Effect of triazine thiols on steady-state mRNA levels in iPSC-derived hepatocytes
Carla Martinez-Morant, Jui-Tung Liu, Yu-Lin Jiang, Josef Blaszkiewicz, Stephen A. Duncan

TL;DR
This study shows that triazine thiols change mRNA levels in liver-like cells, suggesting a protective response.
Contribution
The study reveals specific mRNA changes in hepatocytes treated with triazine thiols, highlighting a protective mechanism.
Findings
145 mRNAs were reduced by ≥ 2-fold in triazine thiol-treated cells.
37 mRNAs were increased by ≥ 2-fold, including those for metallothionines.
The upregulation suggests a protective response through metal buffering.
Abstract
We previously reported that triazine thiols reduce apolipoprotein B (ApoB) secretion from human iPSC-derived hepatocytes (HLCs) and from humanized mice. To determine whether these compounds affected hepatocyte mRNA levels, we performed bulk RNA sequencing of HLCs treated with the triazine thiol DL-1 or with vehicle (DMSO) for 24 hours. Analyses revealed that in triazine thiol-treated cells, 145 mRNAs were reduced and 37 increased by ≥ 2-fold. Several mRNAs encoding cysteine-rich metallothionines were upregulated, implying that HLCs respond to treatment by mounting a protective response through metal buffering.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
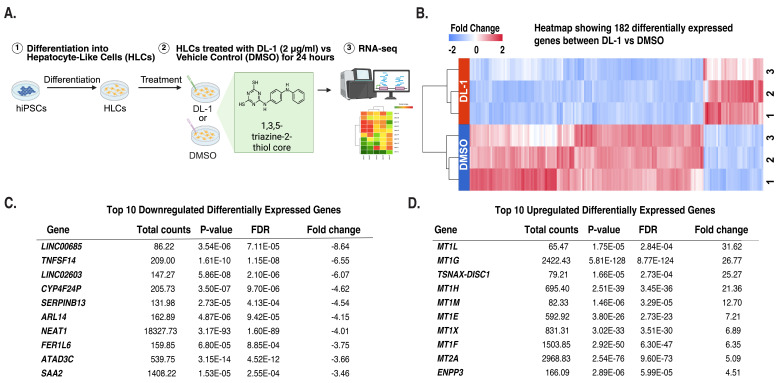 Figure 1
Figure 1|
Reagent / Resource |
Description |
Available from / Methods |
|
K3 human iPSCs |
Human male induced pluripotent stem cells derived from foreskin fibroblasts |
Si-Tayeb et al., 2010 |
|
mTeSR Medium |
Maintenance medium for human iPSCs |
|
|
Zebrafish basic fibroblast growth factor (bFGF) |
Supplement for iPSC maintenance (40 ng/mL) |
Ludwig et al., 2006 |
|
E-cadherin-IgG Fc fusion protein matrix |
Defined matrix for feeder-free iPSC culture |
Nagaoka et al., 2010 |
|
Matrigel (Geltrex) |
Basement membrane matrix (2 mg/mL) |
ThermoFisher Scientific, NY, #A1413302 |
|
RPMI 1640 Medium |
Basal medium for early differentiation |
ThermoFisher Scientific, NY, #22400089 |
|
B27 Supplement without insulin |
Supplement for definitive endoderm induction |
ThermoFisher Scientific, NY, #A1895601 |
|
B27 Supplement with insulin |
Supplement for hepatic progenitor and maturation stages |
ThermoFisher Scientific, NY, #17504044 |
|
Activin A |
Growth factor for definitive endoderm induction (100 ng/mL) |
ThermoFisher Scientific, NY, #PHC9563 |
|
Fibroblast Growth Factor 2 (FGF2) |
Growth factor for hepatic specification (20 ng/mL) |
ThermoFisher Scientific, NY, #PHG0023 |
|
Bone Morphogenetic Protein 4 (BMP4) |
Growth factor for hepatic progenitor induction (20 ng/mL) |
ThermoFisher Scientific, NY, #PHC9533 |
|
Hepatocyte Growth Factor (HGF) |
Growth factor for immature hepatocyte differentiation (20 ng/mL) |
Invitrogen, MA, USA, #PHG0321 |
|
Hepatocyte Culture Medium (HCM) |
Maturation medium for hepatocyte-like cells |
Lonza, MD; #CC3198 |
|
Oncostatin M |
Cytokine for hepatocyte maturation (20 ng/mL) |
Invitrogen, MA, USA, #PHC5015 |
|
DL-1 |
Triazine-2-thiol compound used for treatment (2 µg/mL) |
Liu et. al. , 2023 |
|
Dimethyl sulfoxide (DMSO) |
Vehicle control for DL-1 treatment |
Sigma-Aldrich #67-68-5 |
|
RNeasy Plus Mini Kit |
RNA extraction kit |
QIAGEN #74136 |
|
Partek Genomics Suite v12.3.1 |
RNA-seq data processing and normalization |
Illumina San Diego, CA 92122 USA. |
|
STAR aligner |
RNA-seq read alignment to human genome (hg38) |
Illumina San Diego, CA 92122 USA. |
|
DESeq2(R) |
Differential gene expression analysis |
Illumina San Diego, CA 92122 USA. |
|
Enrichr |
Pathway enrichment analysis (GO BP 2025, Reactome 2024) |
Maayan Lab, Icahn School of Medicine, NY |
|
Science Machine AI platform |
Bioinformatic Analysis Platform |
Science Machine, Inc. (2026) |
|
RNA-seq data |
This study |
GEO: GSE318437 |
- —National Institute of Diabetes and Digestive and Kidney Diseases (United States)https://ror.org/00adh9b73
- —National Institute of Diabetes and Digestive and Kidney Diseases (United States)https://ror.org/00adh9b73
- —National Institute of Diabetes and Digestive and Kidney Diseases (United States)https://ror.org/00adh9b73
- —National Institute of General Medical Sciences (United States)https://ror.org/04q48ey07
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsSulfur Compounds in Biology · Synthesis and Characterization of Heterocyclic Compounds · Redox biology and oxidative stress
Description
The liver plays a central role in regulating plasma lipid levels by synthesizing and secreting apolipoprotein B (ApoB), which is the key protein component of low-density lipoproteins (LDL) (Whitfield et al., 2004). Excessive LDL secretion contributes to familial hypercholesterolemia (FH), which is caused by mutations in the LDL receptor (Hopkins et al., 2011). Therapeutic approaches that target ApoB synthesis or lipoprotein assembly are effective but cause excessive lipid accumulation within hepatocytes (Rader & Kastelein, 2014; Cayo et al., 2012). We previously identified DL-1, a small molecule characterized by a 1,3,5-triazine-2-thiol core (Liu et al., 2023). This molecule has therapeutic potential for FH and other dyslipidemias due to its ability to reduce ApoB secretion in induced pluripotent stem cell-derived hepatocyte-like cells (HLCs) and in humanized mice without causing lipid buildup.
We previously demonstrated that triazine thiols do not affect intracellular steady-state *APOB mRNA * or protein levels but instead affect APOB secretion (Liu et al., 2023). Although the target of the triazine thiols is unknown, the absence of an effect on APOB mRNA levels suggested that the triazine thiols did not directly impact transcription. Nevertheless, we felt it important to determine whether triazine thiols had a substantial effect on hepatocyte gene expression more broadly. We therefore performed bulk RNA sequencing on HLCs treated with either DL-1 at 2 µg/mL or with a DMSO vehicle control for 24 hours ( Figure 1A ). The experiment compared biological triplicates for each condition ( Figure 1B ). We selected this compound concentration because it corresponds to the EC 50 for DL-1 (Liu et al., 2023). The effect of the compound was measured after 24 hrs, consistent with our previous analyses, which showed reduced APOB secretion at this time point (Liu et al., 2023).
Transcriptome analysis identified 182 genes whose steady-state mRNA levels were changed following DL-1 treatment (FDR ≤ 0.001, p-value ≤ 0.001, Fold change [-2,2]) ( Figure 1B ; see Extended Data File for full list of Differentially Expressed Genes (DEGs)). Of these differentially expressed genes 37 were increased and 145 were reduced. Ontology analyses revealed that genes with reduced mRNA levels exhibited diverse biological effects, with little enrichment for specific pathways or processes. The top 10 reduced genes are shown in Figure 1C . In contrast to downregulated genes, 37 genes whose steady-state mRNA levels increased were highly enriched for metallothionein genes (MT1L, MT1G, MT1H, MT1M, MT1E, MT1X, MT1F, and MT2A) (Figure 2D). This profile is characteristic of metal-responsive transcription factor 1 (MTF-1) activation, which occurs when cells’ intracellular zinc or other divalent cations are perturbed. Given DL-1’s triazine-2-thiol structure, this increase in metallothionine expression could potentially be explained by the compound’s thiol structure chelating zinc or other divalent metal cations, depleting free intracellular pools, thereby triggering an increase in metallothioneins.
Methods
Cell Culture and Differentiation
Cell cultures were routinely screened for mycoplasma. Human male K3 iPSCs were generated from foreskin fibroblasts (ATCC CRL2097), and their detailed characterization, karyotyping, and short tandem repeat (STR) analyses have been previously described (Si-Tayeb, Noto, Sepac, et al., 2010). iPSCs were cultured in mTeSR medium supplemented with 40 ng/ml zebrafish basic fibroblast growth factor (Ludwig et al., 2006) on an E-cadherin-IgG Fc fusion protein matrix in 4% O2-5% CO2 incubators (Nagaoka et al., 2010). To initiate differentiation, K3 cells were seeded as a monolayer on Matrigel (2 mg/ml)-coated tissue culture plates for a full 24 hours prior. We induced these cells to differentiate into hepatocyte-like cells following an established protocol (Si-Tayeb, Noto, Nagaoka, et al., 2010). In the initial two days of differentiation, the cells are cultured in RPMI 1640 Medium (Invitrogen, MA, #22400105), enriched with 2% B27 Supplement without insulin (Invitrogen, MA, #A1895601), 100 ng/mL Activin A (Invitrogen, MA, #PHC9563), 20 ng/mL Fibroblast Growth Factor 2 (FGF2) (Invitrogen, MA, #PHG0023), and 10 ng/mL BMP4 (Invitrogen, MA, #PHC9533). Over the next 3 days, the cells were induced to form definitive endoderm by culturing in B27 Supplement without insulin and 100 ng/mL Activin A. Over the next 5 days, the cells were treated with B27 Supplement containing insulin, 10 ng/mL FGF2, and 20 ng/mL BMP4 to generate hepatic progenitor cells. The cells then received an additional 5 days of B27 Supplement and 20 ng/mL Hepatocyte Growth Factor (Invitrogen, MA, #PHG0321) to generate immature hepatocytes. Finally, the cells were cultured in HCM medium (Lonza, MD, #CC3198) and supplemented with 20 ng/mL Oncostatin M (Invitrogen, MA, #PHC5015) for the final 5 days of differentiation, thereby inducing hepatocyte-like cell differentiation (Mallanna & Duncan, 2013).
DL-1 Treatment
On day 20 of differentiation, cells were treated with DL-1 (2 µg/mL) or vehicle control (DMSO) in maturation medium for 24 hours, with three biological replicates per condition.
RNA Extraction and Sequencing
After treatment, cells were washed with PBS, and RNA was extracted using the RNeasy Plus Mini Kit (QIAGEN). High-throughput sequencing was performed by Beijing Genomics (Shenzhen, China), which confirmed RNA integrity and performed library preparation (Illumina).
Bioinformatics Analysis
RNA-seq bioinformatics analysis was performed on FASTQ files using Partek Genomics Suite version 12.3.1. STAR was used to align the reads, and the resulting alignments were annotated to the human genome (hg38). Counts per Million (CPM) were used for normalization. DEseq2 (R) was used to conduct differential expression analysis and statistical significance was reached at p-value (≤ 0.001), FC (-2,2), and FDR (≤ 0.001). Pathway Enrichment Analysis was performed on the DEGs using Enrichr against GO Biological Processes 2025 and Reactome Pathways 2024. Heatmaps were generated using Science Machine, Inc. (2026).
The RNA-seq dataset generated in this study has been deposited in GEO (GSE318437).
Reagents
**: **
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Cayo Max A. Cai Jun De La Forest Ann Noto Fallon K. Nagaoka Masato Clark Brian S. Collery Ross F. Si-Tayeb Karim Duncan Stephen A. 2012917 JD induced pluripotent stem cell-derived hepatocytes faithfully recapitulate the pathophysiology of familial hypercholesterolemia Hepatology 5660270-91392163217110.1002/hep.2587122653811 PMC 3900031 · doi ↗ · pubmed ↗
- 2Hopkins Paul N. Toth Peter P. Ballantyne Christie M. Rader Daniel J. 201161 Familial Hypercholesterolemias: Prevalence, genetics, diagnosis and screening recommendations from the National Lipid Association Expert Panel on Familial Hypercholesterolemia Journal of Clinical Lipidology 531933-2874 S 9S 1710.1016/j.jacl.2011.03.45221600530 · doi ↗ · pubmed ↗
- 3Liu Jui-Tung Doueiry Caren Jiang Yu-lin Blaszkiewicz Josef Lamprecht Mary Paige Heslop James A. Peterson Yuri K. Carten Juliana Debrito Traktman Paula Yuan Yang Khetani Salman R. Twal Waleed O. Duncan Stephen A. 2023424 A human i PSC-derived hepatocyte screen identifies compounds that inhibit production of Apolipoprotein B Communications Biology 612399-364210.1038/s 42003-023-04739-9PMC 1012597237095219 · doi ↗ · pubmed ↗
- 4Ludwig Tenneille E Bergendahl Veit Levenstein Mark E Yu Junying Probasco Mitchell D Thomson James A 2006721 Feeder-independent culture of human embryonic stem cells Nature Methods 381548-709163764610.1038/nmeth 90216862139 · doi ↗ · pubmed ↗
- 5Mallanna Sunil K. Duncan Stephen A. 201391 Differentiation of Hepatocytes from Pluripotent Stem Cells Current Protocols in Stem Cell Biology 2611941-732210.1002/9780470151808.sc 01g 04s 26PMC 392029424510789 · doi ↗ · pubmed ↗
- 6Nagaoka Masato Si-Tayeb Karim Akaike Toshihiro Duncan Stephen A 201062 Culture of human pluripotent stem cells using completely defined conditions on a recombinant E-cadherin substratum BMC Developmental Biology 1011471-213X 10.1186/1471-213x-10-60PMC 289693720525219 · doi ↗ · pubmed ↗
- 7Rader Daniel J. Kastelein John J.P. 201434 Lomitapide and Mipomersen Circulation 12990009-73221022103210.1161/circulationaha.113.00129224589695 · doi ↗ · pubmed ↗
- 8Si-Tayeb Karim Noto Fallon K. Nagaoka Masato Li Jixuan Battle Michele A. Duris Christine North Paula E. Dalton Stephen Duncan Stephen A. 201011 Highly Efficient Generation of Human Hepatocyte–Like Cells From Induced Pluripotent Stem Cells Hepatology 5110270-913929730510.1002/hep.2335419998274 PMC 2946078 · doi ↗ · pubmed ↗