of both activation by actin and inhibition by LegA11 will require structural characterization of the respective complexes in the future.
Supplementary Material
Supplemental Information_v1.docx
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Gomez-Valero L, Buchrieser C. Intracellular parasitism, the driving force of evolution of Legionella pneumophila and the genus Legionella. Microbes Infect. 2019;21(5–6):230–14. doi: 10.1016/j.micinf.2019.06.01231252216 · doi ↗ · pubmed ↗
- 2Lockwood DC, Amin H, Costa TRD, et al. The Legionella pneumophila Dot/Icm type IV secretion system and its effectors. Microbiol (Read). 2022;168(5):168. doi: 10.1099/mic.0.00118735639581 · doi ↗ · pubmed ↗
- 3Mondino S, Schmidt S, Rolando M, et al. Legionnaires’ disease: state of the art knowledge of pathogenesis mechanisms of Legionella. Annu Rev Pathol. 2020;15(1):439–466. doi: 10.1146/annurev-pathmechdis-012419-03274231657966 · doi ↗ · pubmed ↗
- 4Romanov KA, O’Connor TJ. Legionella pneumophila, a Rosetta stone to understanding bacterial pathogenesis. J Bacteriol. 2024;206(12):e 0032424. doi: 10.1128/jb.00324-2439636264 PMC 11656745 · doi ↗ · pubmed ↗
- 5Belyi Y, Tabakova I, Stahl M, et al. Lgt: a family of cytotoxic glucosyltransferases produced by Legionella pneumophila. J Bacteriol. 2008;190(8):3026–3035. doi: 10.1128/JB.01798-0718281405 PMC 2293231 · doi ↗ · pubmed ↗
- 6Belyi Y, Jank T, Aktories K. Effector glycosyltransferases in Legionella. Front Microbiol. 2011;2:76. doi: 10.3389/fmicb.2011.0007621833323 PMC 3153043 · doi ↗ · pubmed ↗
- 7Moss SM, Taylor IR, Ruggero D, et al. A Legionella pneumophila kinase phosphorylates the Hsp 70 chaperone family to inhibit eukaryotic protein synthesis. Cell Host Microbe. 2019;25:454–462.e 6. doi: 10.1016/j.chom.2019.01.00630827827 PMC 6529236 · doi ↗ · pubmed ↗
- 8Syriste L, Patel DT, Stogios PJ, et al. An acetyltransferase effector conserved across Legionella species targets the eukaryotic e IF 3 complex to modulate protein translation. M Bio. 2024;15(3):e 0322123. doi: 10.1128/mbio.03221-2338335095 PMC 10936415 · doi ↗ · pubmed ↗