Identification of a Novel, Oncogenic and Targetable TPR::ABL2 Fusion Gene in T‐Cell Acute Lymphoblastic Leukaemia
Elias Lagonik, Elyse. C. Page, Laura N. Eadie, Caitlin E. Schutz, Jacqueline A. Rehn, Susan L. Heatley, Andrew S. Moore, Muirin Healy, Morag Whyte, Timothy P. Hughes, David T. Yeung, Deborah L. White

TL;DR
A new TPR::ABL2 fusion gene was found in a high-risk T-cell leukemia case and shown to be treatable with existing drugs.
Contribution
Discovery of a novel TPR::ABL2 fusion gene in T-ALL and its targetability with tyrosine kinase inhibitors.
Findings
TPR::ABL2 fusion promotes cytokine-independent growth in Ba/F3 cells, showing its oncogenic potential.
TPR::ABL2 exhibits kinase activation and is sensitive to tyrosine kinase inhibitors in both patient and cell models.
The study expands the known ABL2 fusion repertoire in ALL and suggests TKIs as a treatment option.
Abstract
ABL2 rearrangements represent a subtype of acute lymphoblastic leukaemia (ALL) associated with poor prognosis and survival. This study reports a high‐risk T‐cell ALL (T‐ALL) case with a novel TPR::ABL2 gene fusion resulting from a chromosomal deletion. Overexpression of TPR::ABL2 in Ba/F3 cells promoted cytokine‐independent growth, demonstrating its oncogenic nature. Both primary patient and Ba/F3 cells carrying TPR::ABL2 exhibited kinase activation and sensitivity to tyrosine kinase inhibitors (TKIs). This study expands the repertoire of ABL2 fusions identified in ALL and supports the incorporation of TKIs into T‐ALL treatment regimens to improve outcomes for this subtype.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
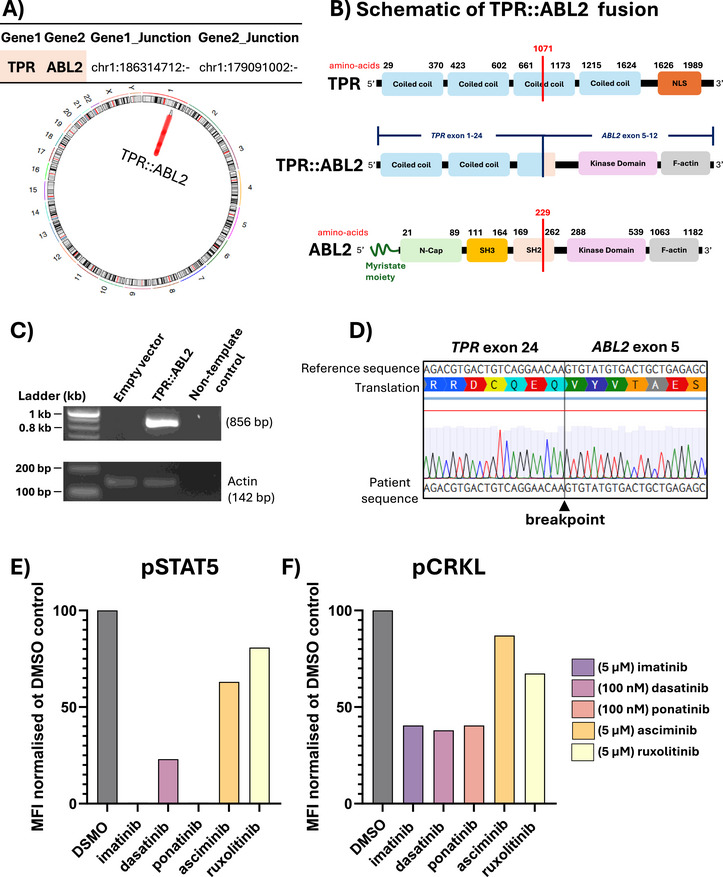 FIGURE 1
FIGURE 1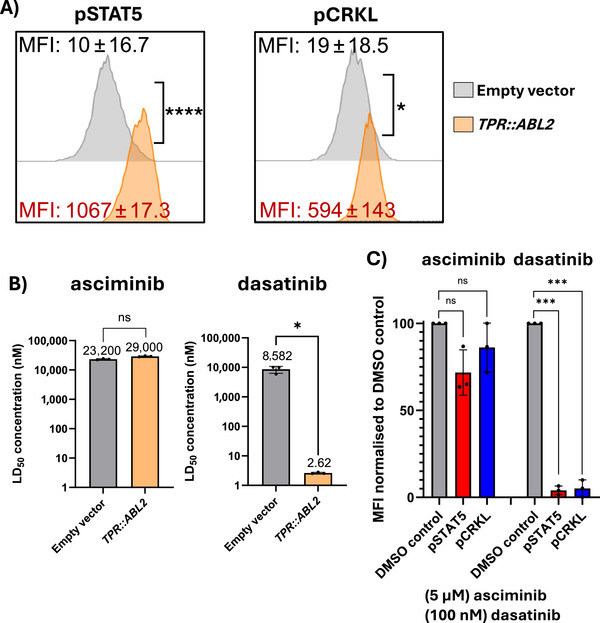 FIGURE 2
FIGURE 2- —National Health and Medical Research Council10.13039/501100000925
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAcute Lymphoblastic Leukemia research · Chronic Myeloid Leukemia Treatments · Carcinogens and Genotoxicity Assessment
Introduction
1
T‐cell acute lymphoblastic leukaemia (T‐ALL) is an aggressive haematological cancer arising from differentiation‐arrested T‐cell precursors. Next‐generation sequencing has revealed the diverse genomic basis of T‐ALL, which comprises multiple subtypes accounting for 15%–20% of ALL cases [1]. T‐ALL is characterised by high relapse rates, poor outcomes and limited clinically approved targeted treatment options [2, 3]. Current treatment regimens consist of intensive multi‐agent chemotherapy followed by haematopoietic stem cell transplantation (HSCT) [3]. Therefore, there is an unmet need to improve therapeutic approaches. In T‐ALL, recurrent genomic alterations often involve ABL1 rearrangements, with the NUP214::ABL1 fusion gene identified in 5%–10% of patients with T‐ALL [3]. The ABL2 gene, a paralogue of ABL1, has also been implicated in T‐ALL, although ABL2 fusion genes are reported less frequently [1, 3, 4]. Similar to ABL1, ABL2 fusion genes result in the loss of the N‐terminal inhibitory elements, leading to the constitutive activation of the ABL kinase and downstream signalling pathways [5, 6]. ABL1 and ABL2 rearrangements (ABL‐r) are particularly interesting due to their susceptibility to inhibition by tyrosine kinase inhibitors (TKIs) [7]. In T‐ALL, ABL‐r often co‐occur with TLX3 enhancer hijacking events, resulting in increased expression of the TLX3 gene [1]. Additional genomic alterations include the activation of signalling pathways (e.g., NOTCH1) and the loss of tumour suppressor pathways (e.g., inactivating mutations in FBXW7 and CDKN2A/B) [1, 8]. In this study, we identify the Translocated Promoter Region (TPR) gene as a fusion partner of ABL2 in a paediatric case of T‐ALL. The leukaemogenic potential of the novel TPR::ABL2 gene and sensitivity to small inhibitors were characterised in vitro and ex vivo.
Methods and Results
2
A retrospective genomic analysis was conducted on a cryopreserved bone marrow (BM) sample from a patient at the Queensland Children's Hospital (Brisbane, QLD). Experiments were performed in accordance with the Women's and Children's Health Network Human Research Ethics Committee. See Supporting Information for detailed methodology protocols.
A 6‐year‐old boy presented with a short history of gastrointestinal symptoms, fever and lethargy. Examination revealed cervical lymphadenopathy, hepatosplenomegaly and ascites. The testicular exam was unremarkable. There was mild mediastinal widening on the chest x‐ray, and peripheral blood leukocytosis (white blood cell count of 467.5 × 10^9^/L) with blasts. BM and peripheral blood investigations confirmed a diagnosis of T‐ALL, subsequently shown by genomic analyses to be driven by the novel TPR::ABL2 fusion gene. There was no CNS involvement. Following induction chemotherapy, residual disease of 8% was demonstrated by flow cytometry. Consolidation was completed as per AALL0434 arm B with nelarabine, and he commenced on dasatinib during this time. End of consolidation testing showed persistent residual disease of 0.08%, so he then received fludarabine, cytarabine, idarubicin (FLA‐IDA) to attain minimal residual disease < 0.01% prior to planned HSCT. He underwent a total body irradiation, cyclophosphamide, thiotepa conditioning and a fully matched unrelated donor peripheral blood HSCT 6 months after initial diagnosis. The procedure was well tolerated with no significant complications, including graft‐versus‐host disease. The patient remains well and in remission, at the time of this report, 2.5 years following the allograft.
Bone marrow mononuclear cells (BMMNCs) were isolated using Lymphoprep density gradient centrifugation. Immunophenotyping of BMMNCs indicated approximately ∼73% lymphoid origin cells. Cells within the lymphoid/blast gate expressed CD3 and CD7 (63%) with negligible expression of CD13, CD33, CD117 (c‐kit) and CD34 (all < 3%) (Figure S1, Table S1). BMMNCs mRNA sequencing revealed a novel cytogenetically cryptic, in‐frame fusion between the 5' end of TPR (Exon 1–24, NM_003292) and the 3' end of ABL2 (Exon 5–13, NM_001168237), resulting from an interstitial deletion on Chr.1q. Breakpoint PCR and Sanger sequencing confirmed the fusion gene, and the complete and partial loss respectively of the ABL2 SH3 and SH2 domain (Figure 1A–D). The novel TPR::ABL2 fusion gene sub‐classifies this case as *ABL‐*rearranged ALL. While this fusion is novel, other TPR fusions have been previously reported in patients with B‐cell and T‐cell ALL [1, 7, 9]. Additional genomic alterations identified in the current patient included the deleterious FBXW7 p.R465C mutation with a variant allele frequency (VAF = 0.29). The FBXW7 protein is a tumour suppressor that regulates NOTCH1 protein activity through direct binding and degradation. The FBXW7 p.R465C mutation results in a loss of function and inability to target NOTCH1 and c‐MYC, a consequence linked to gamma secretase inhibitor resistance [8]. The SALSA Multiplex Ligation‐dependent Probe Amplification (MLPA) assay (#P383, #P202, #P335; MRC‐Holland, Amsterdam, NL) revealed additional high‐risk genomic lesions, including heterozygous deletion of LEF1 (Exon 1–4; Chr4q25) (Figure S2). Deletions of LEF1 first exons are frequent in paediatric T‐ALL, although rare in patients with ABLrearrangements [1]. MLPA also revealed a focal biallelic deletion of CDKN2A (Exon 4; Chr9p21.3) (Figure S2). Deletion of CDKN2A/B is observed in the majority of T‐ALL cases [1]. Interestingly, and in contrast to other ABL‐rearranged T‐ALL cases, increased expression of TLX3 was not noted (data not shown) [1]. BMMNCs expressing TPR::ABL2 demonstrated intrinsically activated phosphorylation of downstream effector proteins, CRKL (pCRKL) and STAT5 (pSTAT5), as measured by intracellular phosphoflow cytometry. TKIs have been used successfully to target ABL2‐r in the clinic [7]. ATP‐competitive TKIs, including imatinib, dasatinib and ponatinib, abrogated pCRKL and pSTAT5 signalling, while asciminib and the JAK inhibitor ruxolitinib had minimal effect (Figure 1E,F) (Table S1).
Identification of a novel TPR::ABL2 fusion gene in T‐cell acute lymphoblastic leukaemia. (A) Circos plot representation of the TPR::ABL2 fusion gene identified by mRNA‐sequencing of BMMNCs at diagnosis. Isoforms TPR (NM_003292) and ABL2 (NM_001168237). (B) Schematic representation of full‐length TPR, full‐length ABL2, and TPR::ABL2 fusion gene. Breakpoints are denoted with a red line. (C) PCR amplification of the TPR::ABL2 breakpoint region. (D) Sanger sequencing confirmation of the TPR::ABL2 breakpoint sequence. (E, F) BMMNCs were incubated for 2 h in either vehicle control (DMSO), TKIs, or ruxolitinib (negative control). Phosphorylation of STAT5 and CRKL was assessed by intracellular flow cytometry, compared to isotype controls IgG1‐PE (pSTAT5) or IgG2a‐PE (pCRKL), and normalised to DMSO vehicle control. MFI, mean fluorescence intensity.
The full‐length TPR::ABL2 fusion gene was isolated from patient material (Table S2) and retrovirally transduced into the Ba/F3 cell line. Expression of TPR::ABL2 resulted in IL‐3‐independent growth, demonstrating constitutive kinase activation and associated phosphorylation of CRKL and STAT5 (Figure 2A). To evaluate the sensitivity of TPR::ABL2 expressing cells to ATP‐competitive TKIs and the allosteric inhibitor asciminib, cell death assays were conducted (Table S1) and demonstrated the efficacy of dasatinib against the oncoprotein in the nanomolar range (Figure 2B). Asciminib failed to induce cell death. Similarly, the constitutive activation of pSTAT5 and pCRKL was diminished only in the presence of dasatinib, confirming the cell death assay results (Figure 2C).
*Ba/F3 cells expressing the TPR::ABL2 fusion gene exhibit constitutive kinase activation and are sensitive to ATP‐competitive TKIs. (A) Cells were assessed for STAT5 and CRKL phosphorylation. Empty vector Ba/F3 cells were cultured in the absence of IL‐3 for 5 h. MFI values of three independent experiments are highlighted. Histograms are representative of n = 3 biological replicates. (B) Ba/F3 cells expressing TPR::ABL2 were incubated for 72 h with vehicle control (DMSO) or increasing concentrations of dasatinib or asciminib. The mean LD50 nanomolar concentration for each cell line is denoted above the respective columns. Columns represent mean ± SD. (C) Ba/F3 cells expressing the TPR::ABL2 were incubated for 2 h in DMSO or 100 nM dasatinib, or 5 µM asciminib. STAT5 and CRKL phosphorylation were assessed by intracellular flow cytometry. Statistical significance was determined by unpaired Student's t‐test with Welch's correction (ns =not significant, *p < 0.05, **p < 0.01, ***p < 0.001, ***p < 0.0001). MFI, mean fluorescence intensity.
Discussion
3
In this study, we identified the novel TPR::ABL2 fusion gene by mRNA and Sanger sequencing and demonstrated its oncogenicity in vitro and susceptibility to ATP‐competitive TKIs currently in clinical use. Few reports have described fusion genes involving TPR in malignancies. The TPR::MET fusion gene is commonly found in gastric carcinoma, while other TPR fusion genes have been identified in paediatric patients with ALL and other malignancies [1, 7, 9, 10, 11]. Our findings suggest that haematological malignancies harbouring ABL2‐r involving the TPR gene may be amenable to targeted therapies, such as TKIs.
TPR is associated with the nuclear pore complex (NPC), encoding a structural component of the nuclear basket, which mediates macromolecule and mRNA export from the nucleus to the cytoplasm. While fusions involving the nucleoporin genes NUP214 and NUP98 have been well‐documented in haematological cancers, TPR alterations remain poorly characterised [10, 12]. TPR features a N‐terminal coiled‐coil domain containing heptad repeats and leucine zipper motifs. The coiled‐coil domain mediates the organisation of the protein into tetramers with two parallel dimers associated in an antiparallel orientation. The leucine zippers stabilise the tetrameric TPR structure by forming a hydrophobic core. The C‐terminal contains a nuclear localisation signal (NLS) [10, 12]. In the TPR::ABL2 fusion gene identified in this study, the breakpoint resulted in the retention of the TPR N‐terminal coiled‐coil domains. Similar to BCR::ABL1, the oncogenic activity of TPR::ABL2 likely results from the loss of the myristate moiety, an auto‐inhibitory element of ABL2 [5, 6]. Furthermore, the N‐terminal coiled‐coil domain of TPR may facilitate the oligomerisation of the chimeric protein and subsequent transphosphorylation of the ABL2 kinase domain, leading to the constitutive activation of signalling pathways. In BCR::ABL1, the oligomerisation of the chimeric protein is critical for its oncogenic activity [5]. In accordance with previous studies, primary BMMNCs and Ba/F3 cells expressing TPR::ABL2 were sensitive to ATP‐competitive TKIs [7]. In addition, the ABL2 breakpoint of the TPR::ABL2 fusion gene is located at Exon 5, and the SH3 domain critical for mediating asciminib efficacy is lost. Absence of a functional SH3 domain abrogated the inhibitory effect of the allosteric inhibitor asciminib on the TPR::ABL2 oncoprotein, confirming our previous findings [7, 13, 14, 15].
Patient response to dasatinib prior to HSCT is suggestive of an effective targeted therapy for the novel TPR::ABL2 fusion gene. Furthermore, the TPR::ABL2 was sensitive to multiple ATP‐competitive TKIs both in patient cells and in an in vitro cell line model. The evidence presented here supports the incorporation of TKIs into the treatment strategies for patients with *ABL2‐*rearranged ALL, improving clinical management.
Author Contributions
E.L. designed the study, performed experiments, analysed the data, wrote the manuscript and created the figures. E.C.P., L.N.E., C.S., J.A.R., and S.L.H. performed experiments. A.S.M., M.H., M.W., D.T.Y., T.P.H., and D.L.W. provided scientific and clinical insight. All authors critically reviewed the manuscript.
Funding
This study was undertaken with the financial support of a National Health and Medical Research Council APP2007908 and an MRFF2007441 grant. Elias Lagonik was supported by a University of Adelaide Research Scholarship. Elyse. C. Page, Laura N. Eadie and David T. Yeung were supported with fellowship funding from the Cancer Council SA's Beat Cancer Project on behalf of its donors and the State Government through the Department of Health.
Conflicts of Interest
David T. Yeung receives research support from BMS and Novartis and Honoraria from BMS, Novartis, Pfizer, Ascentage and Amgen. Timothy P. Hughes receives research support from BMS and Novartis and Honoraria from BMS, Novartis and Fusion Pharma. Deborah L. White receives research support from BMS and Honoraria from BMS and AMGEN. The other authors declare no conflicts of interest.
Supporting information
Supporting Figure 1: Immunophenotypic profile of TPR::ABL2 T‐ALL patient bone marrow mononuclear cells (BMMNCs). A) Immunophenotyping of the primary patient sample revealed that the blast population was approximately ‐73.4%. Leukaemic/blast cells B‐F) were 63% CD3 +/CD7 +, 9% CD3 +/CD7‐ and 20% CD3 −/CD7 + with negligible expression of CD34 +, CD13+, CD33+ and CD117+ (c‐kit). Supplemental Figure 2: Multiplex Ligation‐dependent Probe amplification analysis. The SALSA MLPA Probemix #P202, #P335, and #P383 T‐ALL oligonucleotide probes were utilised. MLPA analysis on genomic DNA revealed a heterozygous deletion of LEF1 (exon 1–4) and a homozygous deletion of CDKN2A (exon 4). Deletions are highlighted in red. Supplemental Table 1: Antibodies List. Supplemental Table 2: Materials.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1P. Pölönen , D. Di Giacomo , A. E. Seffernick , et al., “The Genomic Basis of Childhood T‐Lineage Acute Lymphoblastic Leukaemia,” Nature 632, no. 8027 (2024): 1082–1091.39143224 10.1038/s 41586-024-07807-0PMC 11611067 · doi ↗ · pubmed ↗
- 2T. Terwilliger and M. Abdul‐Hay , “Acute Lymphoblastic Leukemia: A Comprehensive Review and 2017 Update,” Blood Cancer Journal 7, no. 6 (2017): e 577–e 577, 10.1038/bcj.2017.53.28665419 PMC 5520400 · doi ↗ · pubmed ↗
- 3T. Steimlé , M.‐E. Dourthe , M. Alcantara , et al., “Clinico‐Biological Features of T‐Cell Acute Lymphoblastic Leukemia With Fusion Proteins,” Blood Cancer Journal 12, no. 1 (2022): 14, 10.1038/s 41408-022-00613-9.35082269 PMC 8791998 · doi ↗ · pubmed ↗
- 4B. Chen , L. Jiang , M.‐L. Zhong , et al., “Identification of Fusion Genes and Characterization of Transcriptome Features in T‐Cell Acute Lymphoblastic Leukemia,” Proceedings of the National Academy of Sciences 115, no. 2 (2018): 373–378, 10.1073/pnas.1717125115.PMC 577707029279377 · doi ↗ · pubmed ↗
- 5O. Hantschel , B. Nagar , S. Guettler , et al., “A Myristoyl/Phosphotyrosine Switch Regulates C‐ABL,” Cell 112, no. 6 (2003): 845–857, 10.1016/S 0092-8674(03)00191-0.12654250 · doi ↗ · pubmed ↗
- 6J. Colicelli , “ABL Tyrosine Kinases: Evolution of Function, Regulation, and Specificity,” Science Signaling 3, no. 139 (2010): re 6, 10.1126/scisignal.3139 re 6.20841568 PMC 2954126 · doi ↗ · pubmed ↗
- 7K. G. Roberts , Y. Li , D. Payne‐Turner , et al., “Targetable Kinase‐Activating Lesions in Ph‐Like Acute Lymphoblastic Leukemia,” New England Journal of Medicine 371, no. 11 (2014): 1005–1015, 10.1056/NEJ Moa 1403088.25207766 PMC 4191900 · doi ↗ · pubmed ↗
- 8J. Fan , M. Bellon , M. Ju , et al., “Clinical Significance of FBXW 7 Loss of Function in Human Cancers,” Molecular Cancer 21, no. 1 (2022): 87, 10.1186/s 12943-022-01548-2.35346215 PMC 8962602 · doi ↗ · pubmed ↗