Engineering PRRs: A new path to durable resistance
Dongjiao Wang, Zhankun Li, Wentao Yang, Youxiong Que, Ming Chang

TL;DR
Scientists engineered a plant receptor to boost immunity against diseases without affecting plant growth.
Contribution
A reprogrammed immune receptor provides broad-spectrum disease resistance across species.
Findings
Structural optimization of RLP23 enables functional signaling triggered by nlp20.
The engineered receptor confers broad-spectrum disease resistance without growth compromise.
Rationally reprogrammed receptors offer a durable strategy for crop protection.
Abstract
Engineering the Arabidopsis cell-surface immune receptor RECEPTOR-LIKE PROTEIN 23 (RLP23) enhances plant immunity by enabling functional signaling triggered by the conserved pathogen peptide nlp20. Structural optimization enables broad-spectrum disease resistance across species without compromising growth, demonstrating the potential of rationally reprogrammed pattern recognition receptors as a durable and sustainable strategy for crop protection.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
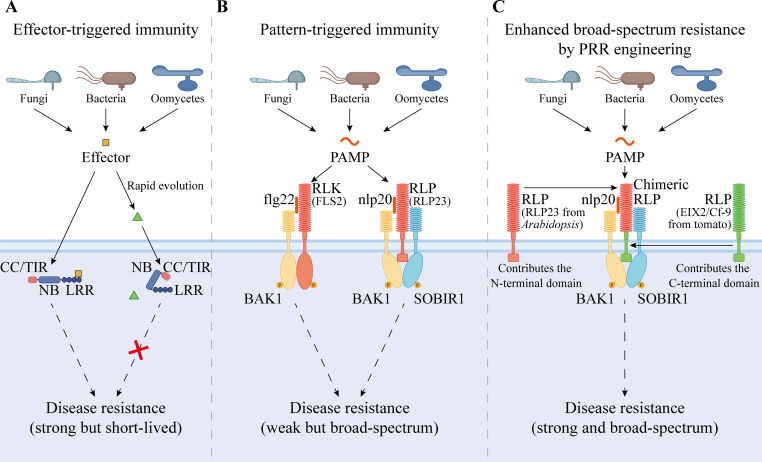 Figure 1
Figure 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsViral Infectious Diseases and Gene Expression in Insects · RNA Interference and Gene Delivery · Antimicrobial Peptides and Activities
Rethinking resistance breeding through cell-surface immune receptors
1
Traditional crop disease-resistance breeding has mainly focused on nucleotide-binding leucine-rich repeat (NB-LRR, NLR) receptors, which detect pathogen effectors and promote effector-triggered immunity (ETI). This strategy provides strong protection, however, pathogen effectors rapidly mutate, causing frequent resistance breakdowns and making NLR-based resistance short-lived in the field (Fig. 1A) [1].Fig. 1. From effector-triggered immunity (ETI) to engineered pattern recognition receptors (PRRs) for durable and broad-spectrum disease resistance. A Intracellular nucleotide-binding leucine-rich repeat (NB-LRR, NLR) receptors, typically composed of an N-terminal coiled-coil (CC) or Toll/interleukin-1 receptor (TIR) domain, recognize pathogen effectors to activate ETI. Although ETI confers strong and specific resistance, it is often short-lived because pathogen effectors rapidly evolve to evade recognition. B Cell-surface PRRs, including receptor-like kinases (RLKs) and receptor-like proteins (RLPs), detect conserved pathogen-associated molecular patterns (PAMPs) and activate pattern-triggered immunity (PTI). For example, the RLK protein FLAGELLIN-SENSING 2 (FLS2) recognizes bacterial flg22 and signals through the co-receptor BRASSINOSTEROID INSENSITIVE 1-ASSOCIATED KINASE 1 (BAK1), while the RLP23 detects the conserved peptide nlp20 derived from necrosis- and ethylene-inducing peptide-like proteins (NLPs) and signals via the co-receptors BAK1 and SUPPRESSOR OF BIR1-1 (SOBIR1). PTI responses are generally weaker than ETI but provide broad and durable protection. C Cross-species engineering of PRRs enables enhanced and stable immune signaling. The Arabidopsis RLP23, which recognizes the conserved peptide nlp20 from diverse fungi, oomycetes, and bacteria, was optimized by swapping its C-terminal region with that from the tomato RLP ETHYLENE-INDUCING XYLANASE RECEPTOR 2 (EIX2) or CLADOSPORIUM FULVUM RESISTANCE PROTEIN 9 (Cf-9). These chimeric receptors achieved stronger PTI activation without fitness cost. This schematic illustrates the general concept of PRR engineering, in which the C-terminal domain from one species (Plant A) and the N-terminal domain from another (Plant B) can be combined to produce receptors with improved signal transduction and broad-spectrum disease resistance. Solid lines indicate direct relationships; dashed lines indicate indirect links.Fig. 1
Attention has shifted to cell-surface immune receptors, known as pattern recognition receptors (PRRs), including two main classes: receptor-like kinases (RLKs) and receptor-like proteins (RLPs) [2]. Located at the plasma membrane, RLKs and RLPs recognize conserved pathogen- or damage-associated molecular patterns (PAMPs/DAMPs) and activate pattern-triggered immunity (PTI) [2]. PTI responses are generally milder than ETI but tend to be broader and more durable, making PRRs attractive targets for engineering long-term, wide-spectrum resistance (Fig. 1B).
Interestingly, cross-species transfer of PRRs often results in limited functionality since receptor and co-receptor interactions and downstream signaling components such as SUPPRESSOR OF BIR1-1 (SOBIR1) and BRASSINOSTEROID INSENSITIVE 1-ASSOCIATED KINASE 1 (BAK1) differ among plant species [3]. The recent study by Yang et al. overturns this trend by demonstrating that the Arabidopsis RLP23 protein can be engineered and transferred into diverse crops, where functional immune signaling is restored or amplified through C-terminal domain replacement, enhancing resistance without penalty to growth [4]. This signals a shift from purely NLR-based breeding to a network-based design of immune receptors.
Structure-guided reengineering of RLP23 enhances immune signaling
2
The Arabidopsis RLP23 protein recognizes nlp20, a highly conserved 20-amino acid peptide derived from necrosis- and ethylene-inducing peptide-like proteins (NLPs) that are widely present in fungi, oomycetes, and bacteria [5]. Perception of nlp20 triggers typical PTI responses such as accumulation of ethylene and reactive oxygen species (ROS) and activation of defense-related genes, providing a broad and stable layer of resistance (Fig. 1B) [5].
Structural analysis revealed that the C-terminal region of RLP23, consisting of juxtamembrane (JM), transmembrane (TM), and intracellular (IC) domains, is essential for signal transduction mediated by the co-receptors SOBIR1 and BAK1 [4]. In Nicotiana benthamiana, deletion of the IC domain weakened nlp20-induced ethylene responses, whereas deletion of the full C-terminal region caused an even stronger reduction, indicating that C-terminal domains are essential for receptor activation in heterologous systems [4].
Replacing the IC or C-terminal domains of RLP23 with those from tomato RLPs ETHYLENE-INDUCING XYLANASE RECEPTOR 2 (EIX2) and CLADOSPORIUM FULVUM RESISTANCE PROTEIN 9 (Cf-9) greatly enhanced immune signaling in N. benthamiana without changing nlp20 recognition [4]. Domain swaps using RLPs from tomato, rice, or poplar further amplified responses, suggesting that species-specific C-terminal features optimize the compatibility between receptors and co-receptors [4]. These findings establish the C-terminal region of RLP23 as a tunable module controlling signaling output and enable functional cross-species transfer of PRRs through downstream signaling adaptation (Fig. 1C).
Engineered RLP23 confers broad resistance without fitness cost
3
Introducing the engineered RLP23 variants into Arabidopsis and N. benthamiana markedly enhanced resistance to multiple necrotrophic and hemibiotrophic pathogens producing NLP effectors [4]. Plants expressing domain-swapped receptors exhibited stronger ethylene accumulation, elevated ROS bursts, and up-regulation of defense marker genes, demonstrating robust PTI [4]. Importantly, these modifications did not impose observable penalties on growth or reproduction, indicating that receptor activation remained precisely regulated [4].
The absence of fitness costs is a crucial achievement. Many constitutively active or overexpressed immune receptors cause dwarfism or yield loss due to chronic defense activation [6]. In contrast, the reengineered RLP23 variants retained low basal activity while mounting rapid and strong responses only upon ligand recognition, highlighting the advantage of signaling-competent but conditionally activated receptor design [4].
Moreover, the engineered receptors could maintain compatibility with endogenous co-receptors SOBIR1 and BAK1, ensuring efficient signal transduction [4]. The combination of enhanced responsiveness, functional stability, and growth neutrality highlights the feasibility of designing precision-tuned PRRs for sustainable disease resistance across crops (Fig. 1C).
Paving the way for sustainable and resilient crop immunity
4
The reengineering of RLP23 exemplifies a rational approach to strengthening PTI through structural optimization of PRRs. This work is part of a growing movement toward precision immune design, which aims to improve disease resistance while maintaining plant vigor and yield.
A compelling precedent is translational regulation of defense genes using upstream open reading frames (uORFs). Traditional transcriptional overexpression of immune regulators, such as the salicylic acid receptor NONEXPRESSER OF PATHOGENESIS-RELATED GENES 1 (NPR1) in Arabidopsis, often leads to constitutive defense and growth penalties [7]. A TL1-BINDING FACTOR 1 (TBF1)-derived uORF cassette that allows NPR1 translation only upon pathogen attack, conferring robust resistance in rice without yield loss [8]. This innovation demonstrates how post-transcriptional control can reconcile immunity with productivity, providing a regulatory “buffer” for engineered defense activation.
At the intracellular immune receptor level, interfamily co-transfer of NLR pairs offers another paradigm. Transfer of the pepper Bs2 sensor and its helper NLR REQUIRED FOR CELL DEATH (NRC) into rice reconstitutes effector recognition against Xanthomonas oryzae pv. oryzicola, generating resistant lines with unaltered field performance [9]. The success of this cross-family receptor reconstruction reveals the feasibility of transplanting complete immune modules across taxonomic boundaries.
More recently, researchers systematically mined over 13,000 LRR-RLKs to identify SELECTIVE COLD SHOCK PROTEIN RECEPTOR (SCORE), a PRR detecting bacterial cold-shock protein peptides [10]. Guided by structural and phylogenomic analyses, synthetic SCORE variants were engineered to recognize diverse pathogens, including Ralstonia, Xanthomonas, and Candidatus Liberibacter asiaticus, demonstrating a scalable framework for receptor discovery and functional reprogramming [10].
Together, these advances mark a turning point from gene-by-gene improvement to network-level engineering of plant immunity. By integrating structure-based PRR reconfiguration, translational fine-tuning, and modular receptor transfer, future breeding can produce ideal crops with durable, broad-spectrum, and fitness-neutral resistance, thereby guiding a new era of resilient agriculture.
CRediT authorship contribution statement
Dongjiao Wang: Writing – original draft. Zhankun Li: Writing – original draft. Wentao Yang: Writing – original draft. Youxiong Que: Writing – review & editing. Ming Chang: Writing – review & editing.
Declaration of competing interest
The authors declare no competing interests.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Wu C.H.Derevnina L.The battle within: how pathogen effectors suppress NLR-mediated immunity Curr Opin Plant Biol 74202310239610.1016/j.pbi.2023.10239637295294 · doi ↗ · pubmed ↗
- 2Albert I.Hua C.L.Nurnberger T.Pruitt R.N.Zhang L.S.Surface sensor systems in plant immunity Plant Physiol 18242020158215963182250610.1104/pp.19.01299 PMC 7140916 · doi ↗ · pubmed ↗
- 3Huang W.R.H.Joosten M.H.A.J.Immune signaling: receptor-like proteins make the difference Trends Plant Sci 301202554683859415310.1016/j.tplants.2024.03.012 · doi ↗ · pubmed ↗
- 4Yang Y.K.Steidele C.E.Huang X.L.Löffelhardt B.Huang L.L.Jones J.D.G.Engineered pattern recognition receptors enhance broad-spectrum plant resistance Nat Biotechnol 202510.1038/s 41587-025-02858-841083902 · doi ↗ · pubmed ↗
- 5Albert I.Böhm H.Albert M.Feiler C.E.Imkampe J.Wallmeroth N.An RLP 23-SOBIR 1-BAK 1 complex mediates NLP-triggered immunity Nat Plants 11020151514010.1038/nplants.2015.14027251392 · doi ↗ · pubmed ↗
- 6Freh M.Gao J.L.Petersen M.Panstruga R.Plant autoimmunity-fresh insights into an old phenomenon Plant Physiol 18832022141914343495837110.1093/plphys/kiab 590PMC 8896616 · doi ↗ · pubmed ↗
- 7Fitzgerald H.A.Chern M.S.Navarre R.Ronald P.C.Overexpression of (At)NPR 1 in rice leads to a BTH- and environment-induced lesion-mimic/cell death phenotype Mol Plant Microbe Interact 17220041401511496452810.1094/MPMI.2004.17.2.140 · doi ↗ · pubmed ↗
- 8Xu G.Y.Uan M.Y.Ai C.R.Liu L.J.Zhuang E.Karapetyan S.u ORF-mediated translation allows engineered plant disease resistance without fitness costs Nature 545765520174914942851444810.1038/nature 22372 PMC 5532539 · doi ↗ · pubmed ↗