Emerging role of epigenetic mechanisms in glaucoma and their translational potential
Altaf A. Kondkar, Tahira Sultan, Taif A. Azad, Saleh A. Al-Obeidan

TL;DR
This paper reviews how epigenetic changes contribute to glaucoma and could lead to new diagnostic and treatment approaches.
Contribution
The paper highlights novel epigenetic mechanisms in glaucoma pathogenesis and their potential for translational applications.
Findings
Aberrant DNA methylation and histone modifications contribute to glaucoma progression.
Epigenetic clocks identify glaucoma as an accelerated aging process.
Epigenetic changes are reversible and could restore visual function in animal models.
Abstract
Glaucoma, a leading cause of irreversible blindness, is a complex polygenic disease where significant clinical and genetic heterogeneity do not explain all glaucoma cases, highlighting the need for a deeper understanding of molecular mechanisms like epigenetics. This review examines the emerging role of key epigenetic mechanisms, specifically DNA methylation, histone modifications, and non-coding RNAs in glaucoma pathogenesis and their potential as biomarkers and therapeutic targets. We discuss how aberrant DNA methylation (e.g., GDF7 hypomethylation/CDKN2B hypermethylation) promotes trabecular meshwork fibrosis and increases optic nerve vulnerability, contributing to disease development and/or progression. The METTL23 histone methylation linked to retinal ganglion cell death at normal eye pressure, and disease-specific microRNA profiles further support the role of epigenetic…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
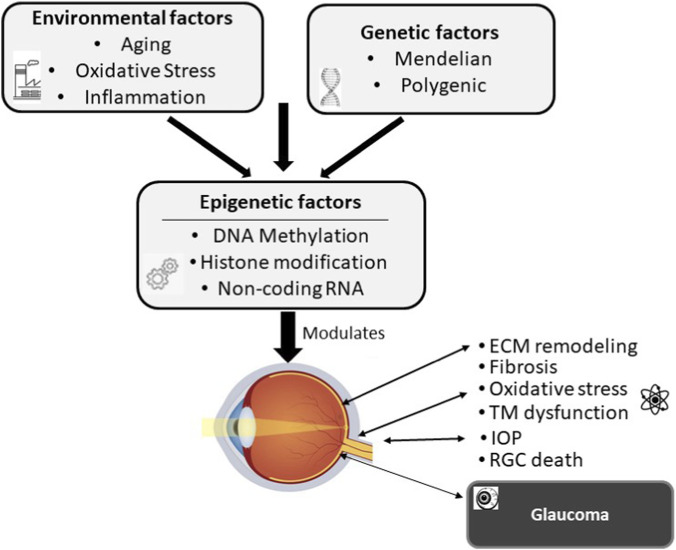 FIGURE 1
FIGURE 1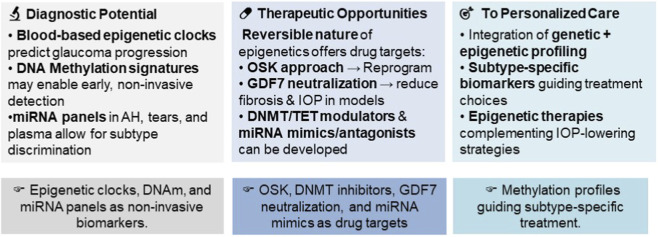 FIGURE 2
FIGURE 2| Glaucoma subtype | Epigenetic mechanism | Specific marker | Change | References |
|---|---|---|---|---|
| PXG | DNA Methylation |
| Hypermethylation |
|
| POAG | DNA Methylation |
| Hypomethylation |
|
| NTG | DNA Methylation |
| Hypermethylation |
|
| NTG | Histone Modification |
| Loss of Function |
|
| PXG | m6A Methylation |
| Upregulated |
|
| POAG | m6A Methylation |
| Downregulated |
|
| Functional role | miRNAs involved | Sample/Source | Postulated mechanism/Relevance | References |
|---|---|---|---|---|
| Anti-Fibrotic/ECM Regulation in TM | miR-26a, miR-29b, miR-139, miR-155, miR-200a/c, miR-18a-5p | Human TM cells, animal models | Downregulate ECM components; inhibit TGF-β/Wnt signaling; reduce outflow resistance |
|
| TM Contractility and IOP Regulation | miR-143/145, miR-200c | Human TM cells, mouse KO models | Regulate actin cytoskeleton; KO reduces IOP, increases outflow facility |
|
| RGC Survival and Neuroprotection | miR-182, miR-100, miR-96 | RGC-5 cells | Modulate apoptosis, oxidative stress; promote survival via AKT/ERK pathways |
|
| Oxidative Stress and Inflammation | miR-27a, miR-182 | Retina, tears | Modulate inflammatory/oxidative responses; potential tear biomarker |
|
| Differential expression of fluid-based circulating miRNAs | miR-125b-5p | Aqueous humor | Down in POAG, up in PXG |
|
| miR-637, miR-1306-5p, miR-3159 | Plasma | AUC = 0.91 for glaucoma detection |
| |
| miR-210-3p | Serum/plasma | Elevated in POAG |
| |
| miR-26b, miR-152, miR-30e, miR-151a | Tear | AUC > 0.75 to differentiate between POAG and OHT |
|
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGlaucoma and retinal disorders · Ophthalmology and Visual Impairment Studies · Retinal Development and Disorders
Introduction
1
Glaucoma is a group of optic neuropathies characterized by elevated intraocular pressure (IOP), progressive loss of retinal ganglion cells (RGCs), damage to the optic nerve, and blindness (Dietze et al., 2025; Weinreb et al., 2014). The estimated worldwide prevalence of adult-onset glaucoma is 3.5% in the population aged 40 years and above, affecting around 80 million people globally, with an estimated projection to nearly 112 million by 2040 (Allison et al., 2020; Tham et al., 2014; Al-Manjoumi et al., 2023). Since elevated IOP is considered a major risk factor for RGC death in glaucoma, the current clinical management is also focused on lowering IOP (Park et al., 2023). However, up to 40% or more of glaucoma patients do not exhibit high IOP but still suffer from progressive optic nerve damage, as seen in patients with normal-tension glaucoma (NTG) (Kim and Park, 2016). These cases demonstrate that there are several other non-IOP factors contributing to glaucoma development.
Glaucoma is a complex multifactorial disease involving both genetic and environmental influences (Wiggs, 2012). A number of genetic studies, including genome-wide, have identified several Mendelian genes (e.g., MYOC, CYP1B1, PAX6, FOXC1), chromosomal loci (GLC1A-P), and DNA variants associated with glaucoma (Kolovos et al., 2025; Tirendi et al., 2023; Zukerman et al., 2020; Abu-Amero et al., 2015). Although these studies have provided significant mechanistic insights into disease pathogenesis, they fail to explain the clinical variability seen in glaucoma onset, progression, and treatment response, suggesting the role of factors beyond genetic determinants (D’Esposito et al., 2024; Wang and Wang, 2023). There is evidence to believe that the environmental factors, such as aging, oxidative stress, and inflammation, interact with the genome in complex ways, inducing aberrant epigenetic regulation that might be the missing link, beyond the genetic determinants (Figure 1) (Wang and Wang, 2023; Dolinoy and Jirtle, 2008; Medeiros et al., 2025).
Conceptual model of epigenetics as the link between genetics and environment in the complex pathogenesis of glaucoma. Environmental factors interact with the genetic blueprint, and epigenetic mechanisms process these signals to regulate and influence crucial pathological outcomes, including extracellular matrix (ECM) remodeling, fibrosis, oxidative stress, trabecular meshwork (TM) dysfunction, intraocular pressure (IOP) alteration, leading to retinal ganglion cell (RGC) death and the eventual clinical manifestation of glaucoma.
Epigenetics refers to the heritable changes observed in gene expression without alteration in the DNA sequence (Kachhawaha et al., 2023). There is strong evidence to suggest that epigenetics might be associated with glaucoma development and progression, plausibly by altering gene expression profiles critical to IOP control, optic nerve integrity, and neuroinflammation. These biological processes are central to aqueous humor dynamics, RGC survival, immune responses, and extracellular matrix (ECM) remodeling, which are strongly implicated in glaucoma (Figure 1) (D’Esposito et al., 2024; Tonti et al., 2024).
Eye represents an excellent model for epigenetic research (Alkozi et al., 2017; Lanza et al., 2019). The purpose of this review is to examine the epigenetic contributions of DNA methylation, histone modifications, N6-methyladenosine (m6A) methylation, and non-coding RNAs in glaucoma pathogenesis and their clinical translational potential.
Epigenetic mechanisms in glaucoma
2
DNA methylation
2.1
DNA methylation is often referred to as ‘the epigenetic switch.’ It involves the addition of a methyl group to a cytosine base, prominently at CpG sites near gene promoters, causing gene silencing when hypermethylated or gene transcription when hypomethylated (Jin and Liu, 2018; Lim and Maher, 2010). Methylation is a tightly regulated process involving specific enzymes that are critical for developmental processes, cellular differentiation, and maintenance of genomic stability. These include the ‘writers’ and ‘maintainers,’ which are the DNA Methyltransferases (DNMTs), like DNMT1 for maintenance and DNMT3A/B for new establishment. The “erasers” are the Ten-Eleven Translocation (TET) enzymes, which control the on/off switch by oxidizing the methyl group (Dan and Chen, 2022). DNA methylation offers a dynamic interface between environmental stimuli (e.g., oxidative stress) and gene expression in ocular tissues. Aberrant expression of these enzymes can lead to abnormal methylation patterns and dysregulation of genes involved in key processes implicated in glaucoma pathogenesis, such as ECM remodeling, oxidative stress, and neurodegeneration (Bou Ghanem et al., 2024; Fernández-Albarral et al., 2024).
Evidence of differential DNA methylation in glaucoma
2.1.1
Several studies have demonstrated aberrant DNA methylation in different types of glaucoma. Junemann et al. reported a significantly higher global DNA methylation in blood cells of POAG patients compared to controls and PXG patients (Junemann et al., 2014). The trabeculectomy sections from glaucomatous eyes were reported to have significant DNA hypomethylation of Alu repetitive elements in POAG and PACG, and significant DNA hypermethylation of HERV-K in POAG patients (Chansangpetch et al., 2018). These methylation changes were proposed to predispose the trabecular meshwork (TM) cells to a profibrotic phenotype (Chansangpetch et al., 2018).
In PXG, the LOXL1 promoter is reported to be hypermethylated in lens capsule and human Tenon fibroblasts that correlated with reduced LOXL1 expression and increased ECM cross-linking (Greene et al., 2020; Ye et al., 2015). Interestingly, treatment of human Tenon Fibroblasts with DNMT inhibitor, 5-aza-dC, restored LOXL1 expression in PXG fibroblasts (Greene et al., 2020). Conversely, Kapuganti et al. reported hypomethylation of the clusterin promoter in patients with PXG (Kapuganti et al., 2023). In another study, CDKN2B gene promoter hypermethylation at the 9p21 locus has been linked to NTG susceptibility in women (Burdon et al., 2018). CDKN2B silencing due to hypermethylaiton causes its suppression that may increase the optic nerve’s vulnerability to non-IOP stressors (Burdon et al., 2018). This study elegantly demonstrated how epigenetic errors through aberrant methylation can impact critical RGC survival pathways, leading to neurodegeneration even at normal IOP.
Mechanistic evidence of DNA methylation in glaucoma
2.1.2
High IOP is the result of decreased outflow capacity at the level of the TM and Schlemm’s canal, the primary exit route of aqueous humor (Carreon et al., 2017). Dexamethasone (DEX)-treated primary human TM cells exhibited hypomethylation of the thrombospondin-1 (THBS1) promoter region and reduced transcript levels of 2 DNA methyltransferases (DNMTs), DNMT1 and DNMT3A (Choy et al., 2025). This change was consistent with DNA methylation inhibitors, 5-azacytosine (5-AC) or 5-aza-2′-deoxycytidine (5-aza-dC), inducing an increase in THBS1 protein levels, leading to the reduced outflow facility ex vivo and increased IOP in vivo adult male C57BL/6 J mice (Choy et al., 2025). These results highlight a clear mechanism in which DNMT downregulation and promoter hypomethylation alter THBS1 expression and influence ECM composition and aqueous outflow resistance. In glaucoma, fibrosis is one of the key mechanisms in glaucoma development and progression, as a result of ECM deposition in the TM at the anterior of the eye, and at the optic nerve head in the lamina cribrosa, leading to aqueous outflow resistance, elevated IOP, and subsequently glaucoma (Keller and Peters, 2022). The GDF7 gene promoter region is aberrantly hypomethylated in glaucomatous TM samples and TET-dependent (Wan et al., 2021). Hypomethylation causes gene activation, which releases the brake, causing GDF7 protein overexpression, increasing proteins like α-SMA, promoting fibrosis which clogs the drainage pathway, raising IOP. This landmark study is a prime example of a targetable epigenetic mechanism in POAG. Crucially, in a therapeutic proof-of-concept, neutralizing GDF7 with antibodies reduced this fibrosis and improved outflow in a primate model. This study showed that GDF7 is a direct, reversible, and therapeutic targetable epigenetic cause of high IOP and fibrosis in POAG, offering a potential for a novel anti-fibrotic therapy (Wan et al., 2021). Likewise, glaucomatous Schlemm’s canal endothelial cells have been shown to exhibit distinct methylation profiles in genes (TGFBR3, TBX3, TNXB1, DAXX, and PITX2) enriched in pathways regulating outflow resistance. However, further studies are needed to validate these findings (Cai et al., 2020).
Taken together, these studies suggest a complex and gene-specific methylation landscape in glaucoma subtypes and illustrate how DNA methylation can act as a molecular switch modulating distinct cellular pathways, promoting ECM remodeling in PXG (via LOXL1 or CLU) and influencing neuroprotective cell-cycle regulation in NTG (via CDKN2B), thereby contributing directly to disease susceptibility. It can thus be speculated that promoter methylation changes may represent potential blood-based biomarkers for glaucoma risk assessment and gender-specific susceptibility, as well as a valuable target for new treatments to mitigate glaucoma or fibrosis-related outflow resistance to restore normal TM function and outflow facility.
Epigenetic reprogramming
2.2
In vivo epigenetic reprogramming using Yamanaka OSK factors (Oct4, Sox2, Klf4) has shown promise in preclinical models. Reprogramming aged or glaucomatous RGCs reverses DNA methylation patterns to a youthful state, restoring a healthy transcriptome and visual function in mice. Transient OSK expression in glaucomatous eyes reversed vision loss in mice. The effect was dependent on DNA demethylases TET1 and TET2 (Lu et al., 2020). Moving beyond the initial proof-of-concept, a follow-up study provided compelling evidence of long-term safety and efficacy for vision recovery in glaucoma (Karg et al., 2023). The studies provide groundbreaking evidence of epigenetic reprogramming as a viable and sustainable approach for recovering lost vision in glaucoma with direct neural rescue and repair, moving us closer to a future where we can complement IOP-lowering.
DNA methylation-based EpiScores
2.3
Recent evidence suggests that DNA methylation (DNAm)-based EpiScores and GrimAge acceleration are positively associated with glaucoma risk, indicating that the disease may represent an accelerated molecular aging process (Medeiros et al., 2025; Jiang et al., 2025). EpiScore and GrimAge are epigenetic clock that predicts biological age and mortality risk based on DNAm patterns. Accelerated epigenetic aging compared to chronological aging is associated with a 15% higher odds of faster glaucoma progression. Importantly, this relationship remains strong even in patients with relatively low IOP, indicating that epigenetic aging may predispose the optic nerve to damage from things like oxidative stress, independent of IOP level (Medeiros et al., 2025). These results highlight the potential clinical importance of epigenetic age acceleration as a non-invasive predictor of progression.
Histone modifications
2.4
Histone modifications, including acetylation, methylation, phosphorylation, and ubiquitination, can regulate gene expression by altering chromatin structure (Davie and Chadee, 1998; Zhang et al., 2015). These modifications, catalyzed by specific enzymes, play significant roles in regulating genes involved in maintenance of IOP and RGC health, thereby influencing glaucoma risk and severity (Tonti et al., 2024; Feng et al., 2023).
Studies on glaucoma-related histone modifications have mainly focused on deacetylation (Pelzel et al., 2010; Schmitt et al., 2014; Alsarraf et al., 2014; Sharma et al., 2016). Histone deacetylation functions as a crucial epigenetic switch that may accelerate glaucomatous neurodegeneration (Pelzel et al., 2010; Sharma et al., 2016; Biermann et al., 2011). The HDAC inhibitors have consistently demonstrated to exhibit neuroprotective effects and improved surgical outcomes in different animal models of glaucoma, highlighting their potential utility as pharmacological agents in glaucoma (Feng et al., 2023; Schmitt et al., 2014; Alsarraf et al., 2014; Sharma et al., 2016; Biermann et al., 2011).
Histone methylation is another critical regulatory mechanism in glaucoma. The trimethylation mark H3K27me3, catalyzed by EZH2 are both detected in RGCs (Rao et al., 2012). A study by Rao et al. (2010) showed that inhibition of EZH2 induces RGC apoptosis, underscoring the essential role of EZH2-mediated H3K27me3 in retinal neuronal survival and suggesting its involvement in glaucomatous degeneration (Rao et al., 2010).
METTL23 and NTG: a histone methylation connection
2.4.1
Mutations in the METTL23 gene encoding a histone arginine methyltransferase have been linked to NTG (Pan et al., 2022). METTL23 catalyzes the dimethylation of H3R17 in the retina. The c.A23G mutation inherited as an autosomal dominant condition results in METTL23 loss of function, leading to impaired dimethylation of histone H3R17. This disrupts gene regulation in RGCs, causing RGC apoptosis through NF-κB signaling. The study highlights a novel epigenetic etiology in NTG and provides a direct evidence linking abnormal histone methylation to glaucomatous neurodegeneration (Pan et al., 2022; Scheetz et al., 2024; Liu and Sun, 2022). In addition, the findings open new therapeutic avenues to target the downstream inflammatory NF-κB signaling or the histone methylation modulators, shifting the paradigm beyond the conventional IOP-based approach of glaucoma management.
N6-methyladenosine (m6A) modification
2.5
m6A is a common mRNA modification regulating RNA stability, splicing, translation, and decay, affecting gene expression (Jiang et al., 2021; Patil et al., 2018). The modification involves regulation by writers (e.g., methyltransferase-like protein 3, METTL3), erasers (e.g., fat mass and obesity-associated, FTO), and readers (e.g., YTH family protein YTHDC2) (Jiang et al., 2021). These writer, eraser, and reader proteins play a significant role in various diseases, but are less studied in glaucoma.
In PXG, elevated global m6A levels and upregulated writer and reader enzymes (e.g., METTL3, YTHDC2) in aqueous humor may serve as biomarkers, with m6A-modified transcripts enriched in matrix organization pathways (D’Esposito et al., 2024; Guan et al., 2023). In another study, differential m6A-methylated lncRNAs in PXG aqueous humor influenced glaucoma-related genes and processes (Guan et al., 2024). In POAG, bioinformatics analyses identified differentially expressed m6A regulators (e.g., upregulated reader YTHDF1, downregulated reader YTHDC2) in TM tissues. Silencing YTHDC2 in human TM cells enhanced migration and ECM synthesis, demonstrating a functional role in outflow resistance (Zhang et al., 2025). Current studies suggest that targeting writer and reader pathways could lead to new treatments. For example, blocking the writer METTL3 might control fibrosis in PXG, and silencing readers like YTHDC2 may prevent TM dysfunction in POAG (Tonti et al., 2024; Guan et al., 2024; Zhang et al., 2025).
To enhance clinical utility and highlight the diagnostic potential of the epigenetic changes discussed in the above sections, a summary of the key markers characterized by their subtype-specific roles in POAG, PXG, and NTG is listed in Table 1.
Noncoding RNAs
2.6
Noncoding RNAs (ncRNAs) like microRNAs (miRNAs), long non-coding RNAs (lncRNAs), and circular RNAs (circRNAs) are pivotal players in cell regulatory networks across a broad spectrum of biological processes implicated in glaucoma (Rong et al., 2021; Costa et al., 2025). Differential expression and abnormal function of these regulatory molecules can affect gene networks related to fibrosis, IOP, and optic nerve health, making them promising biomarker candidates and drug targets (Rong et al., 2021; Zhang et al., 2021; Huang et al., 2022).
MiRNAs
2.6.1
miRNAs are the most extensively studied class of ncRNAs in glaucoma and have been elegantly reviewed elsewhere for detailed molecular summaries (Dobrzycka et al., 2023; Greene et al., 2022; Martinez and Peplow, 2022). Several studies have confirmed that these small RNA molecules (∼19–23 nucleotides in length) regulate a wide range of pathological processes, including ECM remodeling, TM cell function, RGC apoptosis, and oxidative stress.
In the TM, multiple miRNAs (e.g., miR-29b and miR-200a) exert anti-fibrotic effects by targeting pathways like TGF-β and Wnt/β-catenin to counteract ECM accumulation (Keller and Peters, 2022; Luna et al., 2009; Luna et al., 2011; Luna et al., 2012; Yu et al., 2022). miRNAs such as miR-143/145 and miR-200c regulate TM cell contractility and IOP, with knockout studies demonstrating reduced IOP and improved outflow in mice (Luna et al., 2012; Li et al., 2017). Others, including miR-18a-5p, also target TGF-β2 signaling to reduce contractility and fibrosis (Knox et al., 2022). Beyond IOP regulation, miRNAs like miR-182, miR-100, and miR-96 influence RGC survival and offer neuroprotection (Kong et al., 2014; Li et al., 2019; Wang and Li, 2014), while others such as miR-27a modulate oxidative stress and inflammatory responses (Tabak et al., 2021).
miRNAs in ocular fluids show promise as biomarkers. Profiles in aqueous humor and plasma are disease-specific, aiding in subtype differentiation (Drewry et al., 2018; Hindle et al., 2019; Rao et al., 2020; Raga-Cervera et al., 2021; Tanaka et al., 2014; Kosior-Jarecka et al., 2021). For example, specific miRNAs like miR-125b-5p are differentially expressed in POAG versus PXG (Drewry et al., 2018). A plasma panel of miR-637, miR-1306-5p, and miR-3159 identified glaucoma patients with an AUC of 0.91 (Hindle et al., 2019), and serum miR-210-3p is elevated in POAG (Liu et al., 2019; Zhao et al., 2023). In addition, tear-based miRNAs have shown promise as a non-invasive method for glaucoma screening (Raga-Cervera et al., 2021). Moreover, miRNAs identified in aqueous humor (e.g., miR-143-3p, miR-125b-5p, and miR-1260b) have been proposed to serve as drug targets (Martinez and Peplow, 2022; Zhang et al., 2023). The functional roles and diagnostic potential of key identified miRNAs are summarized in Table 2.
LncRNAs
2.6.2
LncRNAs are transcripts over 200 nucleotides that often act as miRNA sponges (Mattick et al., 2023). By impairing the biological activity of miRNAs, lncRNAs increase the production of proteins from the target mRNAs and their dysregulation is implicated in glaucoma. Key lncRNAs like MALAT1, ANRIL (also known as CDKN2B-AS1), and others (ENST00000552367 and NR_038379) influence TM cell proliferation, ECM remodeling, and RGC vulnerability (Burdon et al., 2018; Zhang et al., 2021; Huang et al., 2022; Pasquale et al., 2013). Xie et al. (2019) suggested that lncRNAs T267384, ENST00000607393, and T342877 might be potential therapy biomarkers for POAG. Some, including NR003923, H19, and LINC00028 are prospective targets for preventing post-surgical fibrosis (Yu et al., 2022; Zhu et al., 2020; Zhao et al., 2019; Sui et al., 2020). However, the functions and associated mechanisms of the role of most lncRNAs in glaucoma are not completely clear yet.
CircRNAs
2.6.3
Similarly, circRNAs are closed-loop molecules that sequester miRNAs to regulate gene expression (Wawrzyniak et al., 2018). A recent study demonstrated that the circRNA ZRANB1 is expressed in glial cells and it negatively regulates miR-217, promoting Müller cell proliferation and RGC apoptosis (Wang et al., 2018). The expression of cZRANB1 was upregulated in glaucoma-induced retinal degeneration, and its knockdown provided a protective effect by reducing retinal gliosis and RGC apoptosis. This effect was reversed by overexpression of RUNX2 (Wang et al., 2018). Targeting this cZRANB1/miR-217/RUNX2 network has neuroprotective potential (Wang et al., 2018). Recent research has identified some candidate circRNAs in ocular hypertension glaucoma models by high-throughput sequencing (Chen et al., 2020); however, the role of most circRNAs in glaucoma remains to be elucidated (Choudhari et al., 2025).
Translational and clinical potential
3
The epigenetic mechanisms discussed in this review have significant translational and clinical potential to improve glaucoma diagnosis, prognosis, and treatment (Figure 2). Epigenetic aging derived from blood samples (EpiScore) could serve as non-invasive biomarkers of disease susceptibility (Medeiros et al., 2025). Likewise, DNA methylation profiles (e.g., hypermethylation of LOXL1 in PXG or CDKN2B in NTG) and circulating miRNA panels in aqueous humor, tears, or plasma may enable early detection and differentiation between glaucoma subtypes (Greene et al., 2020; Burdon et al., 2018; Martinez and Peplow, 2022). The inherent reversible nature of epigenetic modifications makes them attractive drug targets for clinical intervention. For instance, the epigenetic reprogramming of RGCs using the OSK approach could be the future paradigm for glaucoma treatment to promote RGC survival and restore vision (Lu et al., 2020; Karg et al., 2023). Similarly, with the demonstrated proof-of-concept, GDF7 neutralization might emerge as a promising anti-fibrotic therapy in glaucoma (Wan et al., 2021). In addition, the development of key epigenetic enzyme inhibitors (e.g., DNMT or HDAC inhibitors) and RNA-based therapies (miRNA antagonists) may precisely rectify dysregulated glaucoma pathways in the TM and retina, to reduce fibrosis, lower IOP, and protect the RGCs.
Translational and clinical potential of epigenetics in glaucoma. Schematic representation of the pathways through which epigenetic research is transitioning from mechanistic insight to clinical application, highlighting opportunities in diagnostics, therapeutics, and personalized care.
Challenges and future perspectives
4
Despite the exciting avenues of epigenetics, there exist several challenges to effectively translate the potential of epigenetics to patient care. The current evidence is mostly based on preclinical and small-scale human studies, highlighting the need to validate the diagnostic and prognostic potential of biomarkers like EpiScore, DNA methylations, and miRNA panels in a diverse population-based longitudinal cohorts. To address these challenges, future research must utilize longitudinal studies and Mendelian randomization to ascertain whether these epigenetic marks are a primary drivers of glaucoma or are secondary consequences of the disease process and cellular stress. Integrating multi-omics data (genomics, epigenomics, and transcriptomics) will be essential to validate these markers as causative agents of disease progression. Furthermore, the high degree of tissue-specific origins of epigenetic changes also presents a significant problem. Consequently, mechanistic studies are required to determine if these changes in accessible samples, such as blood or tears, accurately reflect localized changes in the TM or the optic nerve.
Development of advanced ocular drug delivery systems, such as nanoparticle-based eye drops or viral vectors for CRISPR-based epigenetic editing, is highly crucial for safe and effective application of epigenetic modulators (e.g., OSK factors, GDF7, DNMT/HDAC inhibitors, miRNA mimics). Additionally, studies to investigate the effect of environmental factors like diet, exercise, and lifestyle on the ocular epigenetics would be vital steps towards preventative medicine. The ultimate goal will be to incorporate validated genetic and epigenetic data with routine clinical markers, such as IOP, visual fields, and optical coherence tomography, to pave the way for personalized medicine in glaucoma.
Conclusion
5
In conclusion, epigenetics redefines our understanding of glaucoma from a static pressure disorder to a modifiable disease at the interface of genetics and environment. Although there are challenges in terms of validation, causality, and drug delivery, the reversible nature of epigenetic mechanisms provides a powerful therapeutic opportunity. Future studies are needed to address these limitations to utilize epigenetics as a source of novel biomarkers, specific drug targets, and personalized care strategies to prevent vision loss beyond IOP control alone.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Abu-Amero K. Kondkar A. A. Chalam K. V. (2015). An updated review on the genetics of primary open angle glaucoma. Int. J. Mol. Sci. 16, 28886–28911. 10.3390/ijms 161226135 26690118 PMC 4691082 · doi ↗ · pubmed ↗
- 2Al-Manjoumi A. M. Filimban M. A. Bantan M. S. Abualhamayel H. A. Youldash O. A. (2023). Prevalence of risk factors among patients with glaucoma in jeddah, Saudi Arabia. Cureus 15, e 37689. 10.7759/cureus.37689 37206536 PMC 10191456 · doi ↗ · pubmed ↗
- 3Alkozi H. A. Franco R. Pintor J. J. (2017). Epigenetics in the eye: an overview of the Most relevant ocular diseases. Front. Genet. 8, 144. 10.3389/fgene.2017.00144 29075285 PMC 5643502 · doi ↗ · pubmed ↗
- 4Allison K. Patel D. Alabi O. (2020). Epidemiology of glaucoma: the past, present, and predictions for the future. Cureus 12, e 11686. 10.7759/cureus.11686 33391921 PMC 7769798 · doi ↗ · pubmed ↗
- 5Alsarraf O. Fan J. Dahrouj M. Chou C. J. Yates P. W. Crosson C. E. (2014). Acetylation preserves retinal ganglion cell structure and function in a chronic model of ocular hypertension. Investig. Ophthalmol. Vis. Sci. 55, 7486–7493. 10.1167/iovs.14-14792 25358731 PMC 4240722 · doi ↗ · pubmed ↗
- 6Biermann J. Boyle J. Pielen A. Lagrèze W. A. (2011). Histone deacetylase inhibitors sodium butyrate and valproic acid delay spontaneous cell death in purified rat retinal ganglion cells. Mol. Vis. 17, 395–403. 21311741 PMC 3036563 · pubmed ↗
- 7Bou Ghanem G. O. Wareham L. K. Calkins D. J. (2024). Addressing neurodegeneration in glaucoma: mechanisms, challenges, and treatments. Prog. Retin. Eye Res. 100, 101261. 10.1016/j.preteyeres.2024.101261 38527623 · doi ↗ · pubmed ↗
- 8Burdon K. P. Awadalla M. S. Mitchell P. Wang J. J. White A. Keane M. C. (2018). DNA methylation at the 9p 21 glaucoma susceptibility locus is associated with normal-tension glaucoma. Ophthalmic Genet. 39, 221–227. 10.1080/13816810.2017.1413659 29265947 · doi ↗ · pubmed ↗