glorund functions in the Drosophila intestine to regulate triglyceride storage and the expression of lipid transport protein genes
Roman Voskoboynikov, Justin R. DiAngelo

TL;DR
This study shows that the Drosophila gene glorund helps regulate fat storage and lipid transport in the intestine.
Contribution
The novel finding is that glo functions in the intestine to control lipid transport and fat storage.
Findings
Reducing glo in intestines increases whole animal triglyceride storage.
Lower glo levels decrease expression of lipid transport protein genes.
Glo regulates lipid transport and organismal fat storage in Drosophila.
Abstract
The intestine acts as the primary site for absorption of dietary lipids. These lipids are packaged and transported via lipoprotein complexes, whose altered levels correlate with metabolic disease. The Drosophila splicing factor glorund (glo) has been shown to affect the expression of apoB-family lipoproteins, including microsomal triacylglycerol transfer protein, lipid transfer particle, and lipophorin, in the fly adipose tissue. Here, we demonstrate that decreasing glo in intestines leads to increased whole animal triglyceride storage, but decreased expression of lipid transport protein genes. Together, these data suggest that glo functions in the intestine to regulate lipid transport and organismal fat storage.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
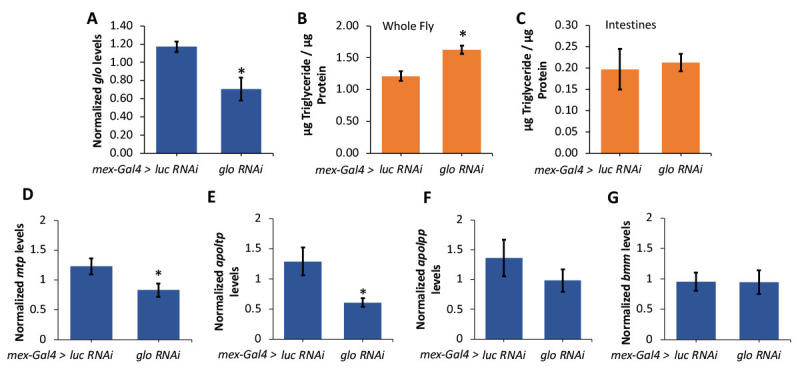 Figure 1
Figure 1|
|
|
|
|
|
GCTGGGCTTCAACAATCTGC |
AATTTCCACCGTTGTTGCCG |
|
(
|
GACGCTTCAAGGGACAGTATCTG |
AAACGCGGTTCTGCATGAG |
|
(
|
GTTCGAGGTGAGTGGTTGGT |
AGCTGCGTCTCATTGGAGAT |
|
(
|
ATCGGCTCAACACAAAAACC |
AGGCAAAAGCGATCTCAAAA |
|
(
|
GTGGGAAGCTTCGTGAAGAG |
AAAACGCGATACCATTCGAG |
|
(
|
ACGTGATCATCTCGGAGTTTG |
ATGGTGTTCTCGTCCAGAATG |
|
|
|
|
|
UAS-luc RNAi |
|
BL#35788 |
|
UAS-glo RNAi |
|
BL#33668 |
|
Mex-Gal4 |
|
BL#91368 |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsInvertebrate Immune Response Mechanisms · Neurobiology and Insect Physiology Research · Lipid metabolism and biosynthesis
Description
After a meal, dietary triglycerides (TAGs) are incorporated into lipoprotein molecules called chylomicrons in the intestine and then carried to muscle and adipose tissue, where they are hydrolyzed releasing free fatty acids, which are subsequently esterified into TAGs for storage in adipose tissue, or metabolized to produce energy in muscle (Mahmood, 2014). The microsomal TAG transport protein (MTP), which is expressed in the liver and intestinal mucosa, catalyzes the transfer of dietary TAGs and cholesteryl esters into chylomicrons and very low-density lipoproteins (Mahmood, 2014). MTP deficiency in humans can lead to a rare disease called Abetalipoproteinemia and deleting mtp in the mouse intestine results in decreased chylomicron assembly and secretion (Xie et al, 2006). However, how the mtp gene and its function are regulated is not fully understood.
Recent work from our lab and others in both humans and the genetic model system Drosophila has identified mRNA splicing factors as regulating lipid metabolism (Gingras et al, 2014, Pihlajamäki et al, 2011). Our lab has also shown that the Drosophila splicing factor glorund ( glo ) promotes lipid transport by stimulating the expression of mtp and other apoB-containing lipoprotein genes, apolipophorin ( apolpp ) and apolipoprotein lipid transfer particle ( apoltp ) (Kolasa et al, 2021). Lipids and sterols are added to the lipophorins, the major lipid carrier in flies, by LTP proteins and then secretion of the lipidated lipophorins occurs with the help of MTP molecules (Palm et al, 2012). Consistent with a role of glo in regulating lipid transport, decreased expression of glo in *Drosophila * adipose tissue results in increased TAGs stored in fat cells and less TAGs stored in non-fat body tissues (Kolasa et al, 2021). However, the function of glo in the Drosophila intestine to regulate lipid transport and metabolism is not yet known.
To study the function of *glorund * in the intestine to regulate lipid metabolism, glo expression was decreased in Drosophila intestines by inducing RNAi using the Gal4-UAS system ( Fig. 1A ). To determine whether glo function in the intestine is necessary for triglyceride storage in the entire animal, we measured triglycerides in the whole fly after intestine-specific glo knockdown ( Fig. 1B ). Total fly body triglycerides were increased in glo-RNAi flies suggesting glorund acts in the fly intestine to regulate overall lipid storage. To investigate whether decreasing *glorund * altered intestinal lipid storage autonomously, intestines were dissected from glo-RNAi flies and triglycerides were analyzed ( Fig. 1C ). Interestingly, triglyceride storage was not altered in the glo-RNAi intestines. However, triglycerides still accumulate throughout the entire fly after glo knockdown, which suggests that glorund may act in the *Drosophila * intestine to regulate lipid transport.
To determine if the decrease of glo is affecting the lipid transport machinery, RNA was isolated from glo-RNAi intestines and qPCR was performed to detect the levels of several key lipoprotein genes involved in lipid transport ( mtp , apoltp , and apolpp ), and the triglyceride lipase bmm . Interestingly, mtp and apoltp were both significantly lower in intestines with decreased *glo * levels ( Fig. 1D- 1E), while *apolpp * and *bmm * expression was not changed ( Fig. 1F- 1G). Together, these data suggest that glorund may function in the fly intestine to regulate the storage and transport of lipids.
In this study, we have shown that the splicing factor, glorund, plays a role in the *Drosophila * intestine to regulate lipid storage, perhaps through controlling lipid transport as the expression of the apoB-containing genes *mtp * and *apoltp * are decreased in *glo-RNAi * flies. Previous studies decreasing *mtp * levels specifically in the intestine in mice has resulted in decreased lipid absorption and chylomicron secretion leading to triglyceride buildup in the intestine (Xie et al, 2006, Iqbal et al, 2010, Iqbal et al, 2014). In addition, intestine-specific *mtp * knockout mice had increased hepatic lipogenesis, perhaps compensating for the decreased lipid absorption and transport from the intestine (Xie et al, 2006).
While we did not observe triglyceride accumulation in *glo-RNAi * intestines, it is possible that lipid transport could be decreased in these animals and measuring apoB-containing protein secretion from the intestine as well as lipid profiles in the hemolymph of *glo-RNAi * flies may help to further characterize any lipid transport defects that may exist in these flies. Moreover, the increased triglycerides in *glo-RNAi * animals could be due to increased lipogenesis in fat body tissue like what was seen in livers of intestine-specific *mtp * knockout mice (Xie et al, 2006); measuring lipogenesis in fat tissue in *glo-RNAi * flies would help test this hypothesis. In addition, it is also possible that additional genes to those shown here are regulated by glorund to control lipid storage; further experimentation measuring differential gene expression in flies with decreased *glo * expression would help characterize how the glorund protein regulates lipid storage and metabolism in the fly intestine.
Methods
Fly husbandry: Flies were cultured on standard yeast-sugar-cornmeal medium (9 g *Drosophila * agar (Genesee Scientific), 100 mL Karo Lite Corn Syrup, 65 g cornmeal, 40 g sucrose, and 25 g whole yeast in 1.25 L water) and grown in an incubator at 25°C with a 12h:12h light:dark cycle. UAS-lucRNAi and UAS-gloRNAi males were crossed to mex-Gal4 virgin females and approximately 1 week old, mated female progeny were used in the experiments described here.
Triglyceride and protein assays: Whole one-week old female fly pairs, or groups of 10 intestines dissected from one-week old females were suspended in lysis buffer (140 mM NaCl, 50 mM Tris-HCl, pH 7.4, 0.1% Triton-X, and 1X protease inhibitor (Roche)), and then homogenized via sonication. Protein concentrations were measured using the Pierce BCA Assay kit (Thermo Fisher Scientific) and triglyceride concentrations were measured using the Infinity Triglyceride Reagent (Thermo Fisher Scientific) according to manufacturer’s instructions. Triglyceride concentrations were normalized to protein levels for each sample.
RNA isolation: Groups of 25-30 intestines dissected from one-week old female flies were suspended in TRIzol Reagent (Thermo Fisher Scientific) and homogenized briefly followed by a 5 min incubation at room temperature. Samples were chloroform extracted and nucleic acids were precipitated by adding isopropanol. Pellets were washed with 70% ethanol, air dried for 5 minutes, and then resuspended in nuclease-free water.
DNase treatment and cDNA synthesis: 5 µg of each RNA sample was DNase treated with the DNA-Free Kit (Ambion), according to manufacturer’s instructions. 0.25 µg of DNased RNA samples were reverse transcribed using qScript Ultra cDNA Supermix (QuantaBio), according to manufacturer’s instructions.
qPCR: qPCR reactions were made from 1µl of cDNA, 2x Perfecta SYBR Green (QuantaBio) and 200nM of the forward and reverse primer for each gene segment. The qPCR cycling conditions were as follows: 3 min at 95 °C; 40 cycles of: 30 s at 95 °C, 1 min at 60 °C, and 30 s at 72 °C, with a melt curve. The following genes were amplified as described in Kolasa et al (2021): glo, rp49 , apoltp , mtp , apolpp , and bmm . Resulting values for each gene were normalized to rp49 . The primer sequences are shown below.
**: **
Reagents
**: **
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Cartwright IJ PlonnéD Higgins JA 2000111 Intracellular events in the assembly of chylomicrons in rabbit enterocytes.J Lipid Res 41110022-22751728173911060342 · pubmed ↗
- 2Gingras RM Warren ME Nagengast AA Diangelo JR 20131211 The control of lipid metabolism by m RNA splicing in Drosophila.Biochem Biophys Res Commun 44320006-291X 67267610.1016/j.bbrc.2013.12.02724333419 PMC 4412844 · doi ↗ · pubmed ↗
- 3Hussain MM 201461 Intestinal lipid absorption and lipoprotein formation.Curr Opin Lipidol 2530957-967220020610.1097/MOL.000000000000008424751933 PMC 4265799 · doi ↗ · pubmed ↗
- 4Iqbal J Li X Chang BH Chan L Schwartz GJ Chua SC Jr Hussain MM 2010217 An intrinsic gut leptin-melanocortin pathway modulates intestinal microsomal triglyceride transfer protein and lipid absorption.J Lipid Res 5170022-22751929194210.1194/jlr.M 00574420164094 PMC 2882750 · doi ↗ · pubmed ↗
- 5Iqbal J Boutjdir M Rudel LL Hussain MM 2014716 Intestine-specific MTP and global ACAT 2 deficiency lowers acute cholesterol absorption with chylomicrons and HD Ls.J Lipid Res 55110022-22752261227510.1194/jlr.M 04795125030663 PMC 4617129 · doi ↗ · pubmed ↗
- 6Kolasa AM Bhogal JK Di Angelo JR 2021123 The heterogeneous nuclear ribonucleoprotein (hn RNP) glorund functions in the Drosophila fat body to regulate lipid storage and transport.Biochem Biophys Rep 2510091910091910.1016/j.bbrep.2021.10091933537463 PMC 7838711 · doi ↗ · pubmed ↗
- 7Palm W Sampaio JL Brankatschk M Carvalho M Mahmoud A Shevchenko A Eaton S 2012726 Lipoproteins in Drosophila melanogaster--assembly, function, and influence on tissue lipid composition.P Lo S Genet 871553-7390 e 1002828 e 100282810.1371/journal.pgen.100282822844248 PMC 3406001 · doi ↗ · pubmed ↗
- 8Pihlajamäki J Lerin C Itkonen P Boes T Floss T Schroeder J Dearie F Crunkhorn S Burak F Jimenez-Chillaron JC Kuulasmaa T Miettinen P Park PJ Nasser I Zhao Z Zhang Z Xu Y Wurst W Ren H Morris AJ Stamm S Goldfine AB Laakso M Patti ME 201183 Expression of the splicing factor gene SFRS 10 is reduced in human obesity and contributes to enhanced lipogenesis.Cell Metab 1421550-413120821810.1016/j.cmet.2011.06.00721803291 PMC 3167228 · doi ↗ · pubmed ↗