Genetic Polymorphism of IL-1A, IL-1B and TNFA Predicting the Presence of Periodontopathogenic Bacteria
Nina Kalajzic, Ajka Pribisalic, Marina Adriana Jezina Buselic, Samra Prentic Bakic, Dunja Petricic, Ferdinand Josip Buselic, Davorka Sutlovic, Sendi Kuret

TL;DR
This study finds that genetic variations in IL-1A and IL-1B are linked to the presence of certain bacteria in periodontitis, suggesting a genetic influence on disease progression.
Contribution
The study identifies specific cytokine gene polymorphisms associated with periodontopathogenic bacteria in a Croatian population.
Findings
The IL-1A CC genotype increases odds of P. gingivalis and T. denticola presence.
The IL-1B CC genotype is independently associated with T. forsythia presence.
TNFA polymorphism showed no significant associations with periodontal bacteria.
Abstract
Background/Objectives: Periodontitis is a chronic inflammatory disease characterized by complex interactions between periodontal pathogens and the host immune response. Pro-inflammatory cytokines, particularly interleukins, may influence bacterial colonization and disease expression, but their association with specific periodontal pathogens remains unclear. This study investigated the associations between single-nucleotide polymorphisms in IL-1A, IL-1B, and TNFA and the presence of key periodontopathogenic bacteria in patients from Croatia. Methods: A cross-sectional study included 63 participants. Genotypes were determined, and subgingival plaque samples were analyzed for Aggregatibacter actinomycetemcomitans, Porphyromonas gingivalis, Prevotella intermedia, Tannerella forsythia, and Treponema denticola using real-time PCR. Multivariable logistic regression models assessed associations…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
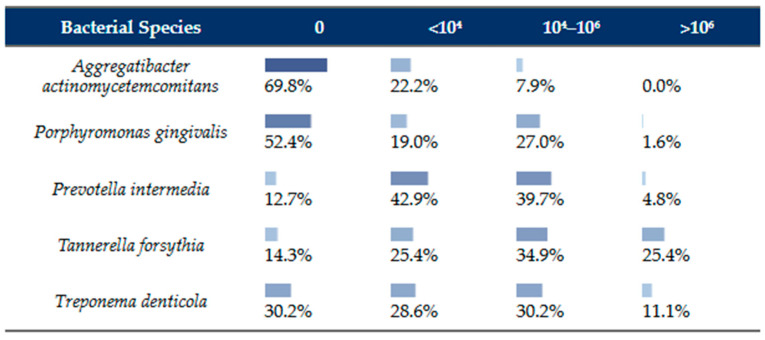 Figure 1
Figure 1- —University of Split, Department of Health Studies
- —Faculty of Health Sciences, University of Split
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsOral microbiology and periodontitis research · Salivary Gland Disorders and Functions · Sinusitis and nasal conditions
1. Introduction
Periodontal disease is a chronic inflammatory condition affecting the tissues supporting the teeth and is characterized by progressive destruction of the periodontal ligament and alveolar bone. If left untreated, this process may ultimately result in tooth loss and impaired oral function [1,2]. Periodontal disease is multifactorial in nature and develops as a consequence of dysregulated host immune response to periodontopathogenic bacteria colonizing the oral cavity [3].
Although periodontal disease is considered an infection-related condition, its onset and progression depend on complex interactions between microbial, environmental and host-related factors, with specific periodontopathogenic bacteria identified as crucial etiological agents [4]. The presence of specific periodontopathogenic bacteria is essential for disease initiation. The most prevalent species detected in subgingival plaque include Aggregatibacter actinomycetemcomitans (A. actinomycetemcomitans), Porphyromonas gingivalis (P. gingivalis), Treponema denticola (T. denticola), Tannerella forsythia (T. forsythia), Treponema denticola (T. denticola), and Prevotella intermedia (P. intermedia). These predominately Gram-negative anaerobic bacteria (with A. actinomycetemcomitans being facultatively anaerobic) express numerous virulence factors, such as lipopolysaccharides, proteolytic enzymes, fimbriae, leukotoxins, and immunomodulating proteins, which facilitate bacterial adhesion, invasion, and immunomodulation [3,5,6,7].
The bacterium A. actinomycetemcomitans is often considered separately, and its presence is associated with three- to four-fold faster bone resorption compared to other periodontopathogens [1,7]. Its presence is often associated with aggressive forms of periodontal disease, but it can also be detected in chronic conditions [8]. This facultative anaerobe possesses a variety of virulence mechanisms, of which the most notable is the production of leukotoxin, which particularly targets the host’s immune cells, contributing to immune system dysregulation and periodontal tissue damage [3,8].
The red complex bacteria—P. gingivalis, T. denticola, and T. forsythia—are strongly associated with advanced periodontal disease and severe periodontal tissue destruction. These bacteria are associated with the formation of subgingival plaque and deep periodontal pockets, as well as bleeding on probing and clinical attachment loss. Furthermore, research indicates that these periodontopathogens exhibit a synergistic effect and cause the host immune response, both innate and adaptive immunity, resulting in the secretion of cytokines, such as interleukin-1 alpha (IL-1α), interleukin-1 beta (IL-1β) and tumor necrosis factor-alpha (TNF-α) [1,2,3,9].
Within this group, P. gingivalis is regarded as a key pathogen due to its capacity to alter the host immune response and promote dysbiosis in the oral cavity, causing tissue damage and chronic inflammation [10,11]. T. denticola, a highly mobile spirochete, contributes to periodontal destruction through proteolytic activity, disruption of epithelial cell membranes and direct interaction with immune cells, whereas T. forsythia produces proteases that evade the host’s immune response [9,12].
The Prevotella genus has high diversity, and some of the 30 subspecies, including P. intermedia, are members of the orange complex. P. intermedia produces a capsule that impairs phagocytosis and enhances intercellular survival, thereby contributing to persistent infection [9,13].
Although the presence of periodontopathogens is one of the necessary factors for disease development, it is evident that the presence of microorganisms alone cannot entirely explain the heterogeneity in disease susceptibility, severity, and progression observed among individuals. Therefore, periodontal disease is currently considered to be caused by interactions between periodontopathogenic bacteria, environmental factors, the host immune system, and particularly genetic variants that can modulate inflammatory and immune system responses [2,14].
Genetic variants, i.e., single-nucleotide polymorphisms (SNPs) in several inflammatory cytokine genes, including IL-1A, IL-1B, and TNFA, have been studied and are associated with the progression of periodontal disease [15]. Moreover, genome-wide association studies suggest that host genetic variability may also influence the composition of the subgingival microbiota [16,17].
One of the most studied pro-inflammatory cytokines associated with periodontal disease is IL-1. The human IL-1 genes are located on the proximal part of the long arm of chromosome 2 and encode IL-1α and IL-1β inflammatory cytokines [15]. A meta-analysis by Feng et al., including 1356 patients with chronic periodontitis and 249 controls, demonstrated an association between the IL-1A (−889 C/T) polymorphism and susceptibility to chronic periodontitis in European populations [18]. Additionally, a Spanish study reported a significant association between IL-1A polymorphism and increased P. intermedia levels [19]. Pani et al. observed a statistically significant association between IL-1β (+3954 C/T) polymorphism and red complex bacteria, accompanied by elevated IL-1β levels in gingival crevicular fluid [20]. However, other studies failed to confirm these associations in European populations, highlighting inconsistent findings [21,22].
TNF-α is another key pro-inflammatory cytokine, primarily mediated by monocytes and macrophages during the early host immune response to bacterial colonization [23]. The TNFA gene has an important role in amplifying inflammatory cascades, inducing other cytokines such as IL-1, and regulating cell proliferation, necrosis, and apoptosis [24]. The TNFA (−308 G/A) polymorphism has been associated with the pathogenesis of aggressive periodontitis, contributing to the over-release of other enzymes, such as proteases, leading to alveolar bone resorption [24,25].
Overall, previous research reported inconsistent findings regarding associations between genetic variants in IL-1A, IL-1B and TNFA genes and either the presence of periodontopathogenic bacteria or susceptibility to periodontitis in different populations. Notably, to our knowledge, only a limited number of studies have directly examined the relationship between periodontopathogenic bacteria and cytokine gene polymorphism, and none have been conducted in the Croatian population. Therefore, our study aimed to investigate the association between the presence of five key periodontopathogenic bacteria and SNPs in IL-1A (−889 C/T), IL-1B (+3954 C/T; in the literature, it is also referred to +3953 C/T) [26] and TNFA (−308 G/A) genes, to evaluate their potential role as genetic risk markers for periodontal disease.
2. Materials and Methods
2.1. Patients and Sample Collection
A total of 63 patients diagnosed with chronic periodontitis (according to the American Academy of Periodontology and European Federation of Periodontology) [27] and positive for at least one periodontopathogenic bacterium were included in this study. None of the patients have received any antimicrobial medications at least three months before the enrollment in the study. Dental examination and microbiological sampling were performed by the same dentist in a private dental office in Split, Croatia. A standardized periodontal examination was performed for all participants to objectively assess oral inflammatory status and periodontal tissue condition. Clinical parameters included gingival index (Löe and Silness), plaque index (Silness and Löe), probing depth (PD), bleeding on probing (BOP), and clinical attachment level (CAL). The diagnosis of periodontitis was made for patients who exhibited PD of ≥4 mm and CAL of 3–4 mm.
Sample collection and analysis were obtained between June 2024 and October 2025. Data for each individual were collected from medical documentation. Pooling method was used for harvesting bacteria from the pocket using 6 paper points of moderate strength (average size, # 30–50) and “pooling” several probes from various pockets equally distributed in both jaws, following which the paper point (samples) would be placed in a cold transport container, delivered to the laboratory and kept at a temperature of +4 °C until the start of the analysis.
All participants provided written informed consent prior to inclusion in the study, agreeing to the use of their personal and clinical data for research purposes. Information regarding systemic conditions, including cardiovascular diseases, diabetes mellitus, and rheumatoid arthritis, as well as smoking status and snoring, was obtained through a structured questionnaire completed at the time of consent. To minimize potential confounding, patients were additionally questioned about current medication use, dietary habits and lifestyle factors.
2.2. Detection of Periodontopathogenic Bacteria
2.2.1. Microbiological Sampling
Cotton rolls were used to segregate the sampling area. The tooth surface was cleaned with 70% ethanol and dried with sterile cotton swabs. Samples were obtained from the diseased areas’ deepest pockets with five sterile paper points inserted into the gingival crevice for 15 s and then placed in a sterile 1.5 mL vial for molecular analysis. The samples were sent to a molecular laboratory and had 7-day stability at temperatures ranging from 2 to 8 °C.
2.2.2. DNA Extraction from Periodontal Sample
To extract the DNA from the periodontal sample, 500 µL of saline was added to the tube and mixed vigorously for 10 s to remove bacterial cells from the paper points. The DNA was then extracted using the NucleoSpin^®^-Microbial kit (Macherey-Nagel, Duren, Germany) according to the manufacturer’s instructions. The quantity of extracted DNA was quantified spectrophotometrically at 260 nm with a NanoDrop ND-1000 Spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA), and its purity was calculated as the ratio of absorbance observed at 260 and 280 nm.
2.2.3. Real-Time PCR for Bacteria Detection
Real-time PCR was carried out using a real-time analyzer, the ABI Prism 7500 Real-Time PCR System (Applied Biosystems, Waltham, MA, USA). The primers for A. actinomycetemcomitans, P. gingivalis, P. intermedia, T. denticola, and T. forsythia were designed as previously described [28].
The real-time PCR was performed in a total reaction volume of 50 µL, which included 5.0 µL of isolated DNA as the template, 25 µL of Power SYBR Green PCR Master Mix (Thermo Fisher Scientific, Waltham, MA, USA), 18 µL of sterile water, and 2 µL (20 µM) of a bacteria-specific primer pair. The primer concentrations were the same for all assays. Negative and positive controls were included in each batch of specimens. Negative controls contained ultrapure water instead of sample DNA. The positive control consisted of the genomic DNA of the five positive targeted bacteria. The 5.0 µL of negative and 5.0 µL of positive control were included in each analysis run. All amplifications and detections were carried out in a MicroAmp optical 96-well reaction plate. The cycling conditions were initial denaturation at 95 °C for 10 min, followed by 40 cycles of denaturation at 95 °C for 5 s, and annealing at 60 °C for 34 s each. The accumulation of PCR products was observed at each cycle by monitoring the increase in fluorescence of the reporter dye from dsDNA binding SYBR Green. After the PCR, the specificity of the amplification was assayed with the use of melting curves which were constructed in the range of 60 °C to 95 °C [29].
2.3. Analysis of Genetic Polymorphisms
2.3.1. Sampling
For the analysis of genetic polymorphisms, five milliliters of venous blood was collected from each participant and transferred into a vacutainer containing 3% ethylenediaminetetraacetic acid (EDTA) to prevent coagulation. Samples were subsequently stored at −20 °C until DNA extraction.
2.3.2. DNA Extraction from Blood Samples
Genomic DNA was extracted from blood samples using a commercially available DNA isolation kit (High Pure PCR Template Preparation Kit, Roche, Basel, Switzerland) according to the manufacturer’s instructions. The DNA concentration was assessed using a NanoDrop spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA).
2.3.3. Real-Time PCR for SNPs Detection
Primer and probe sequences for IL-1A, IL-1B and TNFA polymorphisms (IL-1A −889: rs1800587; IL-1B +3954: rs1143634; TNFA −308: rs1800629) were obtained from Applied Biosystems, Waltham, MA, USA (assay ID for rs1800587: C_9546481_30; assay ID for rs1143634: C_9546517_10; assay ID for rs1800629: C_7514879_10).
Polymorphism detection was done using the ABI Prism 7500 Real-Time PCR System (Applied Biosystems, Waltham, MA, USA). Real-time PCR was carried out using a 25 µL reaction mix solution containing 12.50 µL 2xTaqMan^®^ Genotyping Master Mix (Thermo Fisher Scientific, Waltham, MA, USA), 1.25 µL SNP Assay, 2 µL of DNA. PCR cycling conditions included an initial denaturation at 95 °C for 10 min, followed by 40 cycles of 95 °C for 15 s and 60 °C for 1 min.
2.4. Statistical Analysis
Statistical analyses were performed using standard descriptive and inferential methods. The distribution of numerical variables was assessed for normality using the Kolmogorov–Smirnov test. As the data were not normally distributed, numerical variables are presented as median and interquartile range (IQR), whereas categorical variables are expressed as absolute numbers and percentages.
The prevalence of bacterial species and the genotype distribution in IL-1A, IL-1B and TNFA genes were compared using the Chi-square test or Fisher’s exact test, as appropriate, depending on expected cell frequencies.
The logistic regression models were conducted to examine whether genetic variants in selected genes predicted the presence of specific periodontopathogenic bacteria. IL-1A, IL-1B, and TNFA were entered as primary independent variables, and the presence of each bacterial species (A. actinomycetemcomitans, P. gingivalis, P. intermedia, T. forsythia, T. denticola; coded 1 for present, 0 for absent) was used as the outcome in five separate models. All models were adjusted for age (continuous), sex (female vs. male), systemic disease (yes/no), and smoking status (non-smoker vs. smoker) as potential confounders. Additional clinical variables, including snoring, number of natural teeth, number of implants, and periodontitis location, were initially examined in univariate logistic regression models, but none reached statistical significance and were therefore not included in the multivariable models given the sample size limitations. No further variable selection methods were applied, as all covariates were chosen based on clinical relevance.
Associations were expressed as odds ratios (ORs) with 95% confidence intervals (95% CI) and p-values based on Wald tests; model fit was evaluated with Nagelkerke’s R^2^, model χ^2^ p-value, and the area under the receiver operating characteristic curve (AUC).
All statistical tests were two-sided, with p-values < 0.05 considered statistically significant. Analyses were performed using R software (version 4.5.2, R Foundation for Statistical Computing, Vienna, Austria).
Hardy–Weinberg equilibrium (HWE) with the chi-square test was assessed for each SNP using an online Hardy–Weinberg Equilibrium Calculator [30]. A p-value > 0.05 was interpreted as no significant deviation from expected genotype distribution.
This study was conducted and reported in accordance with the STROBE guidelines for cross-sectional studies.
3. Results
3.1. Demographic and Clinical Characteristics of Patients
The analysis included 63 participants, whose demographic and clinical characteristics are summarized in Table 1. Slightly more participants were female (58.7%) than male (41.3%), although this difference was not statistically significant (p = 0.077). The median age was 57.0 years (IQR 43.5–67.0). Most individuals were free of systemic diseases (n = 42, 66.7%), while 21 (33.3%) had at least one condition. Among those with systemic conditions, elevated blood pressure was most prevalent (n = 17, 27.0%), followed by hypercholesterolemia (n = 6, 9.5%), and diabetes mellitus or rheumatoid arthritis (n = 2, 3.2% each). A few participants presented with multiple conditions, most commonly elevated blood pressure combined with one of the other disorders. Smoking was reported by 22 participants (34.9%), and 24 (38.1%) reported snoring. Participants retained a median of 21 natural teeth (IQR 9.5–26.0; range 0–32) and had a median of 4 implants (IQR 0.0–6.0; range 0–12), reflecting a wide range of oral status within the cohort. Regarding location of periodontitis, patients had a generalized (n = 55, 87.3%) or localized form (n = 8, 12.5%).
3.2. Distribution of Periodontopathogenic Bacteria Among Patients
Real-time PCR analysis detected all five target periodontopathogens in subgingival plaque samples, with bacterial load stratified into four categories (0: undetected; <10^4^ copies/plaque sample; 10^4^–10^6^ copies; >10^6^ copies).
A. actinomycetemcomitans exhibited the lowest detection rate at 30.2% (19/63), predominantly at low loads (<10^4^ copies in 14 samples; 10^4^–10^6^ in 5), with no samples exceeding 10^6^ copies. P. gingivalis was detected in 47.6% (30/63 samples), mainly at moderate loads (10^4^–10^6^ in 17 samples, 10^4^ in 11 samples), and one sample at >10^6^ copies. P. intermedia showed high prevalence (87.3%, 55/63), distributed across low (27 samples), moderate (25 samples), and high loads (3 samples). T. forsythia had an 85.7% detection rate (54/63) and the highest bacterial burdens, with 16 samples >10^6^ copies, alongside 22 at 10^4^–10^6^ and 16 at <10^4^. T. denticola was detected in 69.8% of samples (44/63), with the majority at moderate loads (19 samples at 10^4^–10^6^ copies), followed by low loads (18 at <10^4^), and fewer at high loads (7 at >10^6^). These results indicate that P. intermedia, T. forsythia, and T. denticola were dominant periodontopathogens in this cohort, as seen in Figure 1.
3.3. Genotype and Allele Frequencies of Study Participants
Presence of IL-1A, IL-1B and TNFA polymorphisms and genotype distribution in study participants are presented in Table 2. The genotype frequencies of the three studied SNPs showed no significant deviation and confirmed to be in Hardy–Weinberg equilibrium (p > 0.05).
3.4. Distribution of IL-1A, IL-1B and TNFA Genotypes
The prevalence of bacterial species and genetic polymorphisms in IL-1A, IL-1B, and TNFA genes is summarized in Table 3. Overall, genetic polymorphisms varied between patients, with some differences statistically significant and others not. For A. actinomycetemcomitans and P. intermedia, genetic polymorphisms were similar between groups, with no significant differences. In IL-1A and IL-1B genes, CC genotype was significantly associated with increased loads of P. gingivalis (p = 0.032), whereas differences in genetic polymorphisms in TNFA were not significant. A higher prevalence of T. forsythia was significantly associated with CC genotype in IL-1B gene, while IL-1A and TNFA were not significantly different. For T. denticola, genetic polymorphisms in selected genes did not differ significantly between groups.
Five multivariable logistic regression models were constructed to evaluate whether genetic polymorphisms in IL-1A, IL-1B, and TNFA genes predict the presence of selected periodontal bacteria (A. actinomycetemcomitans, P. gingivalis, P. intermedia, T. forsythia and T. denticola) in patients from the Split–Dalmatia County, after adjustment for age, gender, smoking status, and systemic disease. Spearman correlation analysis demonstrated a strong positive association between IL-1A and IL-1B polymorphisms (ρ = 0.629, p < 0.001). To prevent multicollinearity and unstable coefficient estimates, genetic polymorphisms in IL-1A and IL-1B genes were not entered simultaneously into multivariable logistic regression models. Instead, separate models were fitted including either IL-1A or IL-1B genetic variant, together with the remaining covariates. Overall, the associations between genetic polymorphisms in selected genes and bacterial presence were species-specific, while demographic and clinical covariates showed limited predictive value Table 4.
No statistically significant associations were observed between A. actinomycetemcomitans or P. intermedia presence and any of the investigated genetic polymorphisms or covariates, indicating that the examined polymorphisms do not independently predict A. actinomycetemcomitans nor P. intermedia colonization in our sample. Increasing age was independently associated with a higher likelihood of P. gingivalis detection (OR = 1.06; p = 0.030). Importantly, carriers of the IL-1A CC genotype had a significantly increased probability of P. gingivalis presence compared with CT and TT genotype carriers (OR = 5.54; p = 0.009). No significant associations were found for TNFA polymorphism, smoking status, gender, or systemic disease. This model showed good overall performance, supporting a role of IL-1A genetic variability in susceptibility to P. gingivalis colonization. A strong and independent association was observed between IL-1B CC genotype and the presence of T. forsythia, with carriers exhibiting markedly higher odds of bacterial detection compared with CT and TT genotype carriers (OR = 8.48, p = 0.026). Other polymorphisms and covariates were not significant predictors. Among all models, this model showed the best fit and discriminatory capacity, indicating a potentially important role of IL-1B genetic variants in host susceptibility to T. forsythia (Nagelkerke R^2^ = 0.416; AUC = 0.886; model p = 0.011). Carriers of the IL-1A CC genotype had significantly higher odds of T.denticola presence compared with CT and TT genotype carriers (OR = 3.77; p = 0.041). No significant associations were identified for genetic polymorphisms in IL-1B and TNFA genes, or demographic and clinical variables. Although the overall model was not statistically significant, the observed association suggests a possible contribution of IL-1A polymorphism to colonization by this periodontal pathogen.
4. Discussion
In this population from Split–Dalmatia County, IL-1A and IL-1B genetic polymorphisms, but not TNFA polymorphism, were significantly associated with the presence of specific periodontopathogenic bacteria. Specifically, the IL-1A (−889 C/T) CC genotype significantly predicted the presence of P. gingivalis (p = 0.009) and T. denticola (p = 0.041), while the IL-1B (+3954 C/T) CC genotype significantly predicted the presence of T. forsythia (p = 0.026). These results suggest that host genetic variability in pro-inflammatory interleukin genes may contribute to bacterial colonization patterns in periodontal disease, highlighting the importance of host–microbe interactions in the pathogenesis of periodontitis.
Key pro-inflammatory cytokines, including IL-1α, IL-1β and TNF-α, play a central role in modulating the host inflammatory response to periodontopathogenic bacteria, contributing to tissue remodeling, and directly influencing alveolar bone resorption. Polymorphisms in the IL-1A, IL-1B, and TNFA genes have previously been suggested as genetic risk factors; however, results of previous studies have been inconsistent, likely due to differences in disease phenotype, ethnic structure, and methodology [20,26,31,32].
A recent meta-analysis reported that IL-1A −889 C/T T allele had a favorable association in the prevention of periodontitis risk with OR (95% CI), 1.12 (0.99–1.25). In contrast, IL −889 C/T C allele had a strong association with the periodontitis development with OR (95% CI), 0.75 (0.66–0.85) which is consistent with our results [33]. The results of the study conducted by Loo et al. demonstrate that the IL-1A CC genotype (OR = 4.368; 95% CI = 2.309 to 8.264) was significantly associated with periodontitis [34]. In our study, there was a significant association between IL-1A CC genotype carriers and increased probability of P. gingivalis and T. denticola presence. In contrast, meta-analysis conducted by Feng and Liu indicated that IL-1A −889 TT genotype was closely related to the susceptibility of chronic periodontitis in different populations, including the European population [18].
In their research Mesa et al. analyzed the association of polymorphisms in the IL-1 gene cluster and the load of periodontopathogenic bacteria, where IL-1A C/T, T allele carriers presented significantly increased P. intermedia load [19]. Similarly, Stojanovska et al. demonstrated the association between P. intermedia and composite IL-1 genotype, but the significance was borderline (χ^2^ = 8.17; p = 0.06, p < 0.05). For the other tested periodontopathogens, there was no association between SNPs and bacteria loads [31]. Conversely, a case–control association study conducted on 415 northern European Caucasian patients with aggressive periodontitis and 874 healthy controls examined 10 SNPs in the genes of the IL1 cluster. The results showed no association between SNPs in the IL1 gene cluster and aggressive periodontitis [26]. Furthermore, Scapoli et al. also concluded that there is no association between generalized aggressive periodontitis and genetic variants in the IL1 gene cluster in an Italian Caucasian population [22]. Also, the Polish study did not find an association between periodontitis and IL-1A^−889^ and IL-1B^+3953^ TT genotype [35]. In our study, no statistically significant association was observed between four detected bacteria and genetic polymorphisms in IL-1B gene, which is shown in Table 4.
Considering high polymorphism within the IL-1 gene, there is large individual variability in cytokine production. In particular, the SNP at position +3954 in the IL-1B gene was studied, where the T allele could be associated with a higher risk of developing inflammatory diseases. However, the results of previous studies are contradictory, as some studies associate the C allele with the development of the disease, while others were unable to determine any association.
According to the Dahash and Kusrat study in the Iraqi population, there was a significant decrease in IL-B +3954 TT genotype in periodontitis cases compared to the control group (p = 0.018), indicating that this genotype is associated with the reduced risk for periodontitis, and CC genotype with an increased risk for the disease development [36]. Likewise, Sharma et al. found a significant association between IL-1B +3954 CC genotype and chronic periodontitis [37]. Results of our study indicate that the CC genotype in IL-1B gene could potentially be a genetic risk factor for higher loads of T. forsythia and P. gingivalis, as shown in Table 3.
The results of Pani et al. study indicate an independent association of IL-1B +3954 T allele carriers and red complex bacterial species with increased IL-1β levels in gingival crevicular fluid of periodontitis sites, but their study included a small number of participants from different ethnic groups [20]. Also, some individual case–control studies associated IL-1B +3954 C/T polymorphism with periodontitis. In a Brazilian study, Ribeiro et al. concluded that the IL-1B +3954 T allele represented risk for chronic periodontitis (OR = 2.84; 95% CI = 1.44–5.62), particularly in smokers (OR = 4.43; 95% CI = 1.29–15.24; p = 0.025) and females (OR = 6.00; 95% CI = 2.13–16.93; p = 0.027) [38]. Additionally, Ayazi et al., in their pilot study, found the significant association between distribution of IL-1B genotypes and the risk of periodontal disease (p = 0.017) [39]. Their data suggest that IL-1B +3954 C/T polymorphisms could be potential candidates for genetic biomarkers of periodontitis. Although these studies associated IL-1B +3954 TT genotype with a higher likelihood of periodontitis development, meta-analysis do not support these findings, as there was no association between IL-1B +3954 polymorphism and periodontitis susceptibility, regardless of ethnicity [40]. Furthermore, Linhartova et al. could not find any association between IL-1B polymorphisms and aggressive periodontitis among adult subjects from Czech Republic [14], and no association between IL-1B +3954 TT genotype was seen in Greek patients [41]. These results were in line with our findings, as there was no association between IL-1B +3954 polymorphisms and the presence of four detected bacteria, shown in Table 4.
Our findings also show no association between TNFA (−308 G/A) polymorphisms and tested periodontopathogenic bacteria. The lack of association between TNFA polymorphisms and periodontitis was reported by Yücel et al. [42] and Costa et al. [43]. Additionally, the results of a recent meta-analysis showed no significant associations of the TNFA (−308 G/A) polymorphism with aggressive periodontitis in Caucasians [44]. However, the results of a Serbian study indicate that homozygous carriers of the A allele are significantly more prone to periodontitis [45]. Conversely, Ianni et al. reported an association between TNFA (−308 G/A) GG genotype and increased risk of periodontitis [46].
While genetic variants in the TNFA gene in certain populations were associated with increased susceptibility to periodontitis, the allele and genotype frequencies vary among populations, making it difficult to generalize results and require population-specific research.
The findings of the present study should be interpreted considering both its strengths and limitations. A major strength of this study is that it is, to our knowledge, the first investigation conducted in a Croatian population to evaluate the association between IL-1A, IL-1B, and TNFA genetic polymorphisms and the presence of key periodontopathogenic bacteria, with sensitive detection by real-time PCR and adjustment for age, gender, smoking, and systemic disease.
Nevertheless, several limitations should be acknowledged. The relatively small sample size limits statistical power, while the cross-sectional design precludes causal inference. In addition, recruitment from a single center and the lack of periodontally healthy controls may restrict the generalizability of the findings to the broader population. Although bacterial copy numbers were quantified, the main analyses were based on the presence/absence of bacteria to ensure model stability, which may obscure biologically meaningful gradients of colonization. Furthermore, detailed quantitative periodontal parameters, such as clinical attachment level, bleeding on probing, plaque index, and pocket depth distribution, were not included in the multivariable models and may act as potential confounders. Finally, only selected cytokine polymorphisms were analyzed, and other genetic or environmental factors may also contribute to periodontal bacterial colonization. These limitations highlight the need for larger, well-powered studies incorporating healthy controls, quantitative bacterial outcomes, and comprehensive clinical parameters.
5. Conclusions
In this cohort from Split-Dalmatia County, IL-1A CC genotype was associated with increased likelihood of P. gingivalis and T. denticola colonization, whereas the IL-1B CC genotype was independently associated with T. forsythia detection. These findings suggest that host genetic variability in pro-inflammatory interleukin genes may influence subgingival bacterial colonization patterns. Conversely, the T allele may confer a protective effect, being associated with lower loads of periodontopathogenic species. However, these findings should be interpreted with caution given the study limitations. Larger, longitudinal studies including periodontally healthy controls, a priori power calculations, quantitative bacterial load modeling, comprehensive clinical periodontal indices, and appropriate multiple testing adjustments are warranted to confirm and extend these observations.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Hashim N.T. Babiker R. Padmanabhan V. Ahmed A.T. Chaitanya N. Mohammed R. Priya S.P. Ahmed A. El Bahra S. Islam M.S. The Global Burden of Periodontal Disease: A Narrative Review on Unveiling Socioeconomic and Health Challenges Int. J. Environ. Res. Public Health 20252262410.3390/ijerph 2204062440283848 PMC 12027323 · doi ↗ · pubmed ↗
- 2Hajishengallis G. Chavakis T. Local and systemic mechanisms linking periodontal disease and inflammatory comorbidities Nat. Rev. Immunol.20212142644010.1038/s 41577-020-00488-633510490 PMC 7841384 · doi ↗ · pubmed ↗
- 3Curtis M.A. Diaz P.I. Van Dyke T.E. The role of the microbiota in periodontal disease Periodontology 200083142510.1111/prd.1229632385883 · doi ↗ · pubmed ↗
- 4Kinane D.F. Stathopoulou P.G. Papapanou P.N. Periodontal diseases Nat. Rev. Dis. Primers 201731703810.1038/nrdp.2017.3828805207 · doi ↗ · pubmed ↗
- 5Holt S.C. Ebersole J.L. Porphyromonas gingivalis, Treponema denticola, and Tannerella forsythia: The “red complex”, a prototype polybacterial pathogenic consortium in periodontitis Periodontology 2000387212210.1111/j.1600-0757.2005.00113.x 15853938 · doi ↗ · pubmed ↗
- 6Lamont R.J. Koo H. Hajishengallis G. The oral microbiota: Dynamic communities and host interactions Nat. Rev. Microbiol.20181674575910.1038/s 41579-018-0089-x 30301974 PMC 6278837 · doi ↗ · pubmed ↗
- 7Fine D.H. Patil A.G. Velusamy S.K. Aggregatibacter actinomycetemcomitans (Aa) Under the Radar: Myths and Misunderstandings of Aa and Its Role in Aggressive Periodontitis Front. Immunol.20191072810.3389/fimmu.2019.0072831040843 PMC 6476972 · doi ↗ · pubmed ↗
- 8Talapko J. JuzbašićM. MeštrovićT. MatijevićT. MesarićD. KatalinićD. ErićS. Milostić-Srb A. Flam J. Škrlec I. Aggregatibacter actinomycetemcomitans: From the Oral Cavity to the Heart Valves Microorganisms 202412145110.3390/microorganisms 1207145139065217 PMC 11279289 · doi ↗ · pubmed ↗