The Role and Mechanisms of miRNAs on Ovarian Granulosa Cells: A Literature Review
Siyu Chen, Jiawei Lu, Yuqian Si, Lei Chen, Ye Zhao, Lili Niu, Yan Wang, Xiaofeng Zhou, Linyuan Shen, Ya Tan, Li Zhu, Mailin Gan

TL;DR
This review explores how microRNAs regulate ovarian granulosa cells and their role in conditions like PCOS and POF.
Contribution
The paper systematically integrates findings on miRNA mechanisms in granulosa cells across multiple models and diseases.
Findings
miRNAs regulate granulosa cell functions through key pathways like PI3K/AKT/mTOR and TGF-β/SMAD.
Exosomal miRNAs facilitate communication in the follicular environment.
Nearly 200 miRNAs are linked to PCOS, highlighting their potential as therapeutic targets.
Abstract
Background: Ovarian granulosa cells (GCs) play a pivotal role in folliculogenesis, and their dysfunction is central to disorders such as polycystic ovary syndrome (PCOS) and premature ovarian failure (POF). MicroRNAs (miRNAs) have emerged as crucial post-transcriptional regulators of GC homeostasis. Method: This review synthesizes current evidence by systematically analyzing relevant studies, integrating data from in vitro GC models, animal experiments, human cell lines, and clinical samples to elucidate the specific mechanisms by which miRNAs regulate GCs. Results: miRNAs precisely modulate GC proliferation, apoptosis, steroidogenesis, and oxidative stress responses by targeting key signaling pathways (e.g., PI3K/AKT/mTOR, TGF-β/SMAD) and functional genes (e.g., TP53, CYP19A1). Exosomal miRNAs serve as vital mediators of communication within the follicular microenvironment. To date,…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
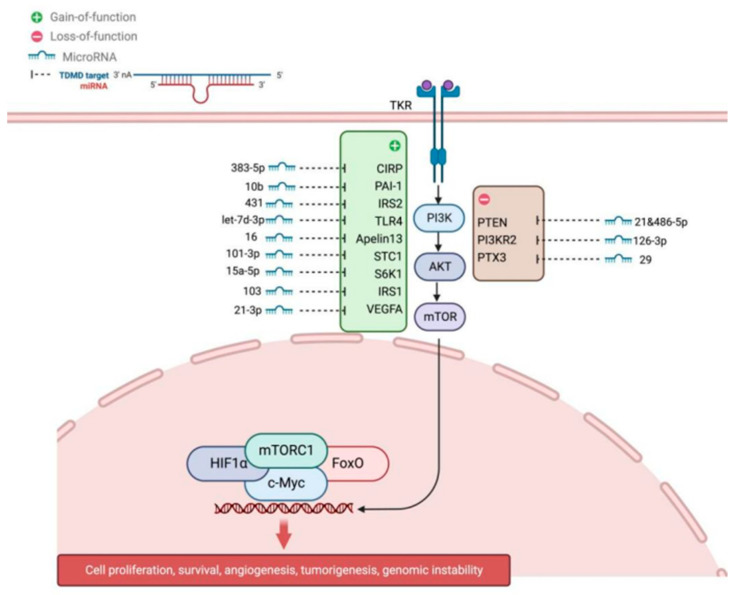 Figure 1
Figure 1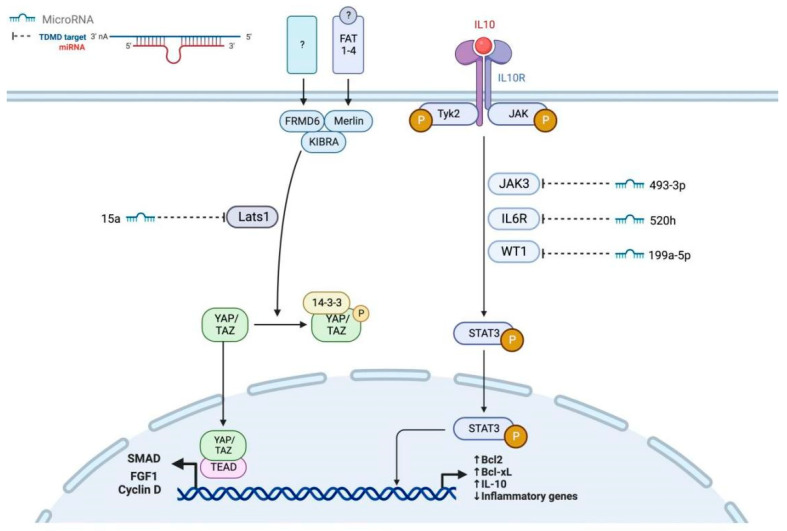 Figure 2
Figure 2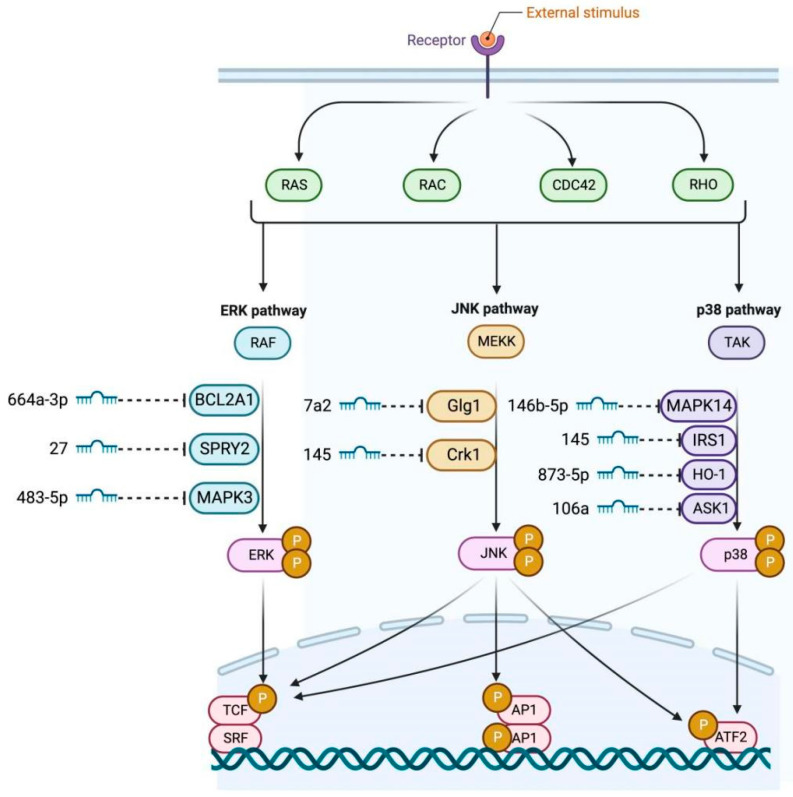 Figure 3
Figure 3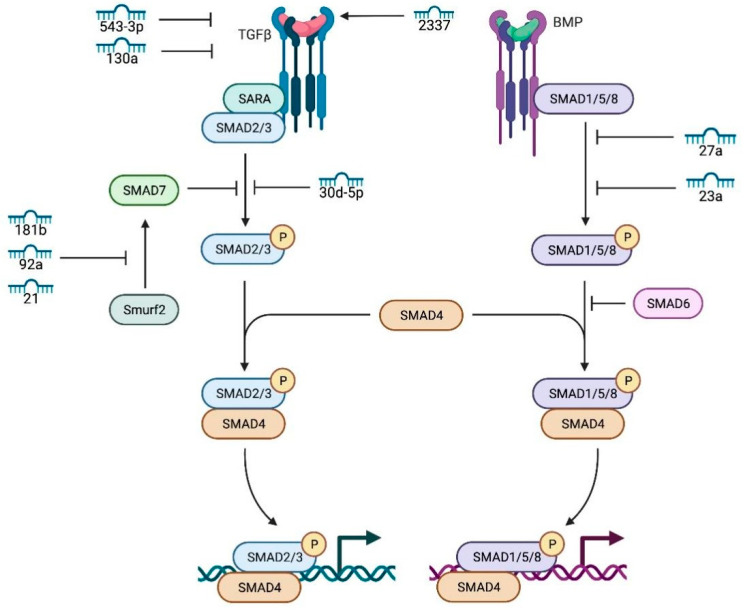 Figure 4
Figure 4| Target Gene/Pathway | Regulatory miRNA(s) | Effect on GCs | Associated Context | Reference |
|---|---|---|---|---|
| miR-644-5p (↑) | Inhibits apoptosis | POF therapy | [ | |
| miR-4110 (↓) | Promotes apoptosis via Sp1/p53 | [ | ||
| p53 → miR-27a (↓) | Promotes proliferation via NFAT5 | [ | ||
|
| miR-3188, miR-3940-5p (↑) | Promote proliferation, inhibit apoptosis | PCOS | [ |
| Mitofusins (Mfn1/2) | miR-214-3p (↑) | Inhibits proliferation & E2 synthesis | Porcine model | [ |
| IGF-1/ | miR-27a-3p, -130a, -99a, -323-3p, -19b (↓) | Mostly inhibit proliferation/promote apoptosis | PCOS models | [ |
|
| miR-107 (↑) | Suppresses proliferation, promotes apoptosis | Porcine GCs | [ |
|
| miR-17-5p (↑, via RNAa) | Inhibits HG-induced apoptosis | Sheep GCs | [ |
| ATF2 pathway | miR-451a (↑) | Modulates proliferation/apoptosis | Rat PCOS model | [ |
- —National The Natural Science Foundation of China
- —Sichuan Science and Technology Program
- —Postdoctoral Fellowship Program of CPSF
- —China Postdoctoral Science Foundation
- —China Agriculture Research System
- —Independent Research Fund of State Key Laboratory of Swine and Poultry Breeding Industry
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsReproductive Biology and Fertility · Ovarian function and disorders · MicroRNA in disease regulation
1. Introduction
Ovarian granulosa cells (GCs) are the essential somatic microenvironment for oocytes, governing folliculogenesis through metabolic support, steroid hormone synthesis, and precise regulation of proliferation and apoptosis. Dysregulation of GC homeostasis is a hallmark of polycystic ovary syndrome (PCOS) and premature ovarian failure (POF), which are disorders affecting millions of women worldwide [1]. The functional integrity of GCs determines follicular fate, with external stressors further compromising their function and highlighting their central importance in female reproductive health.
MicroRNAs (miRNAs), approximately 22-nucleotide non-coding RNAs, have emerged as dominant post-transcriptional regulators across biological systems. Recent genomic studies have expanded their functional repertoires dramatically: systematic re-analysis of 14,035 small-RNA-seq libraries uncovered >6000 newly annotated candidate loci, approximately doubling the searchable repertoire for granulosa-cell studies and revealing ovary-enriched clusters that had escaped canonical annotations [2]. Given this pivotal role in gene regulation and intricate link to reproductive pathologies, miRNAs have justifiably become a major research focus [3].
In the ovarian context, miRNAs serve dual functions: they regulate gene expression cell autonomously while also mediating intercellular communication via extracellular vesicles, particularly exosomes. These vesicles enable trafficking of functional miRNAs between cells within the follicular niche, with exosomal miRNAs derived from sources such as stem cells directly modulating GC gene expression and steroidogenic function [4]. This convergence of GC biology and miRNA mechanisms underscores a vital research frontier.
The clinical urgency is underscored by demographic trends: ovarian function declines with age, a process significantly accelerated by modern lifestyle factors such as circadian disruption and hypercaloric intake, which correlate with altered follicular miRNA profiles in young women. Accumulating evidence indicates that clinical control of follicular atresia, initiated by GC apoptosis, is crucial to alleviate diseases such as PCOS and POF. During gonadotropin-induced follicular development, GC mitochondria generate ATP via oxidative phosphorylation, while reactive oxygen species (ROS) production increases dramatically. Excessive ROS accumulation induces oxidative damage, mitochondrial DNA mutations, and lipid peroxidation, ultimately compromising GC-oocyte communication and oocyte quality [5].
This review systematically synthesizes current evidence on miRNA regulation of GC proliferation, apoptosis, steroidogenesis, and oxidative stress responses. By integrating data from in vitro GC cultures, animal models (mouse, rat, goat, sheep, pig, chicken), human granulosa-like tumor cell lines, and clinical samples—as exemplified by studies that map miRNA-mRNA interactomes in specific models [6]—we elucidate specific mechanisms by which miRNAs target key signaling pathways and functional genes. We emphasize that nearly 200 miRNAs are now associated with PCOS, yet the field faces significant translational challenges, primarily miRNA pleiotropy and the absence of follicle-specific delivery systems [3]. Most evidence derives from in vitro cultures or animal models, highlighting the critical need for validation in human-relevant systems such as organoids before miRNA-based strategies can be considered viable clinical tools for managing ovarian dysfunction. Deciphering the specific miRNA networks that control GC proliferation, apoptosis, and differentiation is therefore a research frontier of both fundamental and translational importance.
2. miRNAs Regulating Proliferation and Apoptosis of Ovarian Granulosa Cells
Post-transcriptional control by miRNAs constitutes a decisive layer governing granulosa-cell fate. The latest release of miRNA databases (e.g., mirDB 4.0) incorporates > 6000 newly annotated human miRNA loci—many enriched in ovarian clusters—providing an expanded repertoire for granulosa-cell-specific target prediction and functional annotation. Recent omics-level datasets now offer a systems-level view of these interactions. By integrating 12 shared miRNAs with matched mRNA profiles across two GC sub-populations, Yousuf et al. constructed a high-confidence miRNA–mRNA interactome that positions miR-129, miR-30a-5p, and miR-146a-5p as master regulators of proliferation vs. apoptosis hubs in human granulosa cells [6]. This extensive miRNA network directly targets key genes controlling cell survival, with major implications for follicular atresia and PCOS development [1,7].
2.1. miRNAs Targeting Core Proliferation and Apoptosis Genes
MiRNAs directly target key genes controlling cell survival, with major implications for follicular atresia and PCOS development. The tumor suppressor TP53 is a critical regulator of GC apoptosis, and several miRNAs modulate its expression. It has been demonstrated that miR-644-5p, delivered via bone mesenchymal stem cell (BMSC)-derived exosomes, regulates p53 to inhibit ovarian granulosa cell apoptosis, showing therapeutic potential for POF [8]. In contrast, Xie et al. found that miR-4110 increases Sp1 expression by suppressing Smad2, leading to p53 upregulation, increased Bax levels, and ultimately GC apoptosis [9]. Interestingly, p53 can also regulate miRNA expression: Liang et al. reported that p53 binds to the proximal promoter region of the GABAA receptor ε subunit of the miR-224 host gene, regulating miR-224 and SMAD4 expression, consequently affecting GC proliferation and estradiol release [10]. Additionally, Tao et al. showed that p53 inhibits miR-27a expression by targeting its promoter, resulting in upregulation of NFAT5 and promotion of mouse granulosa cell proliferation [11].
Potassium voltage-gated channel subfamily A member 5 (KCNA5) represents another important target. In PCOS, miR-3188 has been identified as a key miRNA that inhibits apoptosis and promotes GC proliferation through targeting KCNA5, with downstream upregulation of Bcl-2 and downregulation of Bax and Caspase-3 [12]. Similarly, miR-3940-5p was shown to target KCNA5, thereby promoting GC proliferation in PCOS [13].
Mitochondrial fusion proteins Mfn1 and Mfn2 are essential for female fertility. Hou et al. showed that Mfn1 deficiency leads to follicular development arrest and mitochondrial dysfunction in oocytes [14]. Wang et al. linked low Mfn2 expression in GCs with age and worse ART outcomes by impairing mitochondrial function and promoting apoptosis [15]. Shi et al. reported that miR-214-3p directly regulates Mfn2, inhibiting GC proliferation and estradiol synthesis [16]. These findings highlight mitochondrial fusion proteins as critical, though understudied, miRNA targets in GC biology.
Additional key functional genes are regulated by miRNAs. miR-107 suppresses porcine GC proliferation and estradiol synthesis while promoting apoptosis by targeting PTGS2 [17]. Wang et al. demonstrated that miR-17-5p transcriptionally activates KPNA2 expression via a non-canonical RNA activation mechanism, counteracting high-glucose-induced oxidative stress and apoptosis in sheep granulosa cells [18]. The insulin-like growth factor pathway is also heavily targeted: The IGF-1 pathway is heavily targeted by miRNAs in various contexts. For instance, exposure to bisphenol A can dysregulate the IGF-1/miR-27b-3p axis [19], while in PCOS, mechanisms such as lncRNA HCP5 competitively binding miR-27a-3p [20] or miR-130a directly reducing IGF-1 expression have been reported [21]; Geng et al. demonstrated that miR-99a regulates proliferation and apoptosis of human GCs by targeting IGF1R [22]; Wu et al. found that miR-323-3p regulates steroidogenesis and apoptosis in KGN cells by targeting IGF-1 [23]; Zhong et al. showed that inhibition of miR-19b promotes GC proliferation by targeting IGF-1 in PCOS [24]; and Yang et al. revealed that miR-451a targets the ATF2 signaling pathway, modulating GC proliferation and apoptosis in a rat PCOS model [25]. Key miRNAs and their target genes discussed above are summarized in Table 1.
2.2. miRNAs Regulating Signaling Pathway Activity
MiRNAs precisely modulate core signaling cascades, with pathway-level integration determining GC fate decisions.
2.2.1. PI3K/AKT/mTOR Signaling Pathway
The PI3K/AKT/mTOR axis is central to GC proliferation and follicle dominance. In PCOS, miR-let-7d-3p inhibits the PI3K/Akt pathway and GC proliferation by targeting TLR4 [23]. Guo et al. demonstrated that miR-10b promotes apoptosis by targeting PAI-1 to regulate the PI3K/AKT pathway [24]. An et al. showed that miR-101-3p inhibits PI3K/AKT/mTOR by targeting STC1 in goat ovarian development [26]. High levels of miR-15a-5p correlate with poor ovarian response, likely via inhibiting granulosa cell proliferation through this pathway [27]. Yang et al. reported that miR-431 inhibits GC proliferation by targeting IRS2 and suppressing PI3K/AKT signaling [28]. Wang et al. found that miR-29 regulates goat GC function by targeting PTX3, activating both PI3K/AKT/mTOR and Erk1/2 pathways [29]. Han et al. demonstrated that miR-21 in oocyte secretory factors inhibits PI3K/Akt signaling and promotes GC apoptosis [30]. Recent work in sheep showed that miR-134-3p suppresses proliferation and induces apoptosis by targeting INHBA and dampening the TGF-β/PI3K/AKT cascade [31]. Clinically, Han et al. discovered that PD-L1 upregulation in PCOS suppresses GC apoptosis by activating PI3K/AKT, highlighting a potential immune-endocrine axis [32]. The miRNA-mediated regulation of PI3K/AKT/mTOR signaling is depicted in Figure 1.
Beyond the well-characterized miRNA regulators, emerging evidence from high-throughput screening has identified additional miRNA-target interactions that modulate PI3K/AKT/mTOR signaling, though their functional roles in granulosa cells remain to be fully validated. These include miR-126-3p targeting PIK3R2 (the regulatory subunit of PI3K), miR-103 directly regulating mTOR expression, and miR-21-3p controlling VEGFA-mediated angiogenic signals. A study in endometrial cancer cells demonstrated that miR-126-3p directly binds the 3′UTR of PIK3R2, reducing PI3K activity and downstream AKT phosphorylation [33]. Similarly, miR-103 has been shown to suppress mTOR expression in hepatocellular carcinoma, influencing cell metabolism and proliferation [34]. While these specific interactions have not yet been confirmed in ovarian granulosa cells, the conserved nature of miRNA targeting suggests they may represent novel regulatory nodes in follicular development. Additionally, the cold-inducible RNA-binding protein (CIRP) has been identified as a target of miR-383-5p in stress responses [35], and the tumor suppressor PTEN is co-targeted by miR-21 and miR-486-5p within the PI3K/AKT/mTOR network [36,37], offering potential new avenues for investigating granulosa cell metabolism and survival.
2.2.2. JAK/STAT and Hippo Signaling Pathways
The JAK/STAT pathway is essential for cytokine signaling in GCs. Frost et al. reported conserved roles in human follicle development [38]. Hall et al. confirmed PI3K/AKT and JAK/STAT signaling as conserved biomarkers for ovarian development [39]. Yang et al. showed that IL-6 promotes FSH-induced VEGF expression through JAK/STAT3 in bovine GCs [40]. Ndiaye et al. identified novel JAK3-interacting proteins in GCs, including LEPROTL1, INHBA, and CDKN1B [41]. The highly expressed miR-199a-5p in PCOS patients inhibits WT1-mediated JAK/STAT3 activation, and its own inhibition has been shown to improve PCOS pathology [42].
The Hippo pathway is emerging as a critical regulator. Ai et al. determined that miR-15a targets Lats1 to inhibit GC proliferation and induce senescence. Recent studies expand Hippo crosstalk: Wang et al. demonstrated that RSPO2 coordinates with GDF9:BMP15 heterodimers to promote GC development via Wnt/β-catenin signaling, which extensively crosstalks with Hippo [43]. Shan et al. identified miR-184 as a transcriptional activator that acts as a transcriptional enhancer that indirectly induces SMAD3 expression by targeting an inhibitory factor, potentially interfacing with Hippo through TGF-β crosstalk [44]. The cytokine sensitivity of granulosa cells is further fine-tuned by miRNAs targeting receptor components of the JAK/STAT pathway. Recent transcriptomic analyses have identified miR-520h as a post-transcriptional regulator of IL6R expression, modulating granulosa cell responsiveness to IL-6-mediated signals [45]. Similarly, miR-493-3p has been implicated in controlling MIF (macrophage migration inhibitory factor) expression, thereby influencing macrophage infiltration and inflammatory responses [46]. While these specific receptor-targeting mechanisms have been characterized primarily in immune cells, their presence in ovarian granulosa cell transcriptome datasets suggests they may play important roles in regulating the follicular immune microenvironment during folliculogenesis and atresia. Validation of these interactions in granulosa cell-specific models represents an important area for future investigation. Taken together, these recent studies highlight that the Hippo pathway does not act in isolation; rather, through extensive crosstalk with other key pathways such as Wnt/β-catenin and TGF-β, it forms an integral part of the complex network determining granulosa cell fate. The miRNAs regulating JAK/STAT3 and Hippo signaling pathways are illustrated in Figure 2.
2.2.3. MAPK Signaling Pathway
The MAPK pathway is a major signal transducer in GCs. miR-664a-3p inhibits KGN cell proliferation and promotes apoptosis by targeting BCL2A1 and consequently blocking MAPK/ERK signaling [47]. Liu et al. found that miR-146b-5p overexpression attenuates premature ovarian failure in mice by inhibiting the Dab2ip/Ask1/p38MAPK pathway [7]. Hong et al. showed that miR-106a increases GC viability by inhibiting ASK1 mRNA and p38MAPK phosphorylation [48]. Cai et al. proposed that miR-145 inhibits MAPK/ERK by targeting IRS1 in PCOS [49]. Wang et al. found that miR-145 targets Crkl to promote GC proliferation, differentiation, and steroidogenesis via JNK/p38MAPK [50]. The MAPK pathway also interfaces with less-characterized miRNA-target pairs that may have granulosa cell-specific functions. The diagram depicts miR-146b-5p targeting Golgi glycoprotein 1 (Glg1), a protein involved in vesicular trafficking and secretion [51]. Additionally, miR-873-5p is shown to regulate heme oxygenase-1 (HO-1), an enzyme critical for cellular stress response and antioxidant defense [52]. While these interactions have been documented in cancer and endothelial cell lines, their roles in granulosa cell oxidative stress responses during follicular development remain unexplored. The presence of miR-483-5p in the pathway network without an explicit target suggests additional regulatory complexity that warrants further investigation. These emerging miRNA-target pairs highlight knowledge gaps that must be addressed to fully understand MAPK-mediated control of granulosa cell fate. The miRNAs regulating the MAPK signaling pathway are illustrated in Figure 3.
2.2.4. TGF-β/SMAD Signaling Pathway
TGF-β/SMAD signaling is fundamental to female fertility. miR-543-3p targets decorin (DCN), leading to reduced expression of TGF-β pathway-related genes [53]. Du et al. reported that miR-130a promotes GC apoptosis by inhibiting TGF-β1 expression [54]. Wang et al. identified miR-2337 as a small activating RNA that enhances TGF-β1 promoter activity [55]. Several miRNAs target SMAD proteins: Nie et al. showed miR-23a and miR-27a promote apoptosis by targeting SMAD5 [56]; Yu et al. found miR-30d-5p targets SMAD2, inhibiting proliferation and promoting apoptosis [57]; Yao et al. showed miR-181b-induced SMAD7 downregulation controls GC apoptosis [58]; Liu et al. reported miR-92a inhibits apoptosis by targeting SMAD7 [59]; Yin et al. demonstrated that miR-224 targets SMAD4, regulating GC proliferation and estradiol production [60].
TGF-β1 also trans-regulates miRNA expression: Du et al. found that TGF-β1 induces miR-764-3p expression, which enhances GC proliferation via SMAD4 while inhibiting estradiol synthesis via SF-1 [61]. Yin et al. showed miR-383 transactivates miR-320, inhibiting estradiol synthesis and proliferation by targeting E2F1 and SF-1 [62]. Importantly, SMAD4 activates Wnt signaling to inhibit GC apoptosis [61] and represses multiple miRNAs that negatively regulate TGF-β signaling [63]. Recent evidence identifies the TGF-β1–SMAD4–BMF module as a conserved pro-survival axis in ruminant and human GCs [64,65,66]. The TGF-β/SMAD network also includes miRNAs with less-defined roles in granulosa cells. Figure 4 places miR-33 within the pathway without specifying a direct molecular target, suggesting it may function as a fine-tuner of overall pathway activity rather than a single-gene regulator. Studies in ovarian granulosa cells indicate that miR-33 can modulate TGF-β signaling by targeting key components of the pathway. For instance, in the context of polycystic ovary syndrome, miR-33b is upregulated and directly targets the TGF-β receptor TGFBR1, thereby influencing the signaling cascade and cellular outcomes [67]. In the ovarian context, miR-33 expression has been detected in follicular fluid exosomes, though its specific granulosa cell targets remain uncharacterized. Elucidating the mechanism by which miR-33 interfaces with the TGF-β/SMAD cascade could reveal novel regulatory nodes controlling follicular selection and atresia.
2.3. MiRNAs Regulating Steroidogenesis and Oxidative Stress Homeostasis
2.3.1. Steroid Hormone Production
MiRNAs critically regulate estrogen synthesis by targeting CYP19A1 (aromatase) and CYP11A1. Several miRNAs directly target CYP19A1: miR-210, miR-32, miR-146b, miR-10b, and miR-378 reduce estradiol secretion and promote apoptosis [68,69,70]. Others target transcriptional regulators: miR-29a impairs CREB1 to inhibit CYP19A1 [71]; Ding et al. showed miR-202-5p transactivated by SF-1 targets TGFβR2 [72]; Zhang et al. found miR-17-5p targets E2F1 [73]; Liu et al. reported miR-1275 impairs LRH-1/CYP19A1 axis [74]; Wang et al. showed miR-27a-3p targets CREB1 [75]; and Du et al. found miR-764-3p targets SF-1 [76]. Circular RNAs also participate: Zhang et al. identified hsa_circ_0043532 upregulating CYP19A1 by acting as a ceRNA for miR-1270 [77], while Tao et al. revealed the KRTAP5-AS1/miR-199b-5p/CYP19A1 axis [78]. Conversely, some miRNAs promote steroidogenesis: Zhang et al. showed miR-320a targets RUNX2 to enhance steroidogenesis [79]; Wu et al. found miR-132 targets Nurr1 [80]; and Dai et al. demonstrated miR-133b targets Foxl2 [81]. A 2024 study in chickens showed miR-7 promotes apoptosis and autophagy by targeting KLF4 via JAK/STAT3 [82].
2.3.2. Reactive Oxygen Species (ROS) Balance
MiRNAs regulate ROS metabolism, critical for GC survival. Liu et al. reported that miR-27 increases ROS stress and apoptosis by targeting SPRY2 and downregulating the p-ERK/Nrf2 pathway [83]. Ding et al. found hAMSC exosome-derived miR-320a targeting SIRT4 reduces ROS production [84], while hUMMSC exosome-derived miR-17-5p targeting SIRT7 exerts similar effects [85]. FOXO1 is a key mediator: it induces TXNIP expression, activates NLRP3 inflammasomes, causing oxidative stress and apoptosis [86]. Several miRNAs target FOXO1: Yan et al. found miR-1224-5p reduces PCOS by inhibiting NLRP3 activation via FOXO1 [87]; Wei et al. showed miR-221-3p reduces ovarian reserve by targeting FOXO1 in elderly women [88]; and Zhang et al. found miR-181a increases FOXO1 acetylation and promotes apoptosis through SIRT1 downregulation [77]. A 2025 study demonstrated that miR-361-5p alleviates GC dysfunction in diminished ovarian reserve by targeting SLC25A24 to maintain mitochondrial function [89].
3. Conclusions and Future Perspectives
This review establishes miRNAs as a decisive regulatory network governing GC proliferation, apoptosis, steroidogenesis, and oxidative stress responses. Integrated analyses identify miR-129, miR-30a-5p, and miR-146a-5p as master regulators in human GCs [6], while the TGF-β1–SMAD4–BMF module represents a conserved pro-survival axis across species [64,65,66]. MiRNAs target core components of PI3K/AKT/mTOR, TGF-β/SMAD, JAK/STAT, MAPK, and Hippo pathways, as well as key functional genes including TP53, BCL2 family members, KCNA5, Mfn1/2, and steroidogenic enzymes CYP19A1/CYP11A1.
However, significant translational challenges remain. First, miRNA pleiotropy poses off-target risks—miR-21 alone has four validated targets (PTEN, SMAD7) in PCOS [90,91]. Second, the vast majority of evidence derives from in vitro cultures or animal models, creating a substantial translational gap [1]. Third, the absence of follicle-specific delivery systems limits therapeutic applicability. Recent advances in tissue-specific miRNA delivery using cationic polymer nanocomplexes and peptide-modified AAV variants offer promise, but require further optimization for ovarian applications [3].
Future research must prioritize validation in human-relevant models. The development of human “ovaroid” organoids from induced pluripotent stem cells without exogenous transcription factors represents a crucial step toward bridging this gap [92]. Single-cell and spatial transcriptomics will further refine our understanding of cell type-specific miRNA functions within the follicular niche [2,6]. Addressing these challenges is paramount before miRNA-based strategies can be translated into viable clinical tools for managing PCOS, POF, and other ovarian disorders.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Yi Y. Zhong Y. Liang R.N. Liu C.Y. Zhang Y.F. Zhang Y.X. Polycystic ovary syndrome: The role of granulosa cell proliferation and apoptosis in disease development J. Steroid Biochem. Mol. Biol.202525410683810.1016/j.jsbmb.2025.10683840721086 · doi ↗ · pubmed ↗
- 2Gao F. Wang F. Chen Y. Deng B. Yang F. Cao H. Chen J. Chen H. Qi F. Kapranov P. The human genome encodes a multitude of novel mi RN As Nucleic Acids Res.202553 Gkaf 07010.1093/nar/gkaf 07039964476 PMC 11833695 · doi ↗ · pubmed ↗
- 3Liu L. Fang Y. The Role of Ovarian Granulosa Cells Related-nc RN As in Ovarian Dysfunctions: Mechanism Research and Clinical Exploration Reprod. Sci.2025322098212010.1007/s 43032-025-01854-240175717 PMC 12271301 · doi ↗ · pubmed ↗
- 4Cai J.H. Sun Y.T. Bao S. Huc MS Cs-exosomes containing mi R-21 promoted estrogen production in ovarian granulosa cells via LATS 1-mediated phosphorylation of LOXL 2 and YAP Gen. Comp. Endocrinol.2022321–32211401510.1016/j.ygcen.2022.11401535271888 · doi ↗ · pubmed ↗
- 5Lin L. Gao W. Chen Y. Li T. Sha C. Chen L. Yang M. Wei H. Chen Y. Zhu X. Reactive oxygen species-induced SIAH 1 promotes granulosa cells’ senescence in premature ovarian failure J. Cell. Mol. Med.2022262417242710.1111/jcmm.1726435261172 PMC 8995443 · doi ↗ · pubmed ↗
- 6Yousuf S. Malik W.A. Feng H. Liu T. Xie L. Miao X. Integrated analysis of micro RNA and m RNA interactions regulating fecundity in the ovaries of two distinct sheep breeds BMC Genom.20252670710.1186/s 12864-025-11408-0PMC 1231232440745523 · doi ↗ · pubmed ↗
- 7Liu T. Lin J. Chen C. Nie X. Dou F. Chen J. Wang Z. Gong Z. Micro RNA-146b-5p overexpression attenuates premature ovarian failure in mice by inhibiting the Dab 2ip/Ask 1/p 38-Mapk pathway and γH 2A.X phosphorylation Cell Prolif.202154 E 1295410.1111/cpr.1295433166004 PMC 7791167 · doi ↗ · pubmed ↗
- 8Sun B. Ma Y. Wang F. Hu L. Sun Y. mi R-644-5p carried by bone mesenchymal stem cell-derived exosomes targets regulation of p 53 to inhibit ovarian granulosa cell apoptosis Stem Cell Res. Ther.20191036010.1186/s 13287-019-1442-331783913 PMC 6884862 · doi ↗ · pubmed ↗