Polyamines as a Universal Language of Host–Microbiota Symbiosis
Xiuyu Fang, Yan Guo, Jia Huang, Meimei Zhang

TL;DR
Polyamines, produced by gut microbes and influenced by the host, play a key role in maintaining health and treating diseases like inflammatory bowel disease.
Contribution
The paper highlights polyamines as a central mediator in host-microbiota interactions with therapeutic potential.
Findings
Polyamines support epithelial integrity and immune regulation.
Disruption of polyamine metabolism worsens inflammation and barrier dysfunction.
Engineered therapies targeting polyamine metabolism show promise for treating age-related diseases.
Abstract
Polyamines are ancient metabolites that serve critical functions in maintaining epithelial integrity, regulating immune response, and supporting healthy aging. The gut microbiota actively synthesizes and converts polyamines, while host factors such as inflammation, barrier function, and nutritional status dynamically modulate this metabolic network. Disruption of this host–microbiota axis reduces polyamine availability, impairs barrier function, and exacerbates inflammation. In contrast, polyamines exert protective effects by promoting epithelial repair, modulating macrophage and T-cell responses, and enhancing autophagy-mediated tissue renewal and longevity. Recent advances in engineered probiotics, microbial small RNAs, and postbiotics further highlight the therapeutic potential of precisely modulating polyamine metabolism in clinical contexts such as inflammatory bowel disease,…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
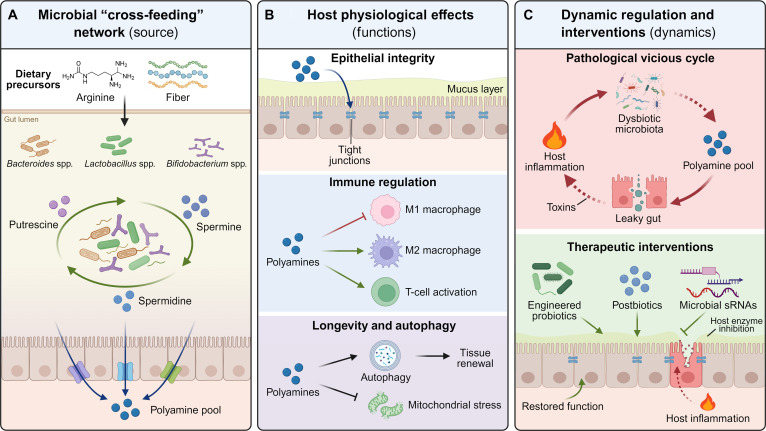 Figure 1
Figure 1- —National Natural Science Foundation of Chinahttp://dx.doi.org/10.13039/501100001809
- —the CAST Youth Talent Support Project-Special Program for Doctoral Students (156-O-230-0000375-5)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPolyamine Metabolism and Applications · Bacterial biofilms and quorum sensing · Biopolymer Synthesis and Applications
Polyamines, among the most ancient and resilient small molecules, play a pivotal role in cellular proliferation, immune regulation, mitochondrial homeostasis, and aging processes [1]. One of the most exciting discoveries in recent years reveals that the gut microbiota acts not merely as a passive source of polyamines but also as an active regulator of their systemic homeostasis. This insight compels us to re-evaluate the metabolic symbiosis between host and microbiota and opens new avenues for understanding inflammatory diseases, cancer, metabolic disorders, and healthy aging. The gut microbiota contributes to polyamine pools through a complex network of biosynthetic, interconversion, and regulatory pathways [2]. Typical gut bacteria, including Bacteroides, Lactobacillus, and Bifidobacterium, harbor core enzymatic systems such as ornithine decarboxylase and agmatine deaminase [3]. These metabolic capacities enable the conversion of amino acid precursors into putrescine, spermidine, and spermine, thereby establishing community-level metabolic networks based on “cross-feeding”. In such metabolic networks, polyamine intermediates produced by one bacterial species can be assimilated and further modified by other members. For example, spermine generated by one taxon may be converted into putrescine by a second. This intermediate can then be elongated into spermidine or spermine by a third species. Consequently, the overall polyamine profile reflects the integrated metabolic capacity of the microbial community rather than the abundance of any single taxon. This distributed metabolic cooperation underlies the stability and functional resilience of the gut ecosystem. At the same time, it reveals critical vulnerabilities that can be disrupted by dietary changes, inflammation, antibiotic exposure, or other environmental disturbances. However, this intricate microbial metabolic network does not function independently. Instead, it is tightly coupled to host physiology through continuous, bidirectional interactions.
In this dialog, host physiology exerts profound feedback regulation over microbial polyamine metabolism. The host’s inflammatory state, intestinal barrier permeability, and nutrient availability collectively shape the microbiota’s capacity to synthesize and interconvert polyamines [4]. In disease states, host polyamine metabolism undergoes marked alterations [3]. For instance, during bloodstream infection, bacterial proliferation elevates levels of acetylated polyamines, while the disruption of the polyamine-producing enzyme SpeG reduces bacterial growth and attenuates disease progression [5]. Moreover, under pathological conditions such as colitis, the abundance of polyamine-producing bacteria is markedly diminished. This reduction leads to impaired polyamine metabolism, compromised intestinal barrier function, and exacerbated mucosal injury [6]. This dynamic establishes a classic vicious cycle of inflammation, polyamine depletion, barrier dysfunction, and microbial dysbiosis, forming a critical pathological basis for numerous chronic gastrointestinal disorders.
Polyamines exert broad and profound regulatory effects on host physiology. Intestinal epithelial cells are highly dependent on spermidine to maintain tight junction proteins, mitochondrial integrity, and mucus layer renewal [7]. Conversely, a polyamine-deficient intestinal epithelium rapidly exhibits barrier disruption [1]. Immune cells such as macrophages are also highly sensitive to polyamines. Putrescine and spermidine can reshape their metabolism, promoting a more reparative M2 phenotype and suppressing excessive inflammatory responses [8]. Polyamine metabolism in macrophages contributes to the maintenance of colonic epithelial homeostasis, with polyamines serving as a critical molecular bridge linking immunometabolism to epithelial repair [9,10]. Moreover, polyamine biosynthesis is essential for the initial activation of T cells [11]. More intriguingly, accumulating evidence links polyamines to lifespan extension, tissue regeneration, and antiaging, largely through their roles in promoting autophagy and maintaining mitochondrial quality control [12]. These pleiotropic effects depend both on local polyamine concentrations and their multi-tissue distribution, which are collectively shaped by the synthetic capacity of the gut microbiota, epithelial transport mechanisms, and the degradation kinetics of host enzymes [13].
Building on these advances, therapeutic strategies targeting polyamine metabolism are poised to offer marked clinical potential. Engineered probiotics may serve as “metabolic factories” capable of high-yield production of spermidine or agmatine, enabling more stable and controllable polyamine delivery. Professor Ren and his colleagues [14] uncovered a groundbreaking mechanism by which Lactobacillus murinus-derived small RNAs suppress host polyamine metabolism by inhibiting polyamine metabolic enzymes, thereby establishing a new paradigm of microbial RNA-mediated regulation of host polyamines in colonic diseases. Compared to traditional probiotics, these “functional microbes” hold promise as a new generation of precision nutrition tools [15]. Concurrently, the emergence of postbiotics has opened novel application scenarios for polyamines [16,17]. For instance, stable spermidine derivatives or gut-targeted microbial fermentation products offer enhanced safety and greater precision of delivery. Dietary interventions remain equally important. Diets enriched in arginine, fructooligosaccharides, resistant starch, or polyphenols can systematically enhance polyamine production by reshaping microbial metabolic pathways, thereby improving host health.
Despite these promising prospects, several fundamental scientific questions remain unanswered. Identifying which microbial communities predominantly govern the synthesis and turnover of the endogenous polyamine pool would lay the groundwork for microbiota-targeted interventions and precision nutritional strategies. Determining whether tissue-specific polyamine requirements exhibit circadian rhythmicity could have important implications for chrono-nutrition and time-dependent therapeutic modulation. At a broader level, elucidating how polyamine signaling integrates with short-chain fatty acids, bile acids, and tryptophan metabolism will be essential for building comprehensive metabolic regulatory networks from a systems biology perspective. In addition, polyamines exhibit clear double-edged properties in inflammatory and tumor microenvironments. While they are indispensable for macrophage-mediated resolution of intestinal inflammation, these same molecules can be co-opted within tumors to drive the polarization of tumor-associated macrophages, thereby facilitating tumor progression and immune escape [18,19]. Achieving spatiotemporally specific regulation of polyamine signaling therefore represents a critical hurdle for the development of safe and effective targeted therapies [20].
Overall, the gut microbiota–polyamine metabolism axis offers a new perspective on host–microbiota symbiosis. Moving beyond a simple metabolic supply–demand relationship, it operates as a sophisticated, multilevel communication system that spans barrier integrity, immunity, metabolism, and aging. Understanding and manipulating this system holds promise for delivering novel solutions to inflammatory diseases, metabolic syndrome, neurodegenerative disorders, and even extending healthy lifespan. Taken together, current findings converge on a unifying conclusion: rather than functioning merely as metabolites, polyamines constitute one of the most promising molecular “languages” mediating communication between the host and its microbiota (Fig. 1).
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Minetti A, Omrani O, Brenner C, Cansiz F, Imada S, Rösler J, Khawaled S, Allies G, Meckelmann SW, Gebert N, et al. Polyamines sustain epithelial regeneration in aged intestines by modulating protein homeostasis. Nat Cell Biol. 2025;27(12):2063–2077.41286441 10.1038/s 41556-025-01804-9PMC 12717014 · doi ↗ · pubmed ↗
- 2Li X, Xiao X, Wang S, Wu B, Zhou Y, Deng P. Uncovering de novo polyamine biosynthesis in the gut microbiome and its alteration in inflammatory bowel disease. Gut Microbes. 2025;17(1):2464225.39924644 10.1080/19490976.2025.2464225 PMC 11812404 · doi ↗ · pubmed ↗
- 3Zhang C, Zhen Y, Weng Y, Lin J, Xu X, Ma J, Zhong Y, Wang M. Research progress on the microbial metabolism and transport of polyamines and their roles in animal gut homeostasis. J Animal Sci Biotechnol. 2025;16(1):57.10.1186/s 40104-025-01193-x PMC 1199841840234982 · doi ↗ · pubmed ↗
- 4Wang Y, He F, Liu B, Wu X, Han Z, Wang X, Liao Y, Duan J, Ren W. Interaction between intestinal mycobiota and microbiota shapes lung inflammation. i Meta. 2024;3(5): Article e 241.39429884 10.1002/imt 2.241PMC 11487552 · doi ↗ · pubmed ↗
- 5Mayers JR, Varon J, Zhou RR, Daniel-Ivad M, Beaulieu C, Bhosle A, Glasser NR, Lichtenauer FM, Ng J, Vera MP, et al. A metabolomics pipeline highlights microbial metabolism in bloodstream infections. Cell. 2024;187(15):4095–4112.e 21.38885650 10.1016/j.cell.2024.05.035PMC 11283678 · doi ↗ · pubmed ↗
- 6Nakamura A, Matsumoto M. Role of polyamines in intestinal mucosal barrier function. Semin Immunopathol. 2025;47(1):9.39836273 10.1007/s 00281-024-01035-4PMC 11750915 · doi ↗ · pubmed ↗
- 7Fang X, Liu H, Liu J, Du Y, Chi Z, Bian Y, Zhao X, Teng T, Shi B. Isobutyrate confers resistance to inflammatory bowel disease through host–microbiota interactions in pigs. Research. 2025;8:0673.40342298 10.34133/research.0673 PMC 12059313 · doi ↗ · pubmed ↗
- 8Nakamura A, Kurihara S, Takahashi D, Ohashi W, Nakamura Y, Kimura S, Onuki M, Kume A, Sasazawa Y, Furusawa Y, et al. Symbiotic polyamine metabolism regulates epithelial proliferation and macrophage differentiation in the colon. Nat Commun. 2021;12(1):2105.33833232 10.1038/s 41467-021-22212-1PMC 8032791 · doi ↗ · pubmed ↗