Variable chlorhexidine MICs across Klebsiella species from a single facility
David Lehman, Aubrey E. Hetzler, Tayloe Friedrich, Madeline M. Strouse, Katie E. Barry, Shireen M. Kotay, Amy J. Mathers

TL;DR
This study found varying levels of resistance to chlorhexidine among different Klebsiella species in a hospital, which could affect microbial populations.
Contribution
The study reveals species-specific differences in chlorhexidine resistance and potential chromosomal factors involved.
Findings
Klebsiella quasipneumoniae showed the highest chlorhexidine MICs.
No clear link to acquired resistance was found, but some species had more chromosomal efflux pumps.
Chlorhexidine MIC differences suggest biocides influence hospital microbial populations.
Abstract
We assessed in vitro chlorhexidine minimal inhibitory concentrations (MICs) across Klebsiella species from a hospital with widespread chlorhexidine use. Isolates underwent MIC testing and whole genome sequencing. Species showed varying resistance, with Klebsiella quasipneumoniae having the highest MICs. There was no clear link to acquired resistance, but some species had more chromosomal efflux pumps. Differences in chlorhexidine MICs between species highlight the role that biocides could have in shaping microbial populations in the hospital environment.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
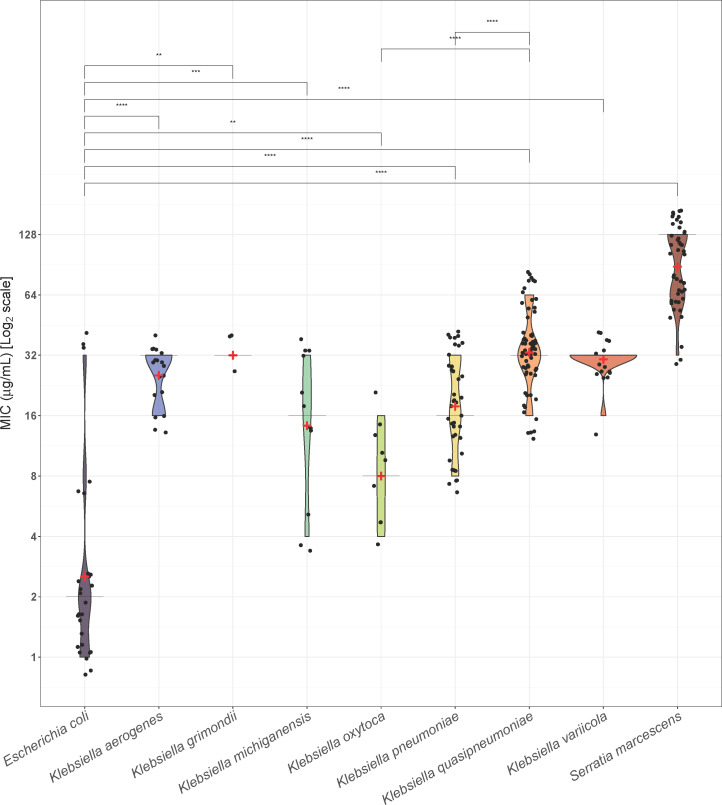 Fig 1
Fig 1| MIC50 µg/mL | MIC90 µg/mL | Range µg/mL | Clinical | Standard | Environmental | |
|---|---|---|---|---|---|---|
| 16 | 32 | 8–32 | 10 | 3 | 0 | |
| 32 | 64 | 16–64 | 10 | 1 | 11 | |
| 8 | 16 | 4–16 | 2 | 0 | 1 | |
| 32 | 32 | 16–32 | 6 | 0 | 0 | |
| N/A | N/A | 32–32 | 0 | 0 | 1 | |
| 32 | 32 | 16–32 | 5 | 0 | 0 | |
| 16 | 32 | 4–32 | 3 | 0 | 1 | |
| 2 | 8 | 1–32 | 7 | 1 | 1 | |
| 128 | 128 | 32–128 | 14 | 1 | 0 |
- —National Institutes of Healthhttp://dx.doi.org/10.13039/100000002
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntimicrobial agents and applications · Antibiotic Resistance in Bacteria · Infection Control in Healthcare
INTRODUCTION
The genus Klebsiella encompasses diverse species occupying both clinical and environmental niches. Klebsiella pneumoniae sensu stricto has emerged as one of the most consequential multidrug-resistant pathogens worldwide, with carbapenemase-producing K. pneumoniae representing an urgent public health threat and a major cause of hospital-associated infections in vulnerable patients (1). Hospital wastewater systems, including sink drains, P-traps, and toilets, are increasingly recognized as persistent reservoirs for carbapenemase-producing Enterobacterales and provide a niche for carbapenemase gene exchange, including among Klebsiella sp. (2, 3).
Species-level identification within the K. pneumoniae complex, including K. pneumoniae sensu stricto, Klebsiella variicola, and Klebsiella quasipneumoniae, is problematic using standard clinical microbiology methods and often requires whole-genome sequencing for accurate classification (4, 5). Although species across the K. pneumoniae complex have been documented to cause infections, their natural niches and virulence vary (6). Despite their prevalence in both patient and environmental reservoirs, the ecological and selective forces shaping species distribution within the hospital setting remain poorly defined, but carbapenemase gene exchange can occur between species (7, 8).
Chlorhexidine is widely used in healthcare as a patient skin antiseptic, hand hygiene agent, and equipment disinfectant (9, 10). While full clinical resistance (i.e., no longer able to achieve killing at topical concentrations) at clinically applied concentrations is unlikely for any Enterobacterales (11), interspecies variation in minimal inhibitory concentrations (MICs) has been described, and selective pressure from antiseptic use is a longstanding concern in infection control (12, 13). Outbreaks associated with Serratia marcescens, a species with intrinsic chlorhexidine elevated MICs, underscore the potential for disinfectants to influence species persistence and selection within hospital environments (11, 12). Whether such selective effects extend across the Klebsiella genus remains unclear, despite their importance as environmental and clinical reservoirs of antimicrobial resistance. This study leveraged whole-genome sequencing to accurately speciate Klebsiella isolates collected from environmental reservoirs and patients. Our goal was to evaluate whether chlorhexidine MICs vary across the Klebsiella genus within a single hospital.
Isolates were previously collected from the University of Virginia, where all intensive care unit hand hygiene soap is SCRUB-STAT (Ecolabs, St. Paul, MN) with 2% chlorhexidine gluconate. We used previously sequenced isolates across species enriched for blaKPC-positive Klebsiella sp. isolates from patients and the environment between 2007 and 2024. We also used several known species from curated strains from the American Type Culture Collection (ATCC Manassas, VA) as species controls where available. Chlorhexidine was diluted in sterile water and then applied to Cation-Adjusted Müeller Hinton Broth (CAMHB), with final concentrations ranging from 0.125 μg/mL to 128 μg/mL in doubling dilutions using the methods described in Lutgring et al. (14). To achieve the highest test concentration of 128 μg/mL without precipitation, an increase from 64 μg/mL previously described, both CAMHB and chlorhexidine solutions were pre-warmed to 37°C prior to combining. All isolates were tested in triplicate. All isolates underwent whole-genome sequencing using a MiSeq v2 300 cycle Reagent Kit on the MiSeq platform (Illumina, San Diego, CA), and some additional isolates had also previously undergone long read sequencing using PacBio or Oxford Nanopore as previously described (15, 16). Speciation and strain typing were performed by calculating the genomic distances of the read sets against a curated collection of complete bacterial chromosomal assemblies in National Center for Biotechnology Information (NCBI) RefSeq database using MASH (17) (v2.1). De novo assembly was checked against the AMRFinder database in NCBI with >99% identity and coverage. Isolates were selected to capture a variety of species within the genus Klebsiella and to compare them with Escherichia coli, which serves as a standard, and S. marcescens, known to exhibit higher MICs (18). To evaluate differences in log2-transformed MIC values among bacterial species, we first conducted a Kruskal-Wallis test to assess overall group differences (α = 0.05). Post-hoc pairwise MICs between species with comparisons using Dunn’s test, implemented from the rstatix R package (v. 0.7.2), with a Bonferroni correction.
We tested 78 unique isolates; 57 clinical, 15 environmental, and 6 ATCC against chlorhexidine (Table 1). The technique had good reproducibility with all replicates being within one dilution, thus 100% essential agreement across the triplicate testing with the median used as the MIC.
In our study, like others, E. coli generally exhibited lower MICs compared to all Klebsiella species, including K. pneumoniae (P = 0.02), Klebsiella aerogenes (P = 0.001), K. quasipneumoniae, K. variicola, and S. marcescens (all P<0.0001) but not Klebsiella grimondii, Klebsiella oxytoca, and Klebsiella michiganensis, although numbers are small (Fig. 1, Table S2). A larger prior evaluation showed E. coli had an MIC₉₀ of 16 µg/mL, whereas K. pneumoniae demonstrated an MIC₉₀ of 32 µg/mL (19). Interestingly, in this collection, several K. quasipneumoniae isolates displayed higher MICs, with an MIC_50_ of 32 and MIC₉₀ of 64 µg/mL. Using the Dunn statistical test, MICs of K. quasipneumoniae were significantly higher than K. oxytoca (P = 0.002) and K. pneumoniae sensu stricto (P = 0.004).
*Violin plots depicting MIC distribution for chlorhexidine by species. Red plus signs indicate the mean log2(MIC) concentration for each species, while black horizontal bars represent the median values. A Kruskal-Wallis test indicates an overall statistically significant difference in MIC distributions across species.The bars represent pairwise comparisons using a Wilcox Test. **: P ≤ 0.01, ***: P ≤ 0.001, ***: P ≤ 0.0001.
We further evaluated resistance gene content across this multidrug-resistant collection using AMRFinder (NCBI). Predicted genes included quaternary ammonium compounds (qac) determinants that may be plasmid borne and drive transferable efflux in gram positives, but these were not consistently found, nor was there a clear correlation with chlorhexidine MICs (Fig. S1 and S2). Previous studies in gram-negative bacilli have failed to demonstrate that mobile AMR genes promote chlorhexidine resistance conclusively but rather the elevated MICs are more likely selected by chromosomal genes and mechanisms allowing resistance to killing, such as outer membrane differences and effective efflux pumps (20, 21). Interestingly, both the K. variicola and K. quasipneumoniae consistently had resistance nodulation cell division family oqxB efflux pumps, as did several of the K. pneumoniae (22). In terms of gene loci, across the 32 isolates with long-read assemblies, all oqx genes were chromosomally encoded in Klebsiella species, tet(41) was consistently chromosomal in Serratia, and in contrast, all qac genes, tet(A) and tet(B), when present were located on plasmids across several species and genera (Table S3).
At our institution, where chlorhexidine-impregnated soap is routinely used, we have found ongoing recovery of blaKPC-positive isolates from hospital wastewater plumbing, particularly sink drains (23). Sequencing has revealed that several isolates initially identified as K. pneumoniae were further speciated via sequencing as K. quasipneumoniae (8). We have also frequently detected blaKPC-positive S. marcescens in this plumbing reservoir, and a strain that has been seen in patients and the environment within our hospital was included (CAV1492) (24).
These preliminary findings suggest a potential link between environmental persistence, biocide resistance to killing, and antimicrobial resistance. However, the small sample size and the selection from a single institution mean the data should be viewed as very preliminary. Variation within the genus Klebsiella may reflect adaptation to distinct ecological niches and contribute to environmental persistence (25). Consistent with this, species more commonly linked to environmental reservoirs, including K. variicola and K. quasipneumoniae, exhibited higher chlorhexidine MICs. This raises the possibility that biocide exposure in healthcare or environmental settings is selecting for strains with both increased MICs to disinfectants and the ability to carry clinically relevant resistance determinants such as blaKPC driving outbreaks related to premises plumbing (26, 27).
Although the data set is limited in size, it highlights the importance of species-level identification and emphasizes the need to consider both clinical and environmental isolates in infection control strategies. The observed differences in chlorhexidine MICs between species underscore the role of ecological pressures in shaping microbial populations. Co-selection of antimicrobial resistance through biocide exposure may be an underappreciated factor in the emergence and spread of multidrug-resistant organisms in healthcare settings, warranting further investigation into the mechanisms of resistance to biocide killing and their epidemiological consequences.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Murray CJL, Ikuta KS, Sharara F, Swetschinski L, Robles Aguilar G, Gray A, Han C, Bisignano C, Rao P, Wool E, et al.. 2022. Global burden of bacterial antimicrobial resistance in 2019: a systematic analysis. Lancet 399:629–655. doi:10.1016/S 0140-6736(21)02724-035065702 PMC 8841637 · doi ↗ · pubmed ↗
- 2Regev-Yochay G, Margalit I, Smollan G, Rapaport R, Tal I, Hanage WP, Pinas Zade N, Jaber H, Taylor BP, Che Y, Rahav G, Zimlichman E, Keller N. 2024. Sink-traps are a major source for carbapenemase-producing Enterobacteriaceae transmission. Infect Control Hosp Epidemiol 45:284–291. doi:10.1017/ice.2023.27038149351 · doi ↗ · pubmed ↗
- 3Anantharajah A, Goormaghtigh F, Nguvuyla Mantu E, Güler B, Bearzatto B, Momal A, Werion A, Hantson P, Kabamba-Mukadi B, Van Bambeke F, Rodriguez-Villalobos H, Verroken A. 2024. Long-term intensive care unit outbreak of carbapenemase-producing organisms associated with contaminated sink drains. J Hosp Infect 143:38–47. doi:10.1016/j.jhin.2023.10.01038295006 · doi ↗ · pubmed ↗
- 4Long SW, Linson SE, Ojeda Saavedra M, Cantu C, Davis JJ, Brettin T, Olsen RJ. 2017. Whole-Genome sequencing of human clinical Klebsiella pneumoniae isolates reveals misidentification and misunderstandings of Klebsiella pneumoniae, Klebsiella variicola, and Klebsiella quasipneumoniae. m Sphere 2. doi:10.1128/m Sphere Direct.00290-17PMC 554116228776045 · doi ↗ · pubmed ↗
- 5Voellmy IK, Lang C, Gasser M, Kronenberg A, Swiss Centre for Antibiotic Resistance (ANRESIS). 2022. Antibiotic resistance surveillance of Klebsiella pneumoniae complex is affected by refined MALDI-TOF identification, Swiss data, 2017 to 2022. Euro Surveill 27:2200104. doi:10.2807/1560-7917.ES.2022.27.45.220010436367012 PMC 9650708 · doi ↗ · pubmed ↗
- 6Thorpe HA, Booton R, Kallonen T, Gibbon MJ, Couto N, Passet V, López-Fernández S, Rodrigues C, Matthews L, Mitchell S, Reeve R, David S, Merla C, Corbella M, Ferrari C, Comandatore F, Marone P, Brisse S, Sassera D, Corander J, Feil EJ. 2022. A large-scale genomic snapshot of Klebsiella spp. isolates in Northern Italy reveals limited transmission between clinical and non-clinical settings. Nat Microbiol 7:2054–2067. doi:10.1038/s 41564-022-01263-036411354 PMC 9712112 · doi ↗ · pubmed ↗
- 7Zhang Y, Gu D, Yang X, Wu Y, Liu C, Shen Z, Zhang R. 2021. Emergence and genomic characterization of a KPC-2-, NDM-1-, and IMP-4-producing Klebsiella michiganensis isolate. Front Microbiol 12:762509. doi:10.3389/fmicb.2021.76250935069468 PMC 8770907 · doi ↗ · pubmed ↗
- 8Mathers AJ, Crook D, Vaughan A, Barry KE, Vegesana K, Stoesser N, Parikh HI, Sebra R, Kotay S, Walker AS, Sheppard AE. 2019. Klebsiella quasipneumoniae provides a window into carbapenemase gene transfer, plasmid rearrangements, and patient interactions with the hospital environment. Antimicrob Agents Chemother 63:e 02513-18. doi:10.1128/AAC.02513-1830910889 PMC 6535554 · doi ↗ · pubmed ↗