Harnessing evolution: leveraging bacterial isoprenoid pathway diversity toward improved bioengineering strategies
Christine M. Qabar, Bailey A. Marshall, Robert Landick, Jeffery S. Cox

TL;DR
This paper explores how bacteria use different isoprenoid pathways, revealing diversity that could help engineer microbes for sustainable production of valuable compounds.
Contribution
The study highlights rare bacteria with unique isoprenoid pathway configurations and their potential for bioengineering.
Findings
Bacteria show significant diversity in isoprenoid pathway usage across and within genera.
Some bacteria encode both or neither isoprenoid pathway, suggesting novel evolutionary adaptations.
MEP intermediates may have roles beyond isoprenoid synthesis, influencing pathway evolution.
Abstract
Isoprenoids play vital roles in all domains of life, from beta-carotene in bacteria to heme in humans. Two distinct metabolic pathways have evolved to synthesize the critical precursor of all mature isoprenoids: the mevalonate (MEV) and the methylerythritol phosphate (MEP) pathways. Here, we quantify the extensive inter- and intra-genus heterogeneity in the usage of these two pathways with particular emphasis on rare bacteria that encode both, or neither, pathways. Furthermore, MEP intermediates themselves have non-isoprenogenic roles that may underlie evolutionary pressures driving pathway diversification. Understanding isoprenoid biosynthesis in bacteria offers new avenues toward more sustainable engineering of economically relevant molecules in microbes.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
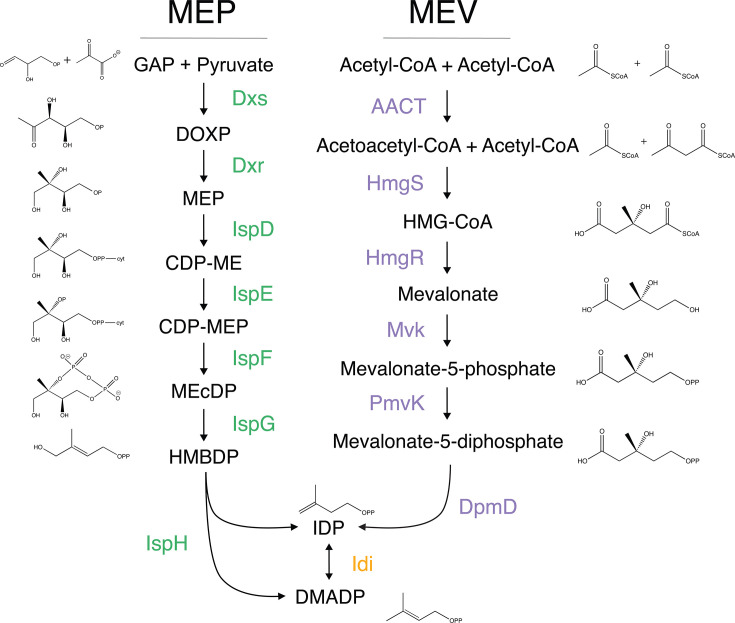 Fig 1
Fig 1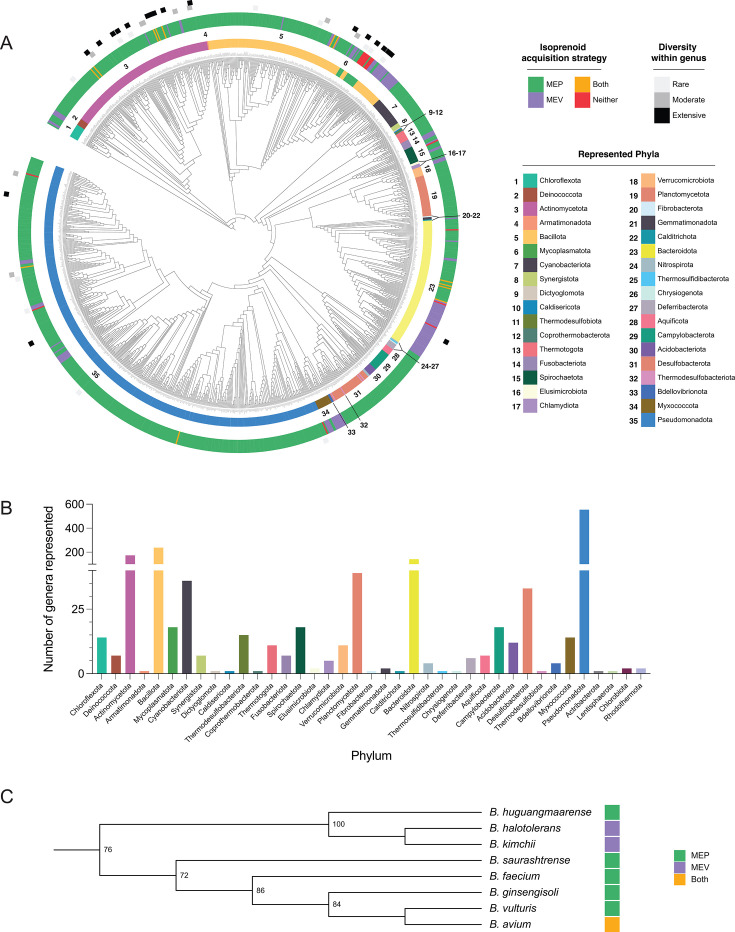 Fig 2
Fig 2| Pathway | Enzyme | Reaction | Other features |
|---|---|---|---|
| MEP pathway | DOXP synthase | Condensation of pyruvate + D-glyceraldehyde 3-phosphate → 1-deoxy-D-xylulose 5-phosphate ( | Cofactors: Mg2+, thiamin diphosphate (TDP). Rate-limiting step of MEP. DOXP can be shunted to thiamin, pyridoxal biosynthesis ( |
| DOXP reductoisomerase | Reversible rearrangement from DOXP → 2-C-methylerythritol 4-phosphate ( | Cofactors: divalent cation (Co2+, Mn2+, Mg2+), NADPH. Types I and II Dxr have scattered taxonomic distribution ( | |
| MEP cytidylyltransferase | MEP → 4-diphosphocytidyl-2-C-methylerythritol ( | Cofactors: CTP, Mg2+. IspDF fusion proteins are common, despite catalyzing nonconsecutive steps ( | |
| CDP-ME kinase | CDP-ME → 4-diphosphocytidyl- 2-C-methyl-D-erythritol 2-phosphate ( | Cofactor: ATP. | |
| MEcDP synthase | CDP-MEP → 2-C-methyl-D-erythritol 2,4-cyclodiphosphate ( | Cofactors: Zn2+ AND Mg2+. Farnesyl diphosphate (FDP) inhibits | |
| HMBDP synthase | MEcDP→ (E)−4-Hydroxy-3-methyl-but-2-enyl diphosphate ( | Cofactors: 4Fe-4S, NADPH, reducing equivalent (flavodoxin). HMBDP activates Vγ9Vδ2 T cells ( | |
| HMBDP reductase | HMBDP → isopentenyl diphosphate ( | Cofactors: FAD, divalent cations (Co2+), 4Fe-4S, NADPH, reducing equivalent (flavodoxin or ferredoxin). Makes 5:1 IDP:DMADP ( | |
| Both | Isopentenyl diphosphate isomerase | Isomerization of IDP ⇌ DMADP | Cofactors: Zn2+, Mg2+ ( |
| MEV pathway | Acetoacetyl-CoA thiolase | Condensation of 2 acetyl-CoA → acetoacetyl-CoA | |
| HMG-CoA synthase | Condensation of acetoacetyl-CoA + acetyl-CoA → 3-hydroxy-3-methylglutaryl-CoA ( | Regulated by sterols ( | |
| HMG-CoA reductase | HMG-CoA → mevalonate | Rate-limiting step of MEV. Regulated by sterols ( | |
| Mevalonate kinase | Mevalonate → mevalonate-5-phosphate | Cofactors: Mg2+, ATP. Feedback is inhibited by downstream isoprenoids (class I) ( | |
| Phosphomevalonate kinase | Mevalonate-5-phosphate → mevalonate-5-diphosphate | Cofactors: Mg2+, ATP. | |
| Diphosphomevalonate decarboxylase | Mevalonate-5-diphosphate → IDP | Cofactors: Mg2+, ATP. Generates only IDP, unlike the terminal enzyme of the MEP pathway. |
- —National Institutes of Healthhttp://dx.doi.org/10.13039/100000002
- —National Institutes of Healthhttp://dx.doi.org/10.13039/100000002
- —National Institutes of Healthhttp://dx.doi.org/10.13039/100000002
- —U.S. Department of Energyhttp://dx.doi.org/10.13039/100000015
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant biochemistry and biosynthesis · Microbial Metabolic Engineering and Bioproduction · Plant Gene Expression Analysis
ISOPRENOIDS ARE UBIQUITOUS AND ESSENTIAL ACROSS THE DOMAINS OF LIFE
Isoprenoids are a massive and diverse class of biomolecules
Isoprenoids, also referred to as terpenoids, are a broad and diverse class of molecules, including ~95,000 different compounds that have been identified to date (1, 2). Defined by repeating five-carbon isoprene units, these compounds play crucial roles in various processes in all domains of life, including ATP synthesis, protein modification, hormone signaling, maintenance of membrane structure and fluidity, pigment production, and stress response (3, 4). Key isoprenoids produced in animals include heme, cholesterol, steroids, and vitamin D, as well as post-translational prenyl or farnesyl modifications (5, 6). Furthermore, archaeal membranes contain essential isoprenoid components (7), and fungal isoprenoids are pharmacologically valuable as antimicrobials and antitumors (8). In addition to mediating basic cellular functions, plant isoprenoids have been used in indigenous cultures for thousands of years and are now being isolated for use in medicine, agriculture, and cosmetics (9). Although isoprenoids are integral in all domains of life, we will focus on bacteria as this kingdom contains the most diverse range in isoprenoid biosynthesis strategies. Importantly, this existing natural diversity can be harnessed in ongoing efforts to bioengineer isoprenoids at scale in bacteria.
Isoprenoids participate directly and indirectly in myriad bacterial processes
In bacteria, isoprenoids directly support both vital cellular processes, such as cell wall synthesis and electron transport, as well as secondary bacterial behaviors, including intermicrobial signaling and host-microbe interactions (3, 4, 10). For example, the critical electron carriers, ubiquinone and menaquinone, have isoprenoid modifications, and isoprenoid lipids, called hopanoids, have been shown to support resistance to pH, bile salt, and antibiotic stress in some bacteria (4). For further reading on the direct effects of isoprenoid biosynthesis in bacterial pathogens and more broadly in microbial ecology, we recommend the reviews of Heuston et al. (11) and Avalos et al. (4).
Surprisingly, recent work has shed light on the important indirect effects of biosynthetic intermediates on cells, which mediate non-isoprenogenic interactions that are independent of isoprenoid biosynthesis (12, 13). Understanding non-isoprenogenic roles of isoprenoid intermediates expands our understanding of the global cellular effects of isoprenoid biosynthesis, as well as the forces guiding pathway evolution. For example, MEcDP and HMBDP are small molecule intermediates of isoprenoid biosynthesis (Fig. 1) that have been implicated in non-isoprenogenic processes such as oxidative stress sensing and host immune sensing, further discussed next.
Two pathways of isoprenoid biosynthesis in bacteria. Shown are intermediates and enzymes of the MEV (purple) and MEP (green) pathways of isoprenoid biosynthesis. Orange indicates that the enzyme is found in both pathways. GAP, glyceraldehyde 3-phosphate; Dxs, DOXP synthase; DOXP, 1-deoxyxylulose 5-phosphate; Dxr, DOXP reductoisomerase; MEP, 2-C-methyl-erythritol 4-phosphate; IspD, 2-C-methyl-D-erythritol 4-phosphate cytidylyltransferase; CDP-ME, 4-(cytidine 5′-diphospho)−2-C-methyl-erythritol; IspE, 4-diphosphocytidyl-2-C-methyl-D-erythritol kinase; CDP-MEP, 2-phospho-4-(cytidine 5′-diphospho)−2-C-methyl-erythritol; IspF, 2-C-methyl-D-erythritol 2,4-cyclodiphosphate synthase; MEcDP, 2-C-methyl-erythritol-2,4-cyclodiphosphate; IspG, 4-hydroxy-3-methylbut-2-en-1-yl diphosphate synthase; HMBDP, 4-hydroxy-3-methyl 2-butenyl diphosphate; IspH, 4-hydroxy-3-methylbut-2-enyl diphosphate reductase; IDP, isopentenyl diphosphate; Idi, isopentenyl diphosphate isomerase; DMADP, dimethylallyl diphosphate; AACT, Acetoacetyl-CoA thiolase; HmgS, HMG-CoA synthase; HMG-CoA, hydroxymethylglutaryl-CoA; HmgR, HMG-CoA reductase; Mvk, mevalonate kinase; PmvK, phosphomevalonate kinase; and DpmD, diphosphomevalonate decarboxylase.
THE MEP AND MEV PATHWAYS FOR ISOPRENOID BIOSYNTHESIS
Bacteria can use two pathways to synthesize isoprenoids
Isopentenyl diphosphate (IDP) and its isomer dimethylallyl diphosphate (DMADP) are the five-carbon precursor units used to build all isoprenoids. Two distinct pathways exist to generate these precursors: the mevalonate (MEV, sometimes referred to as the MVA) pathway and the methylerythritol phosphate (MEP, also known as the non-mevalonate) pathway (Fig. 1). Although the MEP and MEV pathways both generate the same end products, they are not simply interchangeable; they proceed via different enzymatic steps and thus have unique cofactor and energetic requirements, as well as unique non-isoprenogenic effects from metabolic intermediate interactions (Table 1). For example, the MEP pathway is more carbon-efficient as it requires fewer glyceraldehyde-3-phosphate molecules per IDP/DMADP produced, while the MEV pathway is more energy-efficient because it generates a net yield of reducing equivalents (14, 15). The distribution of these pathways has largely been regarded as mutually exclusive, barring a few exceptions; however, in this review, we bring into focus the extensive diversity of pathway utilization even within groups of closely related bacteria (Fig. 2). Understanding the evolutionary forces that influence pathway usage may inform engineering efforts that aspire to maximize isoprenoid production for sustainable human use, which is explored in “Synthetic biology approaches to engineer isoprenoid biosynthesis,” below.
Extensive diversity in isoprenoid acquisition strategies among bacteria. (A) The identity of isoprenoid biosynthesis pathways encoded by each genus is shown in the outer ring: MEP (green), MEV (purple), neither (red), or both (orange). Diversity within a genus is indicated by squares in light gray (rare exceptions, <10% of total species diverge from the primary genus call), dark gray (moderate diversity, 10%–25% of the total species diverge from the genus call), or black (extensive diversity, >25% of total species diverge from the genus call). Phyla are indicated by the numbered inner ring. The data shown represent 1,033 unique genera and 36 unique phyla. The Genome Taxonomy Database (GTDB) Bac120 tree (16) was visualized using iTol v7 (17) and pruned to remove unnamed isolates and display one node per genus. The NCBI Taxonomy and GTDB databases were used to assign phyla, and previously published homology identification (18) was used to make isoprenoid strategy and diversity calls. iTol Tree ID 108654132457551738254768. (B) Genus-level representation in pathway analysis. Table S1 is an expanded, species-level analysis of isoprenoid pathway usage. Analysis includes 4,249 species, representing 1,412 genera and 39 phyla. (C) Brachybacterium, within the phylum Actinomycetota, contains species with different isoprenoid biosynthesis strategies. As in panel A, the pathways encoded by each species are noted in green (MEP), purple (MEV), or orange (both). 16s rRNA sequences were acquired from the NCBI Nucleotide database (19). Geneious Prime (v2024.0.7) was used to perform a MUSCLE alignment (20) and build a PhyML tree (21) with 50 bootstraps. Annotation and tree visualization were performed on iTol (17). iTol Tree ID 10865413219911738351554.
The MEP pathway is the predominant bacterial isoprenoid biosynthesis pathway
The MEP pathway of isoprenoid biosynthesis is found in most but not all bacteria (11, 18, 46), as well as in plant plastids and apicomplexan parasites like Toxoplasma gondii and Plasmodium spp (27). Interestingly, plants have both pathways spatially separated in organelles, with cytosolic IDP production occurring via the MEV pathway and plastidal synthesis occurring via the MEP pathway, a segregation likely originating from an early bacterial endosymbiosis event (38, 47, 48). Furthermore, apicomplexans only encode the MEP pathway in the genome of their apicoplast, an organelle that is cyanobacterial in origin (49). The mechanistic studies that identified MEP enzymes have been reviewed in detail (32, 50–52), and the regulation of this pathway has been most often reviewed in plants (53). The persistence of both isoprenoid biosynthesis pathways across the kingdoms of life and the partitioning of the MEP pathway within organelles indicate selective pressure for each pathway in certain conditions.
Isoprenoid biosynthesis via the MEP pathway generates both IDP and DMADP via seven enzymatic steps and uses 2–3 NADPH depending on isozyme usage, 1 CTP, 1 ATP, and a reducing partner, which may be ferredoxin or flavodoxin (Fig. 1). This pathway is heavily regulated through a network of feedback mechanisms but has a higher theoretical yield than the MEV pathway (54). The tightly regulated MEP pathway branches off from glycolysis via the condensation of pyruvate and D-glyceraldehyde 3-phosphate to produce DOXP by the enzyme Dxs. This is the rate-limiting step of the pathway and is feedback inhibited by IDP/DMADP binding of Dxs, thus serving as the main control point of MEP flux (23, 55). Next, Dxr rearranges DOXP to form MEP, the first committed intermediate for which the pathway is named. The subsequent three steps mediated by IspD, IspE, and IspF activate and cyclize MEP to form MEcDP, an important intermediate which is discussed later in this section. The FeS cluster-containing protein IspG (also referred to as GcpE) then catalyzes the conversion of MEcDP to HMBDP, an intermediate that potently stimulates an immune response in the context of infection (30). IspH (also referred to as LytB) then converts HMBDP to IDP and DMADP at a ratio favoring IDP, although this ratio is variable based on the enzyme of study (34). Understanding this diversity in enzyme function is of interest from a bioengineering perspective; however, the mechanistic underpinnings of this phenomenon are not well understood. Interestingly, while the first step catalyzed by Dxs is rate-limiting, both IspG and IspH are considered important pathway bottlenecks due to their redox-sensitive FeS clusters (56, 57). Finally, IDP and DMADP can be interconverted via the enzyme Idi (Fig. 1).
The MEV pathway is used by some bacteria and is predominant in eukaryotes and archaea
The mevalonate pathway of isoprenoid biosynthesis has been reviewed extensively in eukaryotes (5, 39, 58) and archaea (59, 60) and is of particular interest in human medicine, as statin drugs that target this pathway are routinely used to lower cholesterol and synergize with cancer therapy (61). In the bacterial kingdom, the MEV pathway is less ubiquitous than the MEP pathway but is still used by some bacteria, including Borrelia burgdorferi, Staphylococcus aureus, and Streptococcus pneumoniae (39).
The MEV pathway consists of six enzymatic steps and uses two NADPHs and three ATPs (Fig. 1). First, two acetyl-CoA molecules are condensed to produce acetoacetyl-CoA by the enzyme acetoacetyl-CoA thiolase. HmgS then condenses acetoacetyl-CoA with another molecule of acetyl-CoA to generate Hmg-CoA, which is reduced by the rate-limiting enzyme HmgR to produce mevalonate, for which this pathway is named (62). Two sequential phosphorylation reactions by the kinases Mvk and Pmvk generate mevalonate-5-diphosphate, which is then converted to IDP by the enzyme Dpmd. In contrast to the MEP pathway, the MEV pathway generates only IDP and requires Idi to isomerize IDP to DMADP.
There is a eukaryotic/bacterial-type MEV pathway and an archaeal-type MEV pathway, which differ in the intermediate steps taken, but ultimately both lead to the generation of IDP and DMADP (40, 63). Furthermore, while the bacterial MEV pathway is most similar to that of eukaryotes, some members of the phylum Chloroflexota encode an archaeal-type MEV pathway (64, 65), supporting a model in which multiple mechanisms of MEV gene acquisition occurred in bacteria. Clearly, there exists considerable heterogeneity even among organisms that use the MEV pathway, presenting a fascinating question: how did this pathway evolve?
Origins and evolution of isoprenoid biosynthesis pathways
The evolutionary origin of isoprenoid biosynthetic pathways remains controversial. It is widely accepted that the MEV pathway is ancestral in all domains of life and that the MEP pathway subsequently evolved in bacteria. However, there is disagreement on whether MEV emerged in the last universal common ancestor or, as new phylogeny work suggests, from the newly discovered Asgardarchaeota phylum, which is thought to be ancestral to eukaryotes (63). Even in bacteria, the origin of the MEV pathway remains unresolved. Some suggest that certain bacterial lineages retained MEV genes from the last universal common ancestor (58), while others argue that these genes were horizontally acquired from eukaryotes (3).
While the precise origin of the bacterial MEV pathway remains elusive, the evolutionary course of isoprenoid biosynthesis in bacteria likely unfolded through multiple routes from potentially multiple ancestors. What is clear is that there is considerable genetic, metabolic, and enzymatic diversity to all arrive at the same molecule: IDP. The distinct regulation and energetic requirements of these two pathways were likely critical drivers of this diversification. However, it is possible that the variability in the non-isoprenogenic roles of each pathway, especially of their intermediates, has played a role in the evolutionary pressure to develop multiple routes for isoprenoid biosynthesis.
Gene- and protein-level diversity
Our understanding of the evolutionary course of isoprenoid biosynthesis pathways is complicated by the phylogenies of individual enzymes. For example, HmgR, primarily found in archaea, eukaryotes, and some bacteria (type I), is distinct from those present mostly in bacteria, and a few archaea (type II), and the two are differentially susceptible to statin drugs (10, 58).
Idi presents another node of variability in both the MEP and MEV pathways. It is the only shared enzyme between both the MEV and MEP pathways and is found in two forms that are structurally unrelated and have different cofactor requirements (36). Bacteria possess either the eukaryotic type I Idi or the archaeal type II Idi, with no correlation between pathway usage and Idi type (10). Some bacteria, including Mycobacterium marinum, encode both type I and II Idi enzymes (36).
Gene-level diversity also exists in the MEP pathway, further demonstrating considerable flexibility within this essential biosynthetic process. The first committed step of the MEP path is carried out by the enzyme Dxr, which exists in two forms: Dxr-I and Dxr-II, which only share sequence-level homology at the NADPH-binding domain (10). Type II Dxr is sporadically distributed across the bacterial kingdom in a manner that cannot be easily explained by HGT and instead is thought to have occurred through gene loss from a complex ancestor (66). No defining pattern of Dxr distribution among bacteria has emerged other than overrepresentation among certain pathogens (25). The existence of gene-level variance in both MEP and MEV genes suggests that these pathways are quite flexible, allowing for specialization and survival in a broad range of niches.
There are instances of organisms that use enzymatic steps that are slightly different from the canonical pathways to generate IDP (40, 67). These alternative “routes” still possess the namesake metabolic intermediate of each pathway, methylerythritol phosphate (MEP) or mevalonate (MEV). For example, three routes through the MEV pathway have been identified in archaea, including some with feedback-resistant enzymes, suggesting divergent mechanisms of regulation across the domains of life (68–70). Regarding the MEP pathway, several mutations outside of the MEP pathway have been identified that allow for the suppression of Dxr- and Dxs-knockout lethality, suggesting that there are functional, non-canonical routes through this pathway (71–73). Taken together, this flexibility indicates that although isoprenoid biosynthesis is essential, these pathways are highly adapted to the metabolic demands of a given niche.
Although the ancestral emergence of the MEV pathway is incompletely resolved, the current diversity of isoprenoid biosynthesis strategies reflects the breadth of metabolic requirements and capacities in bacteria. This is especially apparent in the distribution of these strategies across the bacterial kingdom (Fig. 2A). Furthermore, the heterogeneity of pathway types, enzyme classes, and evolutionary routes clearly demonstrates the genetic and mechanistic plasticity of isoprenoid biosynthesis across the domains of life, which provides a rich source of inspiration for metabolic engineers to effectively manipulate isoprenoid biosynthesis.
MEP pathway intermediates have non-isoprenogenic roles
MEcDP may serve as an oxidative stress signaling molecule
There is growing evidence that the MEP pathway intermediate MEcDP (Fig. 1) acts as an oxidative stress sensor because it accumulates under oxidative stress, acts as a stress signaling molecule in plants, and is remarkably stable due to its cyclized-diphosphate structure, which is characteristic of signaling molecules (12, 31, 74, 75). Interestingly, the pathogen L. monocytogenes and the nonpathogenic L. innocua encode genes for both the MEP and MEV pathways, although IspG and IspH are completely absent in L. innocua and IspG is defective in L. monocytogenes (76, 77). Importantly, the partial MEP pathway encoded by these species is functional and produces MEcDP (78), suggesting that this pathway might primarily serve to produce this molecule to signal oxidative stress. Furthermore, the oxygen-sensitive MEcDP synthase IspF was determined to be important to oxidative stress resistance in L. monocytogenes, and IspF, IspG, and IspH all have roles in murine infection despite this pathogen primarily using the MEV pathway for isoprenoid biosynthesis (76, 79, 80). It appears that the MEV pathway in Listeria supplies the IDP needed for isoprenoid production, while the MEP pathway serves as a metabolic stress response system, supporting a model in which isoprenoid biosynthesis pathway diversity enables environmental flexibility. In this model, the MEP pathway role is non-isoprenogenic in Listeria and is instead used to signal oxidative stress and provide an adaptive advantage.
HMBDP is recognized by immune cells
Another critical role mediated by an MEP intermediate is found in HMBDP (also referred to as HMBPP), which is the most potent and specific activator of Vγ9Vδ2 T cells, a specialized subset of γδ T cells in primates (30). While these non-canonical T cells can also respond to host-derived IDP, HMBDP is a 10,000-fold more potent activator (30), strongly suggesting that this sensing pathway represents a self-nonself detection mechanism. Importantly, Vγ9Vδ2 T cells expand significantly in response to infection by HMBDP-producing pathogens, including Mycobacterium tuberculosis, L. monocytogenes, and Plasmodium spp. (13). Once activated, Vγ9Vδ2 T cells mount a robust immune response characterized by the secretion of antimicrobial factors, polarization of αβ T cells, and establishment of immune memory (81). Thus, the Vγ9Vδ2 T cell response appears to be a robust and dedicated mechanism by which the host can detect bacteria via HMBDP binding and is a promising vaccine strategy against M. tuberculosis.
The Vγ9Vδ2 T cell response is protective in the context of infection but may also be involved in symbiotic bacterial-host homeostasis. It has been suggested in the literature that some pathogens evolved to use only the MEV pathway as an immune evasion strategy; however, the distribution of either pathway in successful pathogens and commensals alike suggests that host immunity was not the primary evolutionary pressure responsible for pathway diversification in bacteria (11). While MEV-utilizing bacteria do not stimulate a Vγ9Vδ2 T cell response, there are successful pathogens that necessarily use the MEP pathway, including M. tuberculosis. It is more likely that other factors, including virulence programs and intrinsic resistance, play a larger role in pathogenesis than the utilization of either pathway of isoprenoid biosynthesis. Rather, HMBDP sensing by Vγ9Vδ2 T cells may have evolved as a recent adaptation by primates to communicate with their bacterial symbionts, representing a unique mechanism of host detection of MEP-utilizing microbes.
ISOPRENOID PATHWAY DIVERSITY IN BACTERIA
Bacteria with an incomplete endogenous isoprenoid biosynthesis pathway
Although it is essential that cells have a strategy to obtain IDP, some bacteria have evolved to have an incomplete isoprenoid biosynthesis pathway or to lack one altogether. These auxotrophs must then acquire precursors or mature isoprenoids from prototrophs, resulting in an obligate symbiosis. For example, members of the class Mollicute are often symbionts of eukaryotic organisms and have reduced metabolic capabilities due to genome reduction. As Mollicutes streamline their genome to the bare essential genes for survival with their hosts, they often forego a primary isoprenoid biosynthesis strategy, as in the case of the human pathogen Mycoplasma genitalium, which possesses neither the MEP nor the MEV pathway (82, 83). Instead, M. genitalium’s isoprenoid acquisition strategy is to steal precursors or mature isoprenoids from its host.
Even as some bacteria have lost independent isoprenoid biosynthesis capabilities, their close relatives may retain a partial pathway, likely for non-isoprenogenic reasons. A systematic analysis of isoprenoid biosynthesis enzymes in the available repository of bacterial genomes revealed 72 other Mollicute species from the genera Mesomycoplasma, Mesoplasma, Metamycoplasma, Mycoplasma, Mycoplasmoides, and Mycoplasmopsis that lack homologs for isoprenoid biosynthesis genes (18). Interestingly, two notable exceptions emerged: Mycoplasma tullyi and Mycoplasmoides gallisepticum, in which all but the final MEP enzymes were identified despite having very small genomes at 0.867 and 0.964 Mb, respectively, and whose close relatives have totally dispensed with these genes (18, 84, 85). The presence of a near-complete MEP pathway in an ultra-reduced genomic landscape raises fascinating questions about the evolutionary trajectory of these bacteria and further suggests that the non-isoprenogenic functions of the MEP pathway may drive such diversification.
Endogenous isoprenoid biosynthesis pathways with varying degrees of completeness have been identified in yet more Mollicutes, underscoring the heterogeneity within this class. For example, most host-associated Spiroplasma possess complete MEP pathways, while one species lacks the critical enzyme Dxs and two species lack any MEP genes at all (18). Furthermore, most obligate intracellular Rickettsia and Orientia rely on host-generated isoprenoids (86, 87), although IspD was detected in several Rickettsia species (18). Importantly, an intracellular or otherwise host-associated lifestyle does not indicate an incomplete isoprenoid biosynthesis pathway, as demonstrated by Listeria monocytogenes, Shigella flexneri, and Francisella tularensis, which can reside in host cytosols but encode complete isoprenoid biosynthesis pathways. Additionally, genome size is not a definitive predictor of isoprenoid biosynthetic capability; while Francisella tularensis (1.9 Mb genome) possesses a complete isoprenoid biosynthesis pathway, Orientia tsutsugamushi (2.1 Mb genome) does not and acquires isoprenoids from its host.
While many obligate symbionts encode incomplete pathways or lack them altogether, the detection of isoprenoid biosynthesis genes that do not amount to a complete pathway in ultra-reduced genomes is indicative of extensive heterogeneity in isoprenoid acquisition strategies, which has been underappreciated. Furthermore, the incomplete MEP pathways detected in some species suggest non-isoprenogenic roles and suggest that broader evolutionary pressures may shape the course of pathway utilization in closely related species, commensurate with their particular niche.
Bacteria with dual isoprenoid biosynthesis strategies
In a starkly contrasting strategy, some bacteria possess complete copies of both the MEP and MEV pathways. Both pathways generate the same end products IDP and DMADP (Fig. 1), suggesting that dual pathway-encoding bacteria can utilize either for isoprenoid biosynthesis. However, each pathway may have specialized expression conditions, multifunctional intermediates, or roles in broader cellular processes.
The genus Streptomyces, and more broadly the Actinomycotota phylum, is well-known for producing a wide array of natural products, including antimicrobials, pharmaceuticals, and other economically relevant isoprenoids (88). Some Streptomyces possess genes for both the MEP and MEV pathways, which have each been demonstrated to be functional (89–92). Streptomyces genomes are rich in biosynthetic gene clusters (BGCs), genomic islands in which genes that catalyze the production of specialized metabolites are colocalized. Genes of the MEV pathway are sometimes found in such BGCs, and temporal expression of the MEV pathway has been observed to coincide with specialized natural product synthesis, suggesting that this pathway is being utilized in the production of the BGC end product (92, 93). Taken together, Streptomyces that possess both the MEP and MEV pathways appear to have distinct uses for each, despite generating identical end products. This functional partitioning allows the bacteria to tailor their metabolism during natural product synthesis (92).
Another case of dual pathway-encoding actinobacteria is found in the genus Mycobacteria, which contains the important human pathogen M. tuberculosis. While all Mycobacteria encode the MEP pathway, the Mycobacterium ulcerans-Mycobacterium marinum clade (MuMC) encodes both isoprenoid biosynthesis pathways (94, 95). In M. marinum, the MEP pathway has been determined to be essential, while the non-essential MEV pathway supports metabolic flexibility during stress (96). Interestingly, while the MEV pathway appears to be intact in M. marinum, the M. ulcerans pathway is likely nonfunctional due to a disruption of hmgS (95). The presence of a functional MEV pathway in M. marinum and loss of that functionality in the closely related M. ulcerans further underscores that each pathway is distinct and subject to selective pressure, although they both produce IDP. It is likely that the selective pressures are driven by the non-isoprenogenic differences of the MEP and MEV pathways.
In contrast to Mycobacteria, the genus Listeria primarily uses MEV with some species possessing an incomplete, likely non-isoprenogenic MEP pathway (11, 77, 97). As discussed in “The MEP and MEV pathways for isoprenoid biosynthesis,” above, L. monocytogenes and L. innocua have incomplete MEP pathways that terminate at the intermediate MEcDP, which has been implicated in oxidative stress signaling (12, 77). The termination of this partial pathway at MEcDP synthesis, in contrast to primary IDP synthesis via the MEV pathway, suggests that this could be the root of the evolutionary pressure for Listeria to retain an incomplete MEP pathway. Taken together, encoding both pathways partially or in full enables bacteria to fine-tune isoprenoid synthesis under specific conditions or even to generate intermediates that might protect them from environmental stress.
Intra-genus diversity in isoprenoid acquisition strategies
When bacterial phylogeny is overlaid with isoprenoid pathway usage data, a striking pattern of heterogeneity emerges where phyla and isoprenoid acquisition strategy do not obviously align (Fig. 2A). We performed genus-level pathway analysis and confirmed that the MEP pathway is the predominant bacterial isoprenoid biosynthesis pathway, although there are lineages that use the MEV pathway. Interestingly, the phyla Actinomycetota and Bacillota contain considerable diversity compared to the other represented phyla (Fig. 2A). Isoprenoid biosynthesis gene presence-or-absence analysis was also performed for 4,249 bacterial species representing 1,412 genera (Fig. 2B; Table S1). We refer the reader to this supplementary resource for species-level information regarding which isoprenoid genes were identified. In summary, for the species analyzed, 82.94% use MEP, 12.03% use MEV, 2.42% have both pathways, and 2.61% have neither (Table S1).
Isoprenoid biosynthesis genes may be acquired in a lineage through a variety of mechanisms, including vertical inheritance from a common ancestor, horizontal transfer from another species, or by some combination of both. The distribution of pathways and punctate nodes of heterogeneity observed in Fig. 2A suggests multiple routes of diversification for these pathways. MEP genes are not found in operons and are extensively distributed across all phyla (Fig. 2A), making it unlikely that their distribution is a result of HGT (98). Conversely, MEV genes are more frequently found in operons, suggesting that some occasions of pathway diversity result from HGT.
Within individual genera, patterns emerge that suggest potential pathway decay. Of the 110 Wolbachia species analyzed, most encode MEP genes, while less than 2% have neither pathway (Table S1), potentially suggesting pathway loss from the genus ancestor. Around 8% of Streptococcus species and approximately half of Enterococcus species analyzed have an incomplete MEP pathway in addition to a complete MEV pathway (Table S1), potentially representing another example of a partial MEP pathway acting as an oxidative stress sensor as discussed in “The MEP and MEV pathways for isoprenoid biosynthesis,” above. Furthermore, while Legionella have been characterized as encoding the MEV pathway, all Legionella species analyzed lack the phosphomevalonate kinase PmvK, and most species additionally lack the hydroxymethylglutaryl-CoA synthase HmgS (99). Previous assignments may have relied on single gene presence-or-absence data; however, because these Legionella lack one or more critical enzymes in this pathway, they did not meet our threshold to be assigned as encoding the MEV pathway. Further work is needed to address whether alternative enzymes exist to compensate for the lack of canonical mevalonate enzymes or how Legionella otherwise acquires isoprenoid precursors in their absence. Taken together, these examples present fascinating questions as to whether these are cases of specific gene loss to accumulate a particular intermediate or whether these lineages are shedding an entire pathway.
Alternatively, in some clades, the pattern of distribution suggests pathway acquisition. While most members of the phylum Bacteriodota encode MEP, a considerable subset of related genera instead encodes only the MEV pathway, suggesting that a common ancestor of this group may have acquired MEV or potentially even retained an ancestral MEV pathway (Fig. 2A) (58). This gene acquisition is also observed in Burkholderia and Clostridium, which are largely MEP-encoding but have very rare instances of species with genes for both pathways (~4% of each genus have both, Table S1). When genera are analyzed for diversity in isoprenoid acquisition strategies, over half of the instances of intra-genus diversity were due to species encoding multiple genes for both pathways (25/44 genera, Table S1).
Finally, some genera have such varied pathway usage that it becomes difficult to speculate on an ancestral pathway route. In the case of the order Rickettsiales, there are indications of both vertical inheritance and horizontal gene transfer. Some genera within Rickettsiales possess a complete MEP pathway while others encode only downstream genes to process imported IDP, suggesting vertical inheritance and pathway degradation (86). Meanwhile, Rickettsiales that lack a complete MEP pathway often have Idi, which is hypothesized to have been acquired horizontally from non-alphaproteobacterial origins (87). Furthermore, nearly the full breadth of pathway diversity observed in the bacterial kingdom can be found in the genus Brachybacterium. The majority of the species analyzed encode genes for the MEP pathway, while the closely related Bacillus halotolerans and Brachybacterium kimchii encode genes only for the MEV pathway, and Bordetella avium encodes genes for both pathways (Fig. 2C). Brachybacterium species have been isolated from a wide range of niches, including associations with animals, soil, fresh and seawater, dairy, seafood, and, rarely, as pathogens (100). The wide variety of isoprenoid biosynthesis strategies detected in this genus may reflect the broad niche range of its species and suggest that yet-undetermined evolutionary pressures may be shaping pathway utilization across niches. Further work is needed to understand the evolutionary pressures that influence pathway suitability for a given niche and, in particular, why Brachybacterium species possess a range of isoprenoid biosynthesis strategies.
It is important to note that these analyses examine isoprenoid biosynthesis at the genome level, that is, these suggested routes of diversification are based on homologous gene presence-or-absence data and require experimental validation. For example, several Streptomyces species analyzed encode HmgR from the MEV pathway in addition to a complete MEP pathway, necessitating further study to determine whether a single gene from the other pathway contributes to isoprenoid biosynthesis (Table S1). Nonetheless, by taking a comprehensive view of isoprenoid biosynthesis through the lens of evolution, it becomes clear how evolution acts not only at a species level but also at the level of individual genes and metabolites. The MEP and MEV pathways both fulfill a requirement for cells to generate essential isoprenoid precursors, but each uses unique mechanisms and energetics, which may influence which pathway is selected for in a given niche. Although all bacteria must possess either or both of these pathways—or steal from a host—there is great flexibility for species to utilize the pathway best suited for their niche.
SYNTHETIC BIOLOGY APPROACHES TO ENGINEER ISOPRENOID BIOSYNTHESIS
Economically relevant isoprenoids are critical to many industries, including agriculture, pharmaceuticals, and fuels, and are largely generated via unsustainable practices (101, 102). High demand and current unsustainable production methods have motivated synthetic biology efforts toward a “microbial cell factory” approach to replace or supplement current methods of isoprenoid synthesis (103–105). For example, the need for large-scale petroleum processing can be reduced or eliminated by engineering microbes to produce isoprene and other “platform chemicals,” which serve as building blocks for high-value products (102, 106, 107). Drugs like the antimalarial artemisinin and the anticancer paclitaxel, which were originally isolated from slow-growing plants, can be produced in microbes at much higher yields (108, 109). Additional efforts are underway to develop novel, non-natural isoprenoids with unique properties (110). Altogether, there have been significant and successful efforts to engineer microbes to produce platform chemicals, pharmaceuticals, fuels, and other valuable isoprenoids (106). A deeper understanding of microbial isoprenoid biosynthesis and its regulation will continue to be instrumental in the development of sustainable isoprenoid production technologies.
Through many years of research, the field has moved toward a clearer understanding of effective strategies to engineer microbes for isoprenoid production. Generally, engineering has focused on optimizing the expression of key enzymes for improved flux, ensuring sufficient precursor supply, and blocking competing pathways (111). Some of the highest isoprenoid yields of target molecules are from engineering the MEV pathway in microbes and yeast (104, 112–116). Generally, HmgR, Mvk, and Idi represent the critical enzymatic steps targeted in MEV engineering approaches, although efforts must be tailored to each host organism and target molecule (113, 114, 117, 118). In contrast, engineering approaches addressing the MEP pathway have generally produced lower product yields than those of the MEV pathway, despite the greater theoretical yield of the MEP pathway (54). A critical barrier to successful MEP pathway engineering is an incomplete understanding of how to circumvent the tight regulation of this pathway, which includes feedback inhibition at the key regulatory step mediated by Dxs (119–121). Another challenge in MEP pathway engineering lies in the enzymes IspG and IspH, which must be loaded with iron-sulfur clusters, supplied with reducing equivalents to function, and are notoriously oxygen sensitive (122).
One strategy to increase terpenoid yield is to express both isoprenoid biosynthetic pathways in one organism, although such heterologous expression is limited by imbalanced reducing equivalents, unidentified regulatory networks, and distinct cofactors and energetic requirements. Exogenous MEP expression in an organism that natively uses the MEV pathway is possible, although it remains limited by yield and requires the support of multiple partner enzymes (9, 123–125). For example, MEP expression in yeast has not produced a higher yield of target terpenoid products despite the MEP pathway having a higher theoretical productivity (126). Conversely, exogenous MEV expression in organisms natively using MEP has led to enhanced isoprenoid yields, with some groups demonstrating synergy between the two pathways (14, 104, 127). This improved yield upon expression of both pathways may be due to the generation of reducing power by the MEV pathway (Fig. 1; Table 1) or by the relief of feedback inhibition due to increased overall flux.
There is much to be learned from the dual-isoprenoid biosynthesis pathway-bearing organisms that can help metabolic engineers unlock greater product yields. For example, there is evidence for metabolic compensation in a MEV mutant of M. marinum, which encodes both pathways, although interestingly, MEV pathway overexpression did not compensate for a loss of MEP (96). Further study of such interactions is critical to reveal synergistic properties or non-isoprenogenic roles of the MEP or MEV pathways in these species, advancing the development of engineered microbes for sustainable isoprenoid production. By understanding the extensive heterogeneity generated through natural bacterial evolution, engineering approaches can skip the arduous process of developing and optimizing a novel biosynthetic route, instead utilizing the existing strategies in nature.
CONCLUDING REMARKS
In this review, we argue that the extensive diversity observed at the levels of enzyme isoforms, pathway usage, and pathway completeness necessitates a more nuanced view of evolutionary inheritance of isoprenoid biosynthesis, as well as an appreciation of non-isoprenogenic effects of these pathways.
Each pathway has unique energetic features that may influence its acquisition or retention. The MEP pathway is more carbon-efficient, while the MEV pathway is more energy-efficient, suggesting that some bacteria encode different pathways to meet different energetic needs dictated by varied host niches and environmental challenges. While we cannot at this time define the exact evolutionary pressures influencing whether a bacterial species will encode one pathway, both, or neither, it seems likely that IDP synthesis strategies diversified among bacteria via a variety of mechanisms.
Finally, the heterogeneity observed across bacteria presents a fascinating perspective of the “natural engineering” achieved through evolution and offers insights to rationally design and optimize isoprenoid metabolism to tackle real-world sustainability issues. The extensive heterogeneity of pathway usage, enzyme forms, and end products that exist in nature represents an abundant source of natural diversity that can be harnessed to achieve such engineering goals.
In this review, we have examined the distribution of pathways across bacteria and highlighted previously underappreciated intra-genus diversity, likely owing to a variety of factors including environmental and host niches, as well as yet-undetermined evolutionary pressures. Analysis of natural instances of isoprenoid biosynthesis diversity, especially of those bacteria with incomplete or both pathways, provides an opportunity to further understand what roles the MEP and MEV pathways have beyond IDP generation. These roles may include oxidative stress signaling or other off-target effects of intermediate metabolites yet to be discovered. These non-isoprenogenic roles may be part of the evolutionary pressures that affect species’ acquisition or retention of one path over the other. The continued study of isoprenoid heterogeneity among bacteria will be instrumental in the development of microbes for sustainable isoprenoid production.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Faylo JL, Ronnebaum TA, Christianson DW. 2021. Assembly-line catalysis in bifunctional terpene synthases. Acc Chem Res 54:3780–3791. doi:10.1021/acs.accounts.1c 0029634254507 PMC 8530900 · doi ↗ · pubmed ↗
- 2Holstein SA, Hohl RJ. 2004. Isoprenoids: remarkable diversity of form and function. Lipids 39:293–309. doi:10.1007/s 11745-004-1233-315357017 · doi ↗ · pubmed ↗
- 3Lange BM, Rujan T, Martin W, Croteau R. 2000. Isoprenoid biosynthesis: the evolution of two ancient and distinct pathways across genomes. Proc Natl Acad Sci USA 97:13172–13177. doi:10.1073/pnas.24045479711078528 PMC 27197 · doi ↗ · pubmed ↗
- 4Avalos M, Garbeva P, Vader L, van Wezel GP, Dickschat JS, Ulanova D. 2022. Biosynthesis, evolution and ecology of microbial terpenoids. Nat Prod Rep 39:249–272. doi:10.1039/D 1NP 00047 K 34612321 · doi ↗ · pubmed ↗
- 5Goldstein JL, Brown MS. 1990. Regulation of the mevalonate pathway. Nature 343:425–430. doi:10.1038/343425 a 01967820 · doi ↗ · pubmed ↗
- 6Park J, Matralis AN, Berghuis AM, Tsantrizos YS. 2014. Human isoprenoid synthase enzymes as therapeutic targets. Front Chem 2:50. doi:10.3389/fchem.2014.0005025101260 PMC 4106277 · doi ↗ · pubmed ↗
- 7Lombard J, López-García P, Moreira D. 2012. Phylogenomic investigation of phospholipid synthesis in archaea. Archaea 2012:630910. doi:10.1155/2012/63091023304072 PMC 3533463 · doi ↗ · pubmed ↗
- 8González-Hernández RA, Valdez-Cruz NA, Macías-Rubalcava ML, Trujillo-Roldán MA. 2023. Overview of fungal terpene synthases and their regulation. World J Microbiol Biotechnol 39:194. doi:10.1007/s 11274-023-03635-y 37169980 PMC 10175467 · doi ↗ · pubmed ↗