Fine-tuning RIPENING INHIBITOR (RIN) expression by introducing allelic mutations in its promoter using CRISPR/Cas9 multiplex editing
Jiaqi Zhou, Chiu-Ling Yang, Diane M. Beckles

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
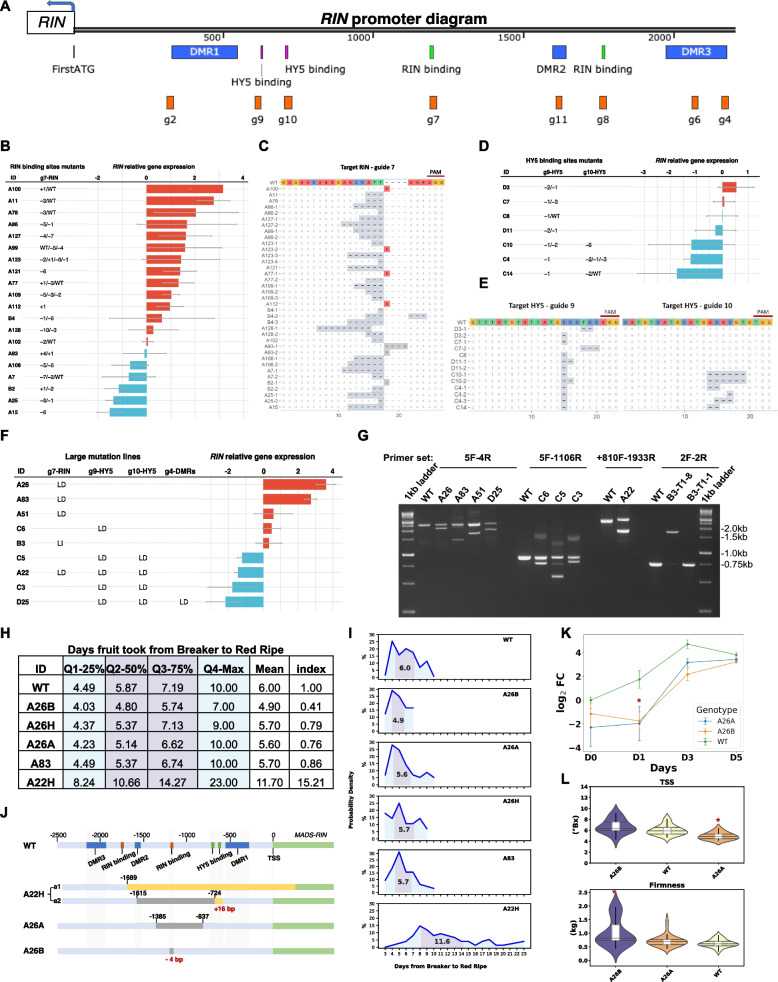 Figure 1
Figure 1- —http://dx.doi.org/10.13039/100022967College of Agricultural and Environmental Sciences, University of California, Davis
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCRISPR and Genetic Engineering · Genetic Neurodegenerative Diseases · interferon and immune responses
Tomato is a valuable vegetable crop but is perishable, and thus has a limited shelf-life. It is also an important functional genomics model for studying the regulation of fleshy fruit ripening, which is complex. Ripening is governed by many transcriptional factors (TFs) which orchestrate the expression of other TFs and downstream genes as part of multiple, but highly integrated ripening regulatory networks (Tonutti et al. 2023). The MADS-box RIPENING INHIBITOR (RIN) directly binds to many downstream TFs and genes, turning them on/off during fruit ripening (Fujisawa et al. 2013). The naturally occurring tomato rin mutant, was discovered in the 1900s, and its allele has been incorporated into tomato breeding programs to extend fruit shelf-life because it slows ripening. Recent studies revealed that rin is a gain-of-function mutation, and a CRISPR/Cas9 null mutant Rin was capable of initiating fruit ripening but disrupted ripening phenotypes (Ito et al. 2017; Li et al. 2020a, b; Vrebalov et al. 2002). Due to severe disruptions in RIN expression, many of the desirable quality traits associated with fruit ripening are destroyed (Ito et al. 2017; Li et al. 2020a, b; Vrebalov et al. 2002).
Editing gene regulatory regions instead of coding regions may provide an opportunity to fine-tune gene expression, therefore leading to a range of subtle phenotypic changes (Albornoz et al. 2022; Huang et al. 2020). This work focuses on RIN transcriptional regulation. RIN gene promoter regions, upstream of the transcription start site, contain a series of Cis-Regulatory Elements (CREs), i.e., specific DNA sequence motifs to which TFs bind, to drive or suppress RIN gene expression (Schwarzer & Spitz 2014). CRE mutations can destroy TF binding sites, introduce additional copies of a CRE, or create a novel CRE to allow new TFs to bind, thereby altering the transcriptional levels of its cognate gene (Schwarzer & Spitz 2014). Analysis of the RIN promoter (Fig. 1A) indicates that there are (a) two CArG motifs to which RIN binds (Bemer et al. 2012; Fujisawa et al. 2013), (b) two G-box motifs that the TF Elongated Hypocotyl 5 (HY5) binds to during tomato fruit ripening (Wang et al. 2021), and, (c) numerous putative CREs to which diverse TFs may bind in response to changes in developmental stages, environmental conditions, and hormones (Lescot et al. 2002). The CArG motifs are of interest because RIN protein binding may create an autoregulatory loop that intensifies the regulation of RIN gene transcriptional levels, with consequences for the expression of RIN-targeted downstream TFs and genes.Fig. 1RIN promoter allelic mutants using CRISPR/Cas9. A Analysis of the RIN promoter, arbitrarily defined as the 2–3 kb upstream of the transcriptional start site. There are three DMRs, targeted by gRNAs—g2, g6, and g4, respectively; two HY5 binding sites targeted by g9 and g10, and two RIN binding sites targeted by g7 and g8. The gRNA sequences are in Table S1. A summary of the mutations, their location, types, and levels are as follows: B and C a RIN binding site, D and E HY5 binding sites, F and G large mutation, i.e., large insertion (LI) or large deletion (LD) at a single RIN binding site, two HY5 binding sites, or DMRs. RIN relative gene expression in tomato fruit at red ripe was normalized to the reference gene SlCAC, using WT red-ripe fruit as the control, and primers used are in Table S4. The log_2_ fold change was presented as the bar in (B, D, F). The red bars show RIN upregulation- log_2_ fold change > 0, and the blue bars show RIN downregulation relative to the WT- log_2_ fold change < 0. The sequence of the mutated alleles for (B) and (D) is shown as (C) and (E). In C and E the color of the nucleotide backgrounds indicates mutation types; white indicates no change compared to the WT, grey shows deletions, and red regions indicate insertions. There are multiple alleles in some mutants, which are presented in individual rows. G A 1% (w/v) agarose gel image showing one or multiple alleles in the large mutation lines, using WT as the control. The primer pairs used to amplify the bands are indicated 5F-4R etc. (details are in Table S3). H Quartile statistic and index of ripening days in selected T_1_ (i.e., the second-generation) edited lines. Data shows the number of days fruit took to go from Breaker to Red Ripe when ripened on-the-vine. Five mutants and the WT are included. A ripening index for each genotype was computed by multiplying all values from Q1-Q4 and dividing the result by the corresponding value in the WT. I Trends of fruit on-the-vine ripening over time based on data generated from (H). The x-axis represents the number of days from Breaker to Red Ripe, and the y-axis shows the probability density, indicating the proportion of fruit within the population. The purple shaded area highlights the interquartile range (Q2 to Q3), and the mean number of days is marked within this range. J Genotypes ‘A22H’, ‘A26A’ and ‘A26B’ showing the locations of introduced mutations in the RIN gene promoter. K RIN relative gene expression during on-the-vine ripening. The y-axis shows log_2_ fold change of RIN, using the WT at Breaker stage (D0) as the control. The x-axis represents days post-breaker (0, 1, 3, 5 days). L Fruit quality traits, including total soluble solids (TSS), and firmness (kg). Fruit were harvested at the breaker stage and stored at 20 °C for one week. Dunnett’s pairwise comparison was performed using WT as the control. Asterisks ‘*’ indicate statistically significant differences (p < 0.05)
RIN binding sites in the gene promoter are frequently close to differentially methylated regions (DMRs), with RIN binding usually occurring at hypomethylation sites (Zhong et al. 2013). DMRs vary in DNA methylation status due to developmental or environmental cues, and may influence transcription by modulating the chromatin accessibility of nearby TFs (Henderson & Jacobsen 2007). In the RIN promoter, DMRs have been identified that are hypermethylated in immature green tomato fruit but then, are demethylated in ripe fruit (Lang et al. 2017). Global DNA demethylation is crucial for tomato fruit ripening and for RIN binding its downstream target TFs and genes. The tomato dml2-3 mutant, which lacks a key DNA demethylase, shows genome-wide hypermethylation and disrupted fruit ripening (Niu et al. 2025). Ectopic expression of RIN in the dml2-3 background partially restores the ripening phenotype, suggesting that RIN may mediate DML2-related ripening regulation (Niu et al. 2025). However, it remains unclear whether changes in DMRs within the RIN promoter, rather than genome-wide methylation shifts, are sufficient to influence RIN transcription and fruit ripening.
Evidence suggests that editing multiple CREs in the promoter of key regulatory genes in tomato, can lead to subtle alterations in the mRNA levels of the edited gene (Rodríguez-Leal et al. 2017). Therefore, differentially editing the RIN promoter using multiplex CRISPR/Cas9 could fine-tune RIN mRNA and protein levels. In addition to the known TF binding sites, recent findings reveal that SlWOX13, a member of the WOX family and a newly identified ripening enhancer, also binds the RIN promoter adjacent to the HY5 binding site (Jiang et al. 2023). This finding illustrates the complexity of the RIN regulatory landscape, where multiple TFs converge on the RIN promoter to coordinate its transcription during ripening. Therefore, subtle changes in RIN gene promoter sequence, could lead to a cascade of nuanced alterations in ripening-related gene expression patterns that would affect ripening rates and fruit quality. Taken together, this work is expected to answer whether mutations in the RIN gene promoter can affect its transcription and create novel ripening phenotypes.
We designed eight 20 bp-gRNAs spanning the RIN promoter, targeting each regulatory site (Fig. 1A). There were four sets of gRNAs combinations assembled, to target regulatory regions, individually or concurrently, i.e., (a) two gRNAs targeting the identified RIN self-binding sites (the CArG motif) (Bemer et al. 2012)—g7 and g8, (b) two gRNAs targeting the HY5 binding (Wang et al. 2021) sites—g9 and g10, (c) four gRNAs on the DMRs (Fig. S1) (Lang et al. 2017)—g2, g11, g4, and g6, and (d) six gRNAs targeting the RIN and HY5 binding sites and the DMRs together—g2, g9, g10, g7, g8, and g4 (Fig. S2). Four CRISPR/Cas9 constructs were assembled and introduced into tomato cotyledons using Agrobacterium-mediated plant transformation. We generated 264 independent first-generation (T_0_) transgenic lines. The genotyping results indicated that 24 lines had large insertions or deletions within the RIN promoter, ranging from hundreds to thousands of base pairs, sometimes spanning multiple CREs. Many lines had small indels at one or multiple sites; for mutants with small indels at a single site, 51 and 29 were at the RIN and HY5 binding sites, and 14 were at the DMRs. Only four mutants had small indels at both the HY5 sites and DMRs. The sequencing results showed that most T_0_ lines were heterozygous, containing at least one wild-type (WT) allele, or exhibited biallelic or chimeric patterns in the targeted regions. Only a few lines, such as ‘A121’, ‘A112’, ‘A15’, and ‘C14’, had homozygous alleles (Fig. 1B-E, S3).
To evaluate the effect of these promoter mutations on RIN transcription during fruit ripening, the relative expression of RIN in mutant fruit was compared to control fruit (WT) using RT-qPCR. Tissue culture plantlets were transferred into a greenhouse and T_0_ lines were selected for fruit tissue sampling. Only red-ripe fruit was assessed, as RIN expression is stable at this developmental timeframe in WT fruit (Li et al. 2020a, b; Shinozaki et al. 2018). We observed a range of RIN expression levels among the T_0_ mutants, and ranked the lines accordingly based on their RIN expression (Fig. 1).
RIN binding sites
T_0_ lines with mutations at the RIN CRE by g7 (Fig. 1B) had both positive and negative effects on RIN expression. However, no edits were associated with g8 when over a hundred lines were screened. This is possibly due to g8’s low editing efficiency or limited chromatin accessibility at its target site. RIN expression in ‘A100’ was 8.9-fold higher, while in ‘A15’, it was 2.9-fold lower than that in the WT. By examining the mutation location and expression levels (Fig. 1B, C), the data suggested (i) a single adenine insertion may underlie the higher RIN expression observed in ‘A100’, ‘A123’, ‘A77’ and ‘A112’, (ii) both ‘A11’ and ‘A78’ had a 3-bp deletion and are heterozygous to WT, with an upregulation of RIN, (iii) ‘A15’ and ‘A121’ had contradictory RIN expression pattern but the same type of promoter mutation; somaclonal variation, induced by tissue culture, may differentially influence RIN expression (Bairu et al. 2011).
HY5 binding sites
Simultaneous mutations at both HY5 sites by g9 and g10 led to a downregulation of RIN expression (‘C10’, ‘C4’, ‘C14’). Four of the seven lines were reduced in RIN expression, one was unchanged, and the other two lines were slightly upregulated (Fig. 1D). It is of note that the location at which the editing occurs is important to transcription; an inverse expression pattern was observed in ‘D3’ and ‘D11’ – these individuals are heterozygous for a 2-bp and a 1-bp deletion (Fig. 1D), but the deletions are at different locations within the g9 target region (Fig. 1E). This variation potentially leads to divergent RIN expression (Fig. 1D).
Large mutations
Two heterozygous large deletions T_0_ lines, ‘A26’ and ‘A83’, had upregulated RIN expression (Fig. 1F, G). ‘A26’ is a biallelic mutant with different deletions at a RIN binding site. The ‘A26’ T_1_ (i.e., the second-generation) lines included the heterozygote ‘A26H’, and two homozygous genotypes, i.e., ‘A26B’ with a 4-bp deletion, and ‘A26A’ with a 321-bp deletion (Fig. 1J). However, the three ‘A26’ T_1_ genotypes did not show the same pattern as their T_0_ RIN expression (Fig. S4). Other lines with large mutations spanning multiple CREs and DMRs all had suppressed RIN, i.e., ‘C5’, ‘A22’, ‘C3’, and ‘D25’; however, there was no correlation between the length of the deletion, and RIN expression. Notably, ‘A22’ is biallelic, where one allele has an 891-bp deletion and a 16-bp insertion in the RIN gene promoter, and the other allele has an insertion in a RIN exon. Both the T_0_ ‘A22’ and its heterozygote T_1_, ‘A22H’, which share the same genotype, had reduced RIN expression (Fig. 1F, S4).
Fruit ripening phenotype
To assess if inducing mutations in RIN CREs affected fruit ripening, five mutant T_1_ lines were selected to evaluate fruit ripening speed, defined as the timeframe from the onset of ripening (Breaker stage) to Red Ripe. Multiple parameters were used to depict the dynamism of fruit ripening within each genotype ripening trends, including quartiles, mean values, i.e., average number of days fruit took to ripen, and a ripening index relative to WT (Fig. 1H). Strikingly, the ‘A22H’ mutant took an average of 11.7 days to ripen compared to 6 days in the WT, with a 15-fold higher ripening index. This ripening delay may be related to an allelic disruption in the RIN exon (Fig. 1J), which is also associated with reduced RIN expression (P < 0.01; Fig. S4). In contrast, other genotypes such as the small indel mutant ‘A26B’, the large deletion mutant ‘A26A’, and the heterozygous lines ‘A26H’ and ‘A83’, had ripening pattern and RIN expression at Red Ripe stage that were comparable to WT (P > 0.05; Fig. 1H, I, S4).
Fruit color was used as a visual marker for ripening progression; however, external color alone may not capture earlier molecular or physiological changes (Zhou et al. 2024). We further investigated whether promoter mutations affect fruit ripening and quality. Two homozygous mutants, ‘A26A’ and ‘A26B’, each carrying mutations at RIN binding site (Fig. 1J), were selected for further analysis. RIN gene expression was suppressed early, at day 1 (P < 0.05) and day 3 post-breaker but returned to WT-like levels by day 5 (Fig. 1K) in both genotypes. Consistent with the gene expression pattern, both genotypes displayed a ‘delayed ripening’ phenotype (Fig. 1L). Assessment of two common fruit quality traits, total soluble solids (TSS) and firmness, revealed that ‘A26B’ fruit had significantly higher firmness (P < 0.05), while ‘A26A’ fruit had reduced TSS contents (P < 0.05). Together, these findings suggest that disruption of the RIN promoter can alter RIN transcription dynamics during early ripening and affect fruit quality traits, even without fully delaying the overall ripening timeline.
In conclusion, we successfully generated various alleles, i.e., small indels, and large structural changes in the RIN gene promoter, resulting in diverse phenotypic outcomes. These include genotypes with extended ripening times (e.g., ‘A22H’), and others with subtle quality traits changes despite a normal ripening progression. Our work reveals a non-linear relationship between promoter mutations and gene expression levels, with certain types of mutations having a more significant effect than others. This study provides evidence that editing gene regulatory regions can create novel allelic resources varying incrementally in the expression of a ripening gene and alter fruit ripening physiology. Future work will focus on evaluating fruit traits from the mutants with differential RIN expression changes or distinct mutation types, as well as understanding if the mutant fruit will respond to postharvest stress differently.
Supplementary Information
Supplementary Material 1.