From Basics to Breakthroughs: A Review on the Evolution of Campylobacter spp. Culture Media
Ana Rita Barata, Maria José Saavedra, Gonçalo Almeida

TL;DR
This paper reviews the development of culture media for Campylobacter bacteria, showing how methods have evolved from basic to advanced techniques over decades.
Contribution
The paper provides a detailed chronological review of Campylobacter culture media evolution, emphasizing scientific milestones and innovations.
Findings
Early Campylobacter culture media relied on blood and charcoal, but modern formulations are selective and chromogenic.
Advancements in media design have improved detection and isolation of Campylobacter in clinical and food settings.
Integration of molecular insights has led to more precise and standardized culture methods.
Abstract
Since their recognition as human pathogens in the 1970s, Campylobacter spp. have posed persistent challenges to microbiologists due to their fastidious growth requirements and environmental sensitivity. The continuous refinement of selective and differential culture media has been crucial for improving their detection, isolation, and characterization in both clinical and food microbiology. This comprehensive review provides a chronological overview of the evolution of Campylobacter culture media, highlighting the scientific milestones that shaped current cultivation practices—from early blood- and charcoal-based formulations to modern selective, chromogenic, and systems permitting incubation under less stringent atmospheric conditions. Emphasis is placed on the rationale behind medium composition, the transition from empirical experimentation to standardized formulations, and the…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1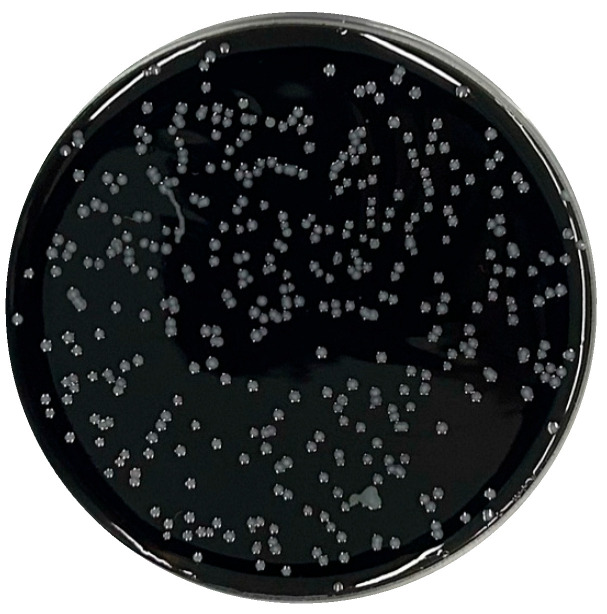 Figure 2
Figure 2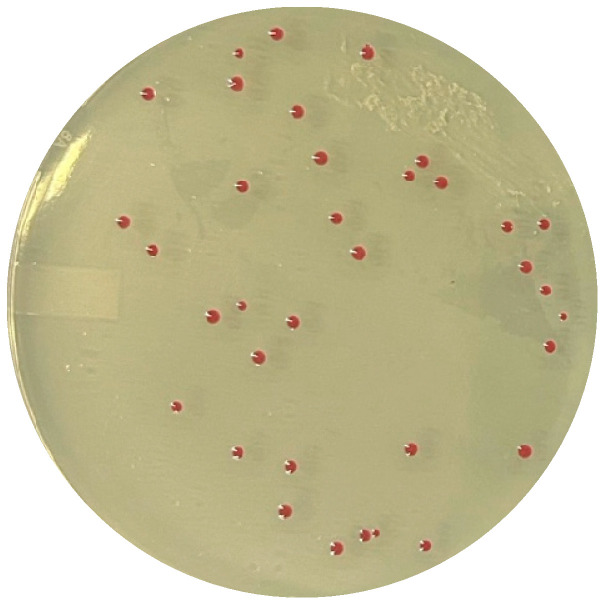 Figure 3
Figure 3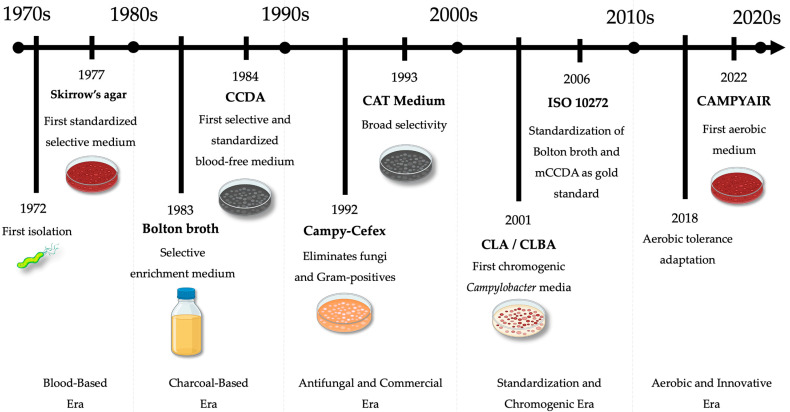 Figure 4
Figure 4| Name | Basal Medium (Per Litter) | Supplements (Per Litter) | Refs. | |
|---|---|---|---|---|
| Broth | Preston broth | Nutrient broth No. 2: peptone (10 g); meat extract (10 g); sodium chloride (5 g) | Actidione (0.0001 g) | [ |
| - | Brucella broth | Cycloheximide (0.05 g) | [ | |
| Agar | Skirrow medium | Blood agar base: peptones (10 g); meat extract (5.0 g); sodium chloride (5.0 g); agar (12.0–15.0 g) | Lysed horse blood (50 g) | [ |
| Preston agar | Same formulation as in Preston broth | Same supplements as in Preston broth | [ | |
| Campy-BAP | Brucella agar base: peptones (10 g); casein enzymic hydrolysate (10 g); yeast extract (2.0 g); dextrose (1.0 g); sodium chloride (5.0 g); agar (12.0–15.0 g) | Amphotericin B (2000 g) | [ | |
| - | Brucella agar base | FBP supplement (0.25 g) | [ | |
| modified Skirrow medium | Agar base | Bacitracin (25 IU) | [ | |
| Campy-thio | Same formulation as in Butzler medium | Amphotericin B (0.02 g) | [ | |
| modified Butzler (BU40) | FTM-A ( | Actidione (0.5 g) | [ |
| Name | Basal Medium | Supplements (Per Liter) | Refs. | |
|---|---|---|---|---|
| Broth | Bolton broth | α-ketoglutaric acid (1.0 g) | Amphotericin B (0.01 g) | [ |
| Tz-Bolton broth | Same formulation as in Bolton broth | Same supplements as in Bolton broth | [ | |
| - | Nutrient broth No. 2 | FBP supplement (0.25 g) | [ | |
| Food Pathogen Enrichment broth (FPE) | Dipotassium phosphate (0.8 g) | Polymyxin B (0.0005 g) | [ | |
| R-Bolton broth | Same formulation as in Bolton broth | Same supplements as in Bolton broth | [ | |
| C-Bolton broth | Same formulation as in Bolton broth | Same supplements as in Bolton broth | [ | |
| Agar | Campy-Line agar | Brucella agar (43.0 g) | 2,3,5-triphenyltetrazolium chloride solution (TCC) (200 ppm) | [ |
| Campy-Line Blood agar | Brucella agar (43.0 g) | Lysed horse blood (50 g) | [ | |
| modified | Brucella agar (44.0 g) | Amphotericin B (0.002 g) | [ | |
| AAV medium | Amphotericin (0.01 g) | [ | ||
| mCCDA | Same formulation as in mCCDA ( | Same supplements as in mCCDA ( | [ | |
| CampyFood Agar (CAMPY agar) | Brucella agar base (43.0 g) | Amphotericin B (0.002 g) | ||
| P-mCCDA | Same formulation as in mCCDA ( | Same supplements as in mCCDA ( | [ | |
| C-mCCDA | Same formulation as in mCCDA ( | Same formulation as in mCCDA ( | [ | |
| Modified Karmali | Same formulation as in Karmali ( | Same formulation as in Karmali ( | [ | |
| BRS agar | Nutrient Broth No. 2 (Oxoid) | Rifampicin (0.010 g) | [ | |
| CM-HT | Agar (15.0 g) | Amphotericin (0.002 g) | [ | |
| CHROMagar™ | Agar (15.0 g) | Chromogenic and selective mix containing antibiotics (2.2 g) | ||
| Same formulation as in mCCDA | Same supplements as in mCCDA | [ | ||
| CAMPYAIR | Beef extract (50.0 g) | Defibrinated sheep blood (100 g) | [ | |
| A-mCCDA | Same formulation as in mCCDA | Same supplements as in mCCDA | [ |
- —FCT—Portuguese Foundation for Science and Technology
- —Centre for the Research and Technology of Agro-Environmental and Biological Sciences
- —LA/P/0126/2020
- —Center for Animal Science Studies (CECA-ICETA)
- —Associated Laboratory of Animal and Veterinary Science (AL4AnimalS)
- —University of Porto
- —The Veterinary and Animal Research Centre (CECAV)
- —AL4AnimalS
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsSalmonella and Campylobacter epidemiology · Probiotics and Fermented Foods · Vibrio bacteria research studies
1. Introduction
Campylobacter spp., particularly Campylobacter jejuni and Campylobacter coli, are among the leading causes of bacterial gastroenteritis worldwide, representing a major concern for public health and food safety [1]. Campylobacteriosis has been the most frequently reported zoonotic disease in the European Union since 2005, consistently surpassing other zoonoses such as salmonellosis, while also being recognized as one of the leading causes of bacterial gastroenteritis worldwide, with a substantial disease burden reported in North America, Asia, Oceania, and parts of Africa [2,3,4]. Its widespread prevalence is largely associated with poultry consumption and foodborne transmission, reinforcing the importance of Campylobacter as a major public health and food safety concern. This trend continues to be confirmed by the most recent European Food Safety Authority (EFSA) report, which identifies Campylobacter as the leading zoonotic agent in 2024 [5]. The ability to accurately detect, isolate, and study these microorganisms is pivotal for understanding their epidemiology, pathogenesis, and antibiotic resistance patterns [6]. Central to these efforts is the development and refinement of culture-based methodologies, specifically the growth media designed to support the unique physiological requirements of Campylobacter species [7]. C. jejuni and C. coli are responsible for the vast majority of reported campylobacteriosis cases, and culture-based detection methods have therefore been historically developed with a strong focus on these thermotolerant species [8]. In the early stages of Campylobacter diagnostics, incubation at 37 °C was commonly applied, largely by analogy with other enteric pathogens and reflecting the recent recognition of Campylobacter as a human pathogen [9]. Subsequent studies in the late 1970s and early 1980s demonstrated that C. jejuni and C. coli grow optimally at approximately 42 °C, a temperature that mirrors the intestinal environment of birds, their primary reservoir [10,11]. The transition to incubation at 42 °C improved recovery and selectivity by enhancing growth of these species while suppressing competing flora, leading to its adoption in selective media and standardized methods [12]. However, this optimization also introduced a methodological bias, as non-thermotolerant Campylobacter species, such as Campylobacter lari and C.ampylobacter upsaliensis, are less efficiently recovered under these conditions and may therefore be under-detected.
Historically, the isolation and cultivation of Campylobacter spp. presented a considerable challenge to microbiologists, attributed to their intricate growth requirements and the lack of suitable culture media. Early efforts to culture these organisms often resulted in low recovery rates and poor growth. However, the ensuing decades have witnessed significant advancements in the formulation of growth media, driven by a deeper understanding of the microbial physiology of Campylobacter and the innovative application of molecular biology techniques [13,14].
This review article aims to trace the evolution of growth media for Campylobacter spp., from the initial attempts at isolation to the sophisticated, selective media available today. By examining the chronological development and scientific rationale behind various media formulations, we seek to highlight the milestones in the cultivation of these important pathogens. Through a comprehensive analysis of the literature, this article will identify emerging trends in media development and suggest future directions for research. In doing so, we aim to provide a valuable resource for microbiologists, food safety experts, and researchers dedicated to the study of Campylobacter spp., facilitating further innovation in this critical area of microbiology.
2. The Genus Campylobacter
Campylobacter comprises a diverse group of Gram-negative, curved bacteria within the family Campylobacteraceae, including more than 30 recognized species. Campylobacter jejuni and C. coli account for most human infections, whereas species such as C. lari, C. upsaliensis, Campylobacter fetus, and Campylobacter hepaticus are mainly associated with animal hosts but retain zoonotic relevance [15]. Variation in host adaptation, temperature preference, and environmental tolerance contributes to differences in pathogenic potential and recovery efficiency during laboratory culture [2].
These bacteria are metabolically specialized, chemoorganotrophic organisms that are generally nonproteolytic, nonlipolytic, and nonsaccharolytic. They derive energy primarily from the oxidation of amino acids and tricarboxylic acid cycle intermediates rather than from carbohydrate fermentation or oxidation. Their fastidious physiology requires reduced oxygen tension and elevated carbon dioxide concentrations for optimal growth. Many pathogenic species are thermotolerant, reflecting adaptation to avian reservoirs, and exhibit marked sensitivity to oxidative stress and inhibitory compounds, necessitating protective growth environments—an important factor that has strongly influenced the formulation of selective culture media [16].
Ecologically, Campylobacter species frequently inhabit the gastrointestinal tract of a broad range of hosts, including poultry, livestock, companion animals, and wildlife, where colonization is often asymptomatic. Their widespread presence in animal reservoirs and contaminated food underscores their zoonotic transmission potential and central role in foodborne disease [17]. From a clinical and ecological perspective, species within the genus can be broadly grouped into thermotolerant strains of primary public health importance, livestock-associated species that infrequently cause human disease, taxa linked to specific human conditions such as periodontal infections, and environmental or host-associated species not currently implicated in foodborne illness. This diversity reflects the adaptive capacity of the genus and contributes to variability in host range, virulence traits, and epidemiological significance [18].
At the genomic level, Campylobacter displays considerable plasticity that supports adaptation to diverse ecological niches and influences virulence, antimicrobial resistance, and metabolic capacity. Motility, adhesion, and toxin-associated functions are tightly regulated in response to environmental cues, shaping bacterial behavior during host colonization and laboratory cultivation [19]. Collectively, these biological traits underpin the demanding growth requirements of Campylobacter spp. and provide essential context for understanding the development of selective media designed to balance organism recovery with suppression of competing microbiota.
3. Literature Overview
This review provides an integrative and chronological overview of the development of culture media for Campylobacter spp. from the late 1960s to the present day. Relevant literature was identified primarily through searches in scientific databases such as PubMed, Scopus, and Web of Science, complemented by reference tracking of key publications and international microbiological standards.
Priority was given to peer-reviewed studies describing the formulation, optimization, or comparative evaluation of selective and enrichment media used for Campylobacter isolation in food, clinical, and environmental contexts. Foundational reports and historical milestones that contributed to the conceptual or technical evolution of Campylobacter cultivation were also included to ensure a coherent representation of the field’s progression.
This narrative and interpretative approach aims to synthesize the main scientific trends, methodological advances, and practical innovations rather than exhaustively list all available studies. Emphasis is placed on identifying the rationale behind medium composition, the transition from empirical to standardized formulations, and the impact of these developments on diagnostic and food safety practices.
In addition to primary research articles, selected review papers, ISO guidelines, and reference protocols were consulted to contextualize major shifts in the understanding of Campylobacter physiology and its implications for culture medium design.
4. Evolution of Campylobacter Culture Media
The development of culture media for Campylobacter spp. represents a relatively recent phase in the history of microbiology. Reliable methods for isolating these organisms only began to emerge in the early 1970s, largely due to their fastidious growth requirements and pronounced sensitivity to environmental conditions. This delayed methodological progress contrasts sharply with that of other major foodborne pathogens, such as Listeria monocytogenes, which was isolated and scientifically described several decades earlier [20], and Escherichia coli, first described by Escherich in the late nineteenth century [21]. Consequently, Campylobacter remained underrecognized as a human and animal pathogen for much of the twentieth century.
Collectively, these studies illustrate how advances in Campylobacter culture media have been shaped by the evolving requirements of clinical diagnostics, veterinary surveillance, and food safety monitoring. The following sections present a chronological overview of these developments, highlighting the key scientific and technical milestones that led from early empirical approaches to the standardized and specialized media used today.
4.1. Pioneering Attempts (1970s)
Although Campylobacter spp. were not recognized as human pathogens until the 1970s, historical records indicate that these organisms had likely caused human and animal illness for decades. In 1886, Escherich described spiral-shaped bacteria in children who had died from “cholera infantum,” but he was unable to culture them successfully [22]. Similar organisms were later observed in sheep by McFadyean and Stockman (1909) [23], and isolated from cattle by Smith (1919), who named the organism Vibrio fetus [24,25]. Throughout the following decades, its pathogenic relevance in livestock became increasingly evident [26,27].
By the 1930s and 1940s, additional links between Vibrio-like organisms and enteric diseases were documented in calves, swine and, occasionally, humans [28,29,30]. King’s work in 1957 marked another important milestone, as he identified a “related vibrio” with unique physiological traits that hindered routine cultivation using existing laboratory methods [31]. Although King attempted to adapt fecal culture techniques, the absence of selective media and the organism’s fragile physiology resulted in poor recovery, contributing to its under-recognition in clinical microbiology.
A critical conceptual shift occurred when Sebald and Véron (1963) used DNA base composition to reclassify these vibrios under the new genus Campylobacter [32]. However, despite clearer taxonomic identity, isolation remained unreliable. Until the late 1960s, most attempts to culture Campylobacter spp. yielded inconsistent or non-viable growth, reflecting the need for selective media and improved atmospheric control.
In summary, early discoveries of Campylobacter-like organisms highlighted their clinical and veterinary relevance but also exposed the major technical barriers to their cultivation. The lack of appropriate media, inadequate incubation conditions, i.e., atmosphere and temperature systems, and reliance on empirical methods led to decades of underdiagnosis [10]. These foundational challenges ultimately set the stage for the first successful isolation from fecal samples in 1968–1972, marking the transition to the modern era of Campylobacter culture methods.
4.2. The Crucial Step
The turning point came in the late 1960s, when Dekeyser, in collaboration with Butzler and colleagues, developed the first reliable method for isolating Campylobacter directly from fecal material. This pivotal advance, achieved in 1968 and published in 1972, overcame many of the technical limitations that had hindered detection for nearly a century. They successfully isolated a ‘related vibrio’ (Campylobacter) from a patient’s blood and subsequently from feces using an innovative filtration technique. This method involved the differential filtration of fecal suspensions through filters with a pore size of 0.65 µm, enabling the passage of Campylobacter organisms. The filtrate was then applied to a selective growth medium (thioglycolate-agar medium supplemented with defibrinated sheep blood, bacitracin, polymyxin B sulfate, novobiocin and actidione) [33]. This landmark fecal culture confirmed that the source of the bacteremia was an intestinal infection.
In the following years, various authors [34,35,36,37,38,39] used the same methodology to isolate Campylobacter from bovine and sheep fetuses, human samples, and bull semen or preputial mucus, facing significant challenges and determining that numerous cultures failed to thrive in deep culture conditions.
In summary, the period from 1968 to the early 1970s marked the first reliable isolation of Campylobacter from fecal and clinical samples. Filtration-based methods [34,35] enabled critical early breakthroughs and broadened the recognized ecological and clinical range of the organism [36,37,38,39]. Yet, their technical complexity underscored the urgent need for more practical, selective culture media, setting the stage for the development of blood-based formulations in the late 1970s. As the limitations of filtration became increasingly clear, attention shifted toward developing selective blood-based media that offered greater reproducibility, simplified workflows, and improved recovery of thermophilic Campylobacter.
The early selective media summarized in Table 1 illustrate the empirical strategies initially employed to overcome the extreme fastidiousness of Campylobacter spp. Fluid thioglycolate-based formulations relied on strongly reduced environments, blood supplementation, and broad-spectrum antibiotics to permit survival during filtration-based isolation. The Butzler medium [40] represented an optimization rather than a conceptual departure from this approach, refining blood concentration and incubation conditions. In parallel, the VA-10 [35] formulations marked an early attempt to introduce more defined metabolic components, including succinate and iron salts, and to partially replace blood with hematin. Despite these innovations, all early media remained technically demanding, slow, and insufficiently selective, underscoring the need for simpler and more reproducible blood-based selective media developed in the late 1970s.
4.3. Blood-Based Selective Media (1977–1982)
The use of blood as a supplement in culture media was already well established in clinical microbiology for the recovery of fastidious and clinically relevant microorganisms, including Haemophilus spp. and Neisseria spp., as it provides essential growth factors, neutralizes inhibitory substances, and enhances bacterial growth and recovery [41].
In 1977, Skirrow [10] introduced a practical and highly influential method for isolating Campylobacter spp. from fecal samples, marking one of the most decisive advancements in Campylobacter diagnostics. He replaced the labor-intensive filtration step with a selective blood-based medium (Table 2). Incubation in a vacuum jar at approximately 43 °C suppressed competing enteric flora while supporting the growth of thermophilic Campylobacter species. Although Skirrow medium significantly improved selectivity and routine recoverability, the antibiotic combination introduced an inherent selection bias, inhibiting antibiotic-susceptible Campylobacter strains and reducing the recovery of non-C. jejuni/C. coli species. Building on this work, several research groups introduced blood-containing selective media that enhanced routine detection and eliminated the need for pre-enrichment or filtration [42,43,44,45,46,47]. These refinements focused on optimizing antibiotic combinations, stabilizing blood components, and improving growth performance across a wider range of sample types; however, their antibiotic composition and incubation conditions increasingly favored C. jejuni and C. coli, introducing an early species-level recovery bias. As these selective formulations became widely adopted, Campylobacter spp. rapidly gained recognition as common human pathogens, transitioning from previously underdiagnosed organisms to clinically significant agents of bacterial gastroenteritis.
The development of blood-based selective media during this period established the foundation for modern culture-based diagnostics (Table 2). These media enabled consistent recovery from fecal specimens, supported epidemiological investigations, and paved the way for further diversification, including the development of charcoal-based, antifungal-supplemented, and commercially formulated media suitable for food testing and automated laboratory workflows. Nevertheless, subsequent blood-based selective media, while enhancing the recovery of Campylobacter species, retained antibiotic-driven selection bias toward veterinary-associated Campylobacter species. Between 1977 and 1982, the introduction of selective blood-based media transformed Campylobacter detection from an experimental challenge into a routine laboratory procedure. Skirrow’s formulation offered the first practical alternative to filtration, and subsequent blood-containing media refined selectivity and reliability [42,43,44,45,46,47]. Lysed horse blood is incorporated into culture media for Campylobacter spp. because it provides essential growth factors and protection against inhibitory compounds. Campylobacter species are fastidious, microaerophilic organisms with limited capacity for de novo heme synthesis; therefore, erythrocyte lysis is required to release heme and hemin, which are critical for respiratory enzyme function and energy metabolism [48,49]. In addition, lysed blood supplies readily available iron and other cofactors and reduces oxidative stress by neutralizing reactive oxygen species and binding toxic metabolites such as free fatty acids and peroxides. Horse blood is preferred due to its consistent composition, lower variability, and reduced levels of inhibitory substances compared with other animal blood sources, and its use has been extensively validated in standardized media formulations for reliable recovery of Campylobacter, particularly from stressed samples [11].
These advances not only confirmed Campylobacter as a major enteric pathogen but also laid the groundwork for the transition, during the 1980s, toward charcoal-based and fully blood-free media. Although blood-based media (Figure 1) were initially successful, they presented inherent limitations, including variability associated with animal-derived components and difficulties in colony visualization. These challenges ultimately drove the development of charcoal-based and completely blood-free formulations, marking the next major phase in the evolution of Campylobacter culture methods.
4.4. Charcoal-Based and Non-Blood Media (1983–1990)
The period between 1983 and 1990 marked a decisive turning point in the development of growth media for Campylobacter spp., characterized by the gradual replacement of blood with activated charcoal and reducing agents (Table 3). This shift aimed to eliminate biological variability, improve colony visibility, and reduce costs associated with animal-derived ingredients. Activated charcoal functions as a non-specific adsorbent of toxic metabolites, peroxides, and fatty acids. Bolton et al. (1983) demonstrated this effect by systematically comparing basal media with and without charcoal, showing that charcoal-containing formulations enhanced aerotolerance and supported Campylobacter growth under otherwise inhibitory oxygen conditions, indicating removal of oxygen-derived toxic compounds from the medium, thereby reducing oxidative stress and creating a protective microenvironment for oxygen-sensitive Campylobacter cells [56].
The first significant advancement came from Bolton et al. (1983) [56], who introduced a blood-free medium supplemented with bacteriological charcoal, ferrous sulfate, and sodium pyruvate, creating a reductive environment that protected oxygen-sensitive organisms. A year later, Bolton et al. (1984) [43] refined the formulation to produce the selective medium Charcoal Cefoperazone Deoxycholate Agar (CCDA) (Table 3), which effectively inhibits competing flora while promoting the growth of C. jejuni and C. coli.
Subsequently, Hutchinson and Bolton (1984) [57] modified this formulation into the mCCDA (Figure 2), standardizing the cefoperazone concentration. This version later became the international reference medium incorporated into ISO 10272-1:2017 Part 1 [58] and ISO 10272-2:2017 Part 2 [59] protocols for food and animal feeding stuffs testing. Parallel to these developments, other researchers explored enrichment and semi-solid formulations. Doyle and Roman (1982) [50] proposed an enrichment broth based on Brucella broth, cysteine, and lysed blood, allowing rapid growth under agitation at 42 °C. In turn, Goossens et al. (1989) [46] introduced a semi-solid, blood-free motility medium (SSM) that exploited Campylobacter’s characteristic motility, facilitating differentiation from competing fecal flora.
The introduction of charcoal-based and blood-free formulations represented a paradigm shift, eliminating reliance on blood-containing media, improving productivity and oxygen tolerance, and laying the groundwork for the global standardization of Campylobacter isolation. However, the widespread use of cefoperazone in these media preferentially favors intrinsically resistant C. jejuni and C. coli, thereby reinforcing species-level selection bias.
4.5. Rise in Antifungal and Commercial Formulations (1991–2000)
During the 1990s, innovation focused on the integration of antifungal agents and the commercial availability of preformulated selective media, addressing the growing need for standardized, safer, and faster laboratory workflows (Table 4). In 1992, Stern et al. [60] developed the Campy-Cefex Agar (CCA), combining antimicrobials to suppress fungal and Gram-positive contaminants in poultry samples. Subsequently, Aspinall et al. (1993) [61] introduced the Cefoperazone Amphotericin B Teicoplanin (CAT) medium, incorporating amphotericin B and teicoplanin, further enhancing selectivity for thermophilic Campylobacter species, including C. upsaliensis. These refinements significantly improved the recovery of target isolates in mixed cultures.
The decade culminated with Jeffrey et al. (2000) [62], who designed a semi-solid aerobic medium containing rifampicin, cephalothin, and amphotericin B, allowing for Campylobacter recovery under non-microaerophilic conditions. This innovation anticipated the emergence of field-adapted and low-oxygen-tolerant media, bridging traditional laboratory culture and environmental surveillance methodologies.
4.6. Modern Complex and Enrichment Media (2001–2024)
The twenty-first century has witnessed unprecedented diversification in Campylobacter growth media, driven by technological advances and the demand for rapid, selective, and standardized detection in food safety and clinical diagnostics (Table 5). Line (2001) [64] introduced the Campy-Line Agar (CLA) and its blood-supplemented counterpart (CLBA), which integrated α-ketoglutaric acid, hemin, and 2,3,5-triphenyltetrazolium chloride (TTC) to allow visual differentiation of colonies and enhance detection sensitivity. Shortly thereafter, Oyarzabal et al. (2005) [65] and Thomas et al. (2005) [66] improved direct enumeration from poultry carcasses using formulations such as modified Campy-Cefex Agar (mCCA) and AAV medium, which incorporated amphotericin B, aztreonam, and vancomycin to reduce background contamination.
In 2006, the international standardization of Campylobacter culture methods was formalized through ISO 10272-1:2017 [58] and 10272-2:2017 [59], which established Bolton broth (for enrichment) and mCCDA (for selective isolation) as reference media. Around the same time, bioMérieux commercialized CampyFood Agar, facilitating consistent results and widespread adoption in routine testing.
Subsequent developments targeted antimicrobial resistance and recovery efficiency. Between 2012 and 2018, Chon et al. [67,68,69,70] produced a series of optimized formulations—including P-mCCDA, C-mCCDA, Modified Karmali, and Tz-Bolton broth—by supplementing traditional recipes with high concentrations of polymyxin B, potassium clavulanate, and tazobactam, improving detection of β-lactam-resistant isolates. Meanwhile, Yoo et al. (2014) [71] developed BRS agar, and Teramura et al. (2015) [72] introduced the first chromogenic medium (CM-HT) for Campylobacter spp., enabling rapid color-based identification.
Building on this concept, CHROMagar later commercialized CHROMagar™ Campylobacter (Figure 3), a selective chromogenic medium designed for the routine detection and enumeration of thermotolerant Campylobacter spp. in food, clinical and environmental samples. Although the exact year of its initial formulation is not publicly documented, its widespread adoption in the late 2010s reflects the consolidation of chromogenic technology as a standardized, user-friendly alternative to traditional charcoal- or blood-based media.
In the 2020s, innovation has emphasized metabolic optimization and sustainability. Ha et al. (2021) [73] formulated CSA-S50, a selective agar enriched with L-serine, improving energy metabolism and recovery rates for C. jejuni.
Most recently, Levican and Hinton (2022) [74] presented CAMPYAIR, the first aerobically cultivable selective medium, containing soluble starch, sodium deoxycholate, and TTC, enabling Campylobacter spp. growth in ambient oxygen conditions.
These modern formulations embody the fusion of classical microbiological principles with molecular-era innovation, advancing toward intelligent, adaptable, and eco-efficient culture systems capable of supporting both traditional and high-throughput applications [75].
Across the evolution of Campylobacter culture media, antibiotic supplementation has played a central role in shaping selectivity and recovery efficiency. Beyond their selective role, the composition of antibiotic supplements in Campylobacter culture media has evolved in direct response to the progressive emergence of antibiotic resistance among competing background microbiota in clinical, food and environmental matrices. As resistance to commonly used agents such as polymyxins, trimethoprim, quinolones and macrolides increased among Enterobacteriaceae, Pseudomonas spp., Aeromonas spp. and Gram-positive bacteria, classical selective formulations required continuous revision to preserve suppression of competing flora without compromising Campylobacter recovery. It should also be acknowledged that the use of antibiotic supplements represents a potential methodological bias, as Campylobacter strains susceptible to the antibiotics included in selective media may be partially or completely inhibited, leading to an underestimation of their occurrence in tested samples.
This dynamic interplay between selective pressure and background flora adaptation is clearly reflected in the historical transition from early blood-based media containing relatively simple antibiotic combinations (e.g., vancomycin–polymyxin–trimethoprim in Skirrow agar) to progressively more complex formulations incorporating cefoperazone, rifampicin, aztreonam, teicoplanin and antifungal agents. More recently, the inclusion of β-lactamase inhibitors in modified mCCDA and Bolton-derived formulations further exemplifies how increasing multidrug resistance among competing microorganisms has driven iterative media redesign.
Consequently, the evolution of Campylobacter selective media has been shaped not only by the need to enhance selectivity, but also by the necessity to adapt to changing resistance landscapes in accompanying microbiota, ensuring robust recovery of C. jejuni and C. coli from increasingly complex and environmentally challenging matrices. This trajectory aligns with a broader One Health perspective, highlighting the influence of antibiotic use and resistance dissemination along the food chain and in the environment on diagnostic culture methodologies. Table 6 summarizes the main antibiotics employed, their biological targets, typical concentration ranges, representative media, and historical period of use.
5. Overview, Challenges, and Future Directions
5.1. Technical and Diagnostic Impact
Over five decades, the evolution of Campylobacter growth media has transformed the landscape of clinical and food microbiology (Figure 4). The Historical and contemporary culture media used for the isolation, enrichment, and enumeration of Campylobacter spp., including basal compositions, selective supplements, incubation time, temperature and atmospheric conditions era summarized in Appendix A (Table A1).
The introduction of blood-free, charcoal-based formulations in the 1980s represented a pivotal step toward reproducibility and accessibility. These media, exemplified by Bolton’s CCDA and Hutchinson’s mCCDA, addressed major limitations of earlier blood-containing systems, such as lot-to-lot variability and interference from heme compounds
The adoption of mCCDA in the ISO 10272-1:2017 [58] and ISO 10272-2:2017 [59] standards further cemented its role as a global benchmark for Campylobacter isolation in both clinical and food matrices. In addition, the Bacteriological Analytical Manual (BAM) [63] of the U.S. Food and Drug Administration specifically states that Campylobacter isolation should be performed using one of two selective agars: Abeyta–Hunt–Bark (AHB) [64] agar or modified Campy blood-free agar (mCCDA) [58,59], thereby formally recognizing both media as suitable options for the isolation of Campylobacter spp. in food microbiological analysis.
The refinement of enrichment broths, including Bolton broth and its derivatives (R-, C-, and Tz-Bolton), has also been crucial. These formulations enhanced recovery from low-level or stressed cells, enabling reliable detection even in complex samples such as poultry rinses, dairy products, and environmental water.
Collectively, the integration of selective agents such as cefoperazone, vancomycin, and trimethoprim standardized selectivity across laboratories and reduced false negatives caused by competing flora.
Furthermore, the development of antifungal and β-lactamase–inhibitor-supplemented media during the 1990s and 2000s (e.g., CAT, AAV, P-mCCDA, and C-mCCDA) addressed emerging issues of antimicrobial resistance and contamination, aligning culture performance with evolving microbial ecology.
Commercial production by companies such as bioMérieux facilitated widespread access to standardized formulations, enabling harmonized monitoring by EFSA, FDA, and WHO surveillance programs.
Decades of methodological development have resulted in a wide range of selective media for the isolation of Campylobacter spp. However, comparative studies consistently demonstrate that no single medium ensures optimal recovery across all sample matrices and species. In the present evaluation, all media complied with ISO 11133:2014 [80], productivity requirements (PR ≥ 0.5) and showed comparable specificity based on typical Campylobacter colony morphology followed by mandatory confirmation. Consequently, the main differences between media relate to their degree of selectivity and the trade-off between suppression of competing flora and recovery of stressed or less dominant species, as summarized in Table 7. Qualitative performance categories (Low, Moderate, High, Very high) were assigned based on consistent comparative trends reported across multiple studies, including relative recovery rates, detection frequencies, background flora suppression, and reference status in standardized protocols, rather than on absolute quantitative thresholds.
Charcoal-based, blood-free media such as mCCDA are widely adopted in food safety and surveillance due to their high selectivity and operational simplicity. Nevertheless, several studies have shown that cefoperazone-containing media may underrepresent non-jejuni/coli species, contributing to a recognized recovery bias. In contrast, blood-based media, including Preston agar, provide effective suppression of background flora and may support broader species recovery, albeit with higher costs and greater logistical complexity.
Beyond the choice of solid medium, methodological parameters such as direct plating versus selective enrichment, incubation temperature and atmospheric conditions further influence recovery outcomes. Direct plating preserves quantitative information and species diversity but is less sensitive at low contamination levels, whereas selective enrichment increases detection sensitivity at the expense of representativeness. Together, these considerations highlight that culture-based diagnostics for Campylobacter continue to evolve, with ongoing efforts to balance selectivity, sensitivity and practical applicability.
5.2. Limitations and Persistent Challenges
Despite major progress, culture-based detection of Campylobacter spp. remains challenging due to their fragile physiology and microaerophilic dependency. Traditional media still require controlled atmospheres (typically 5% O_2_, 10% CO_2_, 85% N_2_) and precise temperature regulation (41–43 °C), limiting their use in decentralized or resource-poor settings.
A recurrent limitation highlighted across comparative studies is the historical optimization of selective media for C. jejuni and C. coli. Antibiotic supplements, incubation at 42 °C and selective atmospheres systematically disadvantage non-thermotolerant and emerging Campylobacter species, including C. lari, C. upsaliensis, C. fetus and C. hepaticus. Recent species-focused studies demonstrate that commonly used media may fail entirely for these organisms unless formulations, supplements and incubation parameters are specifically adapted. These findings indicate that culture-based surveillance data should be interpreted with caution, as apparent epidemiological dominance may partially reflect methodological bias rather than true prevalence [93].
Additionally, sub-lethally injured or viable but non-culturable (VBNC) cells may evade recovery even on optimized media, leading to underestimation of prevalence in environmental or processed food samples [94]. VBNC Campylobacter cells are of particular concern in food safety and public health, as they may retain pathogenic potential while escaping detection by conventional culture-based methods. This can result in underestimation of contamination levels in foods, especially in matrices subjected to stress conditions such as refrigeration, freezing, drying, or exposure to sanitizers, and complicates quantitative risk assessment and epidemiological surveillance [95]. Furthermore, VBNC cells may resuscitate under favorable conditions along the food chain or within the host, highlighting their potential role in foodborne transmission [96]. The persistence of VBNC cells therefore reinforces the need for integrated detection strategies that combine optimized culture media with molecular approaches, rather than viewing these methodologies as mutually exclusive [94].
Moreover, the continued reliance on antibiotic supplementation, while essential for selectivity, raises ecological and biosafety concerns, particularly regarding the disposal of antimicrobial-containing waste [97].
5.3. Emerging Trends and Future Directions
Contemporary research is steering Campylobacter culture toward closer integration with molecular and omics-based methods. Several promising trends are evident: aerobic and oxygen-tolerant media enable simplified incubation without the need for microaerophilic systems, facilitating field applications and automation [92]. In addition, chromogenic and differential systems enhance colony identification and allow semi-quantitative enumeration, reducing reliance on confirmatory biochemical testing [72].
Together, these developments are transforming Campylobacter culture from a purely diagnostic tool into a broader research platform that increasingly complements molecular detection approaches. Despite major advances in polymerase chain reaction (PCR) and metagenomics, culture-based methods remain indispensable and are expected to retain a central role in future surveillance and research frameworks [98]. The recovery of viable isolates is essential for strain-level sequencing, outbreak investigation, source attribution, and the study of Campylobacter population structure and evolutionary dynamics over time [99]. In addition, culture-based isolation enables antimicrobial susceptibility testing and phenotypic characterization, while providing the reference material required to validate and contextualize molecular findings [99]. By contrast, non-culture-based approaches cannot reliably distinguish viable from non-viable or injured cells, limiting their applicability for quantitative risk assessment, regulatory decision-making, and long-term epidemiological surveillance.
In addition to these general trends, recent studies have proposed concrete modifications to culture media composition and design. These include aerobic selective formulations incorporating antioxidant components and adjusted antibiotic combinations to support Campylobacter growth under atmospheric conditions, as well as revised selective formulations that reduce inhibition of non-jejuni/coli species by modifying or limiting specific antibiotics [74]. Furthermore, studies have demonstrated that recovery of underrepresented Campylobacter species, such as C. fetus and C. hepaticus, often requires tailored supplementation and incubation parameters, as standard selective media are frequently inhibitory to these organisms [100,101].
Beyond incremental modifications of existing formulations, future development of Campylobacter culture media may benefit from conceptual shifts in design. One emerging direction is the use of modular or purpose-driven media systems, in which a common basal formulation is combined with adjustable selective and supplement modules tailored to the sample matrix and diagnostic objective, rather than relying on a single universal medium. Such an approach could help mitigate the trade-offs between selectivity and species diversity that characterize current formulations.
In parallel, there is growing interest in culture strategies that incorporate a preliminary recovery or resuscitation phase for sub-lethally injured or VBNC (viable but non-culturable) Campylobacter cells prior to exposure to selective pressure [94]. Media designs that transiently reduce oxidative and antibiotic stress may improve recovery of stressed but viable cells, with important implications for food safety surveillance and public health risk assessment.
6. Conclusions
The historical trajectory of Campylobacter spp. culture media—spanning from thioglycolate agar from Dekeyser’s (1972) [33] to CAMPYAIR from Levican and Hinton (2022) [74]—reflects an ongoing pursuit of precision, efficiency, and adaptability. Each generation of media has addressed the scientific challenges of its era: from isolation feasibility, through selectivity and standardization to modern compatibility with high-throughput technologies.
Today, the focus shifts toward multifunctional media that are not only selective and sensitive but also ecologically responsible and digitally integrable. Future breakthroughs are likely to emerge from computational design, metabolomic mapping, and synthetic biology, yielding next-generation media capable of dynamically adapting to pathogen physiology and environmental conditions.
In essence, the evolution of Campylobacter growth media encapsulates the broader story of microbiology itself—transforming from art to precision science, from isolation to innovation. By providing a chronological and mechanistic synthesis of Campylobacter culture media development, this review fulfills its objective of contextualizing past innovations while identifying future directions for diagnostic and food safety applications.
By integrating strain-specific genomic sequencing with artificial intelligence–driven modeling, next-generation culture media might be developed to enhance selective growth and recovery.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Sheppard S.K. Maiden M.C.J. The Evolution of Campylobacter jejuni and Campylobacter coli Cold Spring Harb. Perspect. Biol.20157 a 01811910.1101/cshperspect.a 01811926101080 PMC 4526750 · doi ↗ · pubmed ↗
- 2Kaakoush N.O. Castaño-Rodríguez N. Mitchell H.M. Man S.M. Global Epidemiology of Campylobacter Infection Clin. Microbiol. Rev.20152868772010.1128/CMR.00006-1526062576 PMC 4462680 · doi ↗ · pubmed ↗
- 3Kirk M.D. Pires S.M. Black R.E. Caipo M. Crump J.A. Devleesschauwer B. Döpfer D. Fazil A. Fischer-Walker C.L. Hald T. World Health Organization Estimates of the Global and Regional Disease Burden of 22 Foodborne Bacterial, Protozoal, and Viral Diseases, 2010: A Data Synthesis P Lo S Med.201512 e 1001921 Erratum in P Lo S Med. 2015, 12, e 100194010.1371/journal.pmed.100192126633831 PMC 4668831 · doi ↗ · pubmed ↗
- 4Liu F. Lee S.A. Xue J. Riordan S.M. Zhang L. Global Epidemiology of Campylobacteriosis and the Impact of COVID-19Front. Cell. Infect. Microbiol.20221297905510.3389/fcimb.2022.97905536519137 PMC 9742372 · doi ↗ · pubmed ↗
- 5The European Union One Health 2024 Zoonoses Report EFSA J.202523 e 975910.2903/j.efsa.2025.975941377167 PMC 12686834 · doi ↗ · pubmed ↗
- 6Igwaran A. Okoh A.I. Human Campylobacteriosis: A Public Health Concern of Global Importance Heliyon 20195 e 0281410.1016/j.heliyon.2019.e 0281431763476 PMC 6861584 · doi ↗ · pubmed ↗
- 7Ricke S.C. Feye K.M. Chaney W.E. Shi Z. Pavlidis H. Yang Y. Developments in Rapid Detection Methods for the Detection of Foodborne Campylobacter in the United States Front. Microbiol.20189328010.3389/fmicb.2018.0328030728816 PMC 6351486 · doi ↗ · pubmed ↗
- 8Denis M. Rose V. Nagard B. Thépault A. Lucas P. Meunier M. Benoit F. Wilhem A. Gassilloud B. Cauvin E. Comparative Analysis of Campylobacter jejuni and C. coli Isolated from Livestock Animals to C. jejuni and C. coli Isolated from Surface Water Using DNA Sequencing and MALDI-TOF Pathogens 202312106910.3390/pathogens 1209106937764877 PMC 10535298 · doi ↗ · pubmed ↗