NKGD2 Ligands (NKG2DLs) in Breast Cancer: In Silico Analysis and Narrative Review
Jesús Peña-López, Angelo Gámez-Pozo, Lucía Trilla-Fuertes, Fernando Becerril-Gómez, Marta Mendiola, Victoria Heredia, Laura Yébenes, Beatriz Castelo, Virginia Martínez-Marín, Enrique Espinosa, Pilar Zamora, Alfonso Alba-Bernal, Cristina Aguirre-Portolés, Antonio Pérez-Martínez

TL;DR
This paper explores the role of NKG2D ligands in breast cancer using in silico analysis and a literature review, highlighting their potential as immunotherapy targets.
Contribution
The study provides a comprehensive in silico and narrative analysis of NKG2D ligands in breast cancer, revealing their expression patterns and therapeutic relevance.
Findings
NKG2DL transcripts are consistently expressed in breast cancer cell lines and tumor tissues.
Higher NKG2DL expression is associated with ductal histology, advanced tumor stage, and basal molecular subtype.
NKG2DLs are detectable in tumor tissues but not in surrounding healthy tissues according to immunohistochemistry.
Abstract
Breast cancer (BC) is a global health problem. BC is a biologically heterogeneous disease in which novel immunotherapeutic strategies are needed, particularly in the metastatic setting. The NKG2D/NKG2D ligand (NKG2DL) axis is a key component of innate antitumor immunity and represents a potential therapeutic target, but its relevance in BC has not been fully characterized. We performed an in silico analysis of NKG2DL expression in BC cell lines, healthy breast tissue, and tumor samples using publicly available transcriptomic databases (DSMZCellDive, ShinyTHOR, GTEx, TCGA, Human Protein Atlas), complemented by survival analyses from TCGA and KMPlot and a narrative review of the literature. NKG2DL transcripts were consistently expressed in BC cell lines and tumor tissues, with higher expression observed in ductal histology, higher tumor stage, and basal molecular subtype. Survival…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
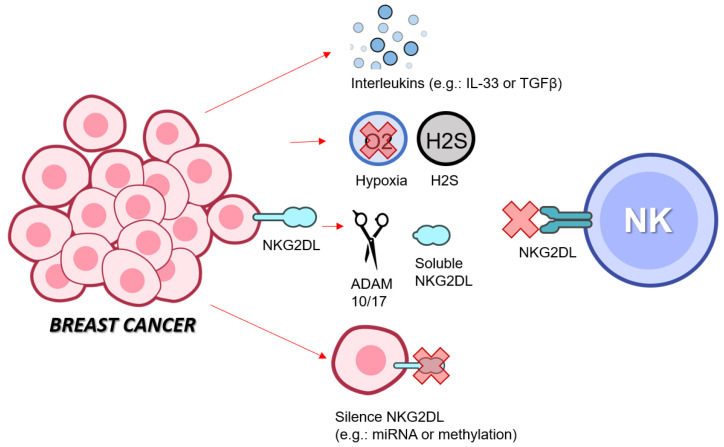 Figure 6
Figure 6Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsImmune Cell Function and Interaction · Glycosylation and Glycoproteins Research · Immunotherapy and Immune Responses
1. Introduction
Breast cancer (BC) is a global health issue. In 2022, it was the second most frequently diagnosed cancer, accounting for around 2.3 million new cases or approximately one in nine cancer diagnoses worldwide (11.6% of all cancers). It is also the fourth largest cause of cancer-related mortality, responsible for an estimated 0.6 million deaths in 2022 (6.9% of all deaths from cancer). In women, it is the most common tumor and the leading cause of cancer mortality [1]. BC is a biologically heterogeneous disease, with different molecular subtypes with prognostic and therapeutic implications. The triple-negative subtype (TNBC) is particularly important due to its greater aggressiveness, poorer prognosis, and fewer treatment options [2]. Perou et al. established four molecular intrinsic subtypes based on a 50-gene expression signature (PAM50): Luminal A, Luminal B, HER2-enriched and basal-like [3]. In clinical practice these subtypes are subrogated through immunohistochemistry (IHC) of four key biomarkers: estrogen receptor (ER), progesterone receptor (PR), human epidermal growth factor receptor 2 (HER2/ERBB2) and Ki67 (proliferation marker). Depending on the expression of these proteins, tumors are classified as: luminal or hormone receptor-positive (ER+/PR+), HER2-positive (HER2+) or triple-negative (ER-/PR-/HER2-) [2]. There is approximately 80% concordance between the basal molecular subtype and TNBC surrogate subtype [4].
There have been multiple therapeutic innovations in metastatic BC. Traditionally, there was classic chemotherapy, hormone therapy (for luminal tumors), and anti-HER2 targeted therapy (for HER2-positive tumors). However, new treatments have been introduced in recent years:
- Immune checkpoint inhibitors have been incorporated in triple-negative cases if PD-L1 (CD274) is positive (around 40% of the trial population) [5].
- Antibody–drug conjugates (ADCs). Sacituzumab–govitecan (anti-Trop2/TACSTD2) has demonstrated benefits in TNBC and luminal BC [6,7]. Trastuzumab–deruxtecan (antiHER2) has not only demonstrated benefits in HER2-positive tumors, but also in HER2-low tumors (expression present but insufficient to meet the classic criteria for positivity) [8,9]. Others, such as enfortumab–vedotin (anti-Nectin4), are currently being studied [10].
Nevertheless, metastatic BC is incurable in practically most cases and the aim of systemic cancer-specific treatment is palliative [11]. Overall survival rates in the metastatic setting achieved in the luminal and HER2 subtypes are around 5 years, while in TNBC the survival rate is around 1–2 years [2].
The Natural Killer Group 2 member D (NKG2D) receptor plays an important role in protecting the host against infections and cancer. It is constitutionally expressed in γδ T lymphocytes, CD8+ αβ T lymphocytes, and NK cells. It modulates the activation of these lymphocytes by recognizing ligands on target cells such as major histocompatibility complex class I-related proteins A and B (MICA and MICB) and UL16 binding proteins 1–6 (ULBP1, ULBP2, ULBP3, ULBP4/RAET1E, ULBP5/RAET1G and ULBP6/RAET1L). The expression of these ligands can be induced in cells that are infected, damaged by DNA, undergoing carcinogenesis, or stressed by other inducers. Thus, cellular immunity is promoted to eliminate cells that express the ligand [12,13]. These ligands are not widely expressed on healthy adult tissue so NKG2D ligands may present a useful target for immunotherapeutic approaches in cancer [14].
The NKG2D/NKG2DL axis is being targeted in pediatric leukemia with chimeric antigen receptor (CAR) anti-NKG2DL or with ex vivo IL-15-stimulated NK cells [15,16]. The question arises as to whether it is possible to transfer this approach to BC. Notably, early-phase clinical trials are being conducted with CAR-based adoptive cell therapy against NKG2DLs in solid tumors, including BC [17]. To assess the feasibility of this strategy, there are two aspects to consider. First, it is necessary to determine whether NKG2DL expression exists through in silico analysis in different databases. Second, the available evidence regarding BC (especially TNBC) and NKG2DLs must be analyzed.
2. In Silico Analysis
2.1. In Silico Analysis of Cell Lines
DSMZCellDive [https://celldive.dsmz.de/ (accessed on 23 December 2025)] contains RNA sequencing (RNA-seq) transcriptome data (expressed as TPM, transcripts per million) of different immortalized human cell lines (breast cancer n = 29, leukemia/lymphoma n = 101, neuroblastoma n = 18 and retinoblastoma n = 9) (Table S1) [18]. We focused specifically on NKG2DL genes and breast cancer cell lines (Table S2) [19]. In the case of breast cancer, the values for CD274, TACSTD2, ERBB2, and NECTIN4 (genes currently being targeted) were added.
The expression of NKG2DLs in the different human cell lines (grouped by tumor) is shown in Figure A1 and Figure A2. The data obtained are illustrated in Table 1 and Figure 1. When specifically analyzing BC cell lines, no statistically significant differences in gene expression were found when stratified by primary vs. metastatic origin or by luminal vs. basal subtype.
ShinyTHOR [https://alexismurillo.shinyapps.io/ShinyThor/ (accesed on 6 February 2026)] is a web app for access to multi-omic data from different datasets (CCLE, miRTarBase, circInteractome, and the Genomics of Drug Sensitivity in Cancer) [20]. This allowed us to create a heatmap of NKG2DL expression in 60 immortalized human BC cell lines (Figure A3).
2.2. In Silico Analysis of Healthy Tissue
The Adult Genotype Tissue Expression (GTEx) Project [http://www.gtexportal.org/home/ (accessed on 23 December 2025)] collected samples from up to 54 non-diseased tissue sites across nearly 1000 deceased individuals [21]. It was combined with RNA-seq transcriptome data of NKG2DL genes (collected as nTPM, normalized transcripts per million) present in The Human Protein Atlas [https://www.proteinatlas.org/ (accessed on 23 December 2025)] [22]. From this, we selected the 514 breast samples (Table S3).
The expression of NKG2DLs in healthy breast tissues from the GTex database is illustrated in Table 2. The heatmap of NKG2DL expression in different healthy tissues is shown in Figure A4.
The Human Protein Atlas was also used to evaluate the protein expression of ULBP1, ULBP2, and ULBP3 (the only ones for which information was available). No protein expression was detected in ULBP1 and ULBP2. “Low” protein expression of ULBP3 was detected in the glandular component.
2.3. In Silico Analysis of Tumor Tissue
The Cancer Genome Atlas (TCGA) breast cancer database, contained in cBioportal [http://www.cbioportal.org/ (accessed on 23 December 2025)], includes clinical–pathological information from 1084 breast cancer samples [23,24,25,26]. It was combined with RNA-seq transcriptome data of NKG2DL genes (collected as pTPM, protein-transcripts per million) present in The Human Protein Atlas [https://www.proteinatlas.org/ (accessed on 23 December 2025)] [22]. Then, we selected the 1009 samples from patients diagnosed with invasive breast carcinoma (Table S4). In the case of breast cancer, the values for CD274, TACSTD2, ERBB2, and NECTIN4 (genes currently being targeted) were added.
Figure A4, obtained from The Human Protein Atlas, shows the expression of NKG2DLs in different tumors (using TCGA data). The baseline characteristics of the 1009 BC patients are summarized in Table 3. Figure 2 shows the differences in expression by histology (ductal vs. lobular), tumor stage code (T1 vs. T2–4), and nodal stage code (N0 vs. N+). Ductal histology is associated with higher NKG2DL expression (except MICB). T ≥ 2 is associated with higher ULBP1-ULBP2-ULBP3 expression. In N+, higher expression was only detected in ULBP1.
Figure 3 shows the differences in expression by molecular subtype. There are significant differences in ULBP1, ULBP2, ULBP3, RAET1G, and RAET1L. In these cases, the basal subtype has higher levels than the other subtypes (significance is not detected only versus HER2 for ULBP2 and RAET1G).
The Human Protein Atlas was also used to evaluate the protein expression of ULBP1, ULBP2, and ULBP3 (the only ones for which information was available). In ULBP1, “medium” protein expression was recorded in 1/11 samples. In ULBP3, “medium” protein expression was recorded in 1/11 samples. In ULBP2, “weak” staining was recorded in <25%.
With the TCGA data, a survival analysis (estimated using the Kaplan–Meier method) could be performed. The median follow-up was 31.4 months (95%CI: 28.6–34.3) and death had occurred in 141 patients (14.0%). The median overall survival (mOS) was 132.0 months (95%CI: 115.3–148.7).
A univariate analysis was conducted using the Cox regression model to correlate overall survival (OS) with NKG2DL RNA expression (“low” and “high” groups were created using the median expression as the cut-off point) or clinical–pathological factors. No significant differences were found in mOS based on levels of MICA (p = 0.402), MICB (p = 0.202), ULBP1 (p = 0.081), ULBP2 (p = 0.215), ULBP3 (p = 0.647), RAET1E (p = 0.165), and RAET1L (p = 0.166). Significant differences were found in RAET1G (low vs. high; 131.5 months vs. 132.0 months; p = 0.044; HR = 1.4, 95%CI: 1.01–1.96; Figure 4). In relation to clinical–pathological factors, no differences were found in terms of histology (ductal vs. lobular; p = 0.510). Differences were found in tumor stage code (T1 vs. T2–4; p = 0.026), nodal stage code (N0 vs. N+; p < 0.001), and molecular subtype (p = 0.008).
The significant variables in the univariate model were incorporated into a multivariate model. The independent prognostic factors identified were molecular subtype Luminal A (p = 0.020, aHR = 0.58, 95%CI: 0.37–0.92) and N+ (p = 0.001, aHR = 1.91, 95%CI: 1.31–2.80).
A similar analysis was performed using another database, The Kaplan–Meier (KM) Plotter Online Tool [https://www.kmplot.com/analysis/ (accessed on 23 December 2025)]. It has RNA-seq data on 2976 breast cancer patients obtained from the GEO and EGA repositories [27]. It allowed us to analyze the relationship between the relevance of different NKG2DL RNA expressions (“low” and “high” groups were created using the median expression as the cut-off point) and patients’ clinical outcomes (OS). No significant differences were found in mOS when stratified by low–high levels of MICA (p = 0.48), MICB (p = 0.70), ULBP3 (p = 0.62), RAET1G (p = 0.29) and RAET1L (p = 0.75). Significant differences were found in ULBP1 (p = 0.001; HR = 1.9, 95%CI: 1.5–2.4), ULBP2 (p = 0.013; HR = 1.3, 95%CI: 1.1–1.7) and RAET1E (p = 0.044; HR = 1.26, 95%CI: 1.01–1.6) (Figure 5). When specifically analyzing the TNBC subtype (n = 126) and the basal subtype (n = 309), no significant differences in mOS were found based on NKG2DL expression.
3. Narrative Review
We conducted a keyword-search-based literature review using PubMed. The following output was introduced in December 2025: (“breast neoplasms”[MeSH Terms] OR BREAST CANCER[Text Word] AND NKG2D) OR (“breast neoplasms”[MeSH Terms] OR BREAST CANCER[Text Word] AND NKG2DL). A total of 114 studies were obtained and their distribution over time was recorded (Table S5).
3.1. Potential Resistance Mechanisms
The mechanisms by which each cancer (not only BC) can evade the NKG2D system are numerous and complex. Table 4 summarizes the available evidence.
3.2. Findings in Breast Cancer Patients
Over the past 20 years, data on NKG2DL in patients with BC has been collected. Table 5 summarizes the available evidence.
3.3. Activating Mechanisms
Various mechanisms are being studied to improve the function of the NKG2D system. Table 6 shows the available evidence.
4. Discussion
In this study, we reviewed the published evidence and in silico data available on NKG2DL in BC. At the transcriptional level, NKG2DL is expressed in BC cell lines and tumor samples. This expression is robust compared to other tumor types. However, expression levels are lower than those of some genes whose targets are currently used in the treatment of metastatic BC, such as TACSTD2 or ERBB2. Nevertheless, we have the case of CD274 (which also participates in the immune response) that resembles that of NKG2DL. It should be noted, however, that in metastatic TNBC, immune checkpoint inhibitors only work in a selected PD-L1-positive population [5], so we wonder whether future therapies against the NKG2D/NKG2DL axis will also need to be selected based on expression. One way to stratify indirectly is to see which subpopulations have higher levels of NKG2DL. Using TCGA data, we see that expression is generally higher in ductal histology, T ≥ 2, and basal molecular subtype. This last molecular aspect could support the idea that the NKG2D/NKG2DL axis could be a target in TNBC.
An important aspect is that in silico studies of healthy breast tissue also show NKG2DL mRNA expression. This could lead to problems in terms of “on target, off tumor” toxicities. However, the expression of RNA transcripts does not necessarily imply effective protein expression. There is evidence that healthy tissues can inhibit NKG2DL protein expression despite mRNA transcripts existence through post-transcriptional mechanisms (e.g., miRNA) or post-translational mechanisms (e.g., preventing protein translocation from the endoplasmic reticulum to the cell surface) [133]. However, the in silico data we have on protein expression to clarify this is scarce. Therefore, to determine the evidence of NKG2DL protein expression in BC, we conducted a narrative review in PubMed. As previously mentioned, there is NKG2DL protein expression in BC tumor cells, while immunohistochemical staining in non-tumor border tissues is low. This supports the use of NKG2DL as a tumor target.
There is evidence that NKG2DL protein expression may be associated with aggressiveness (higher TNM stage, higher histological grade, etc.) [64]. In fact, this is what we have seen at the transcription level with TCGA data. However, this also seems to be accompanied by greater dysfunction of NKG2D+ effector cells. This is where the resistance mechanisms described above come into play and form part of the complex process of immune editing that occurs in the tumor to evade the immune system. Among these, the secretion of soluble factors appears to be important. It has been observed that soluble NKG2DL levels are increased in patients with BC and that these levels are associated with aggressiveness [30,78]. In the proposed model, more aggressive tumors would produce more NKG2DL transcripts, but part of the translated proteins would be shed into the microenvironment and bloodstream. This shedding, among other mechanisms, would cause dysfunction of NKG2D+ effector cells (Figure 6).
The data on prognostic value are mixed. With TCGA data, only high RAET1G expression is associated with a worse prognosis. However, the absolute difference between the two subgroups is not clinically significant (0.5 months), with a modest HR (HR 1.4) and no significance in the multivariate analysis. With KMPlot data, high expression of ULBP1, ULBP2, and RAET1E is associated with a worse prognosis. The study by Madjd et al. finds that high expression of MICA/MICB and ULBP-2 is associated with a worse prognosis [63]. However, this is not unique to BC. Similar studies in colorectal or ovarian cancer have identified specific NKG2DLs as prognostic factors [134,135]. One challenge in analyzing NKG2DL expression is that at least eight different ligands are being evaluated, which may lead to concordance issues across datasets. In addition, the distribution of patients into “low” and “high” expression subgroups depends on the cut-off values chosen, influencing subgroup size and survival analyses. In future studies, it may be valuable to develop a composite score integrating expression across multiple NKG2DLs to provide a more robust and reproducible assessment of their prognostic relevance.
Finally, we reviewed potential therapeutic strategies currently being developed to target the NKG2D/NKG2DL axis in BC. On the one hand, it is interesting to note how classic chemotherapy or monoclonal antibodies used in routine clinical practice are capable of overcoming tumor immune-editing mechanisms and promoting NK-mediated cytotoxicity. Looking ahead, clinical studies in humans appear to be focused on adoptive T-cell therapy (especially CAR technology). A search on www.clinicaltrials.gov (accessed on 27 December 2025) using NKG2D as the “intervention/treatment” yields 44 studies. In 41 studies, the target population is cancer patients (12 studies may have included breast cancer patients). Most studies are phase I (with some phase I/II). The experimental treatment is adoptive cell therapy in 39 studies (35 of which are CAR-based) [136].
Potential mechanisms by which BC tumor cells can evade NK cell cytotoxicity mediated by NKG2D/NKG2DL. Illustrations obtained from https://bioart.niaid.nih.gov/ (accessed on 5 February 2026) [137].
5. Future Directions
However, as we have seen, the tumor develops different strategies to evade NKG2D+ cells. Therefore, it seems necessary to develop new approaches to overcome the resistance in the NKG2D/NKG2DL axis [138]. One option would be to combine adoptive cell therapy with other treatment modalities (e.g., concomitant/sequential use of chemotherapy, immune checkpoint inhibitors, ADAM inhibitors, deacetylating agents, etc.). Another option is to optimize adoptive cell therapy with technologies such as CIK, CAR-NK or TRUCK (T cells redirected for antigen-unrestricted cytokine-initiated killing) [139]. In Figure 7, we propose a potential phase Ib/II clinical trial that could be considered after positive preclinical studies have been conducted.
An important consideration is CAR-T cell dosing. Based on the CAR4SAR study in sarcoma, weekly intravenous doses of 3 × 10^6^ cells/kg (up to three doses, with discontinuation in case of ≥grade 3 toxicity) are proposed as a rational reference framework [140].
Anthracyclines and taxanes are two chemotherapy drugs widely used in BC, and there is also evidence that they increase NKG2DL expression [85,86]. However, the classic conditioning regimen for CAR-T therapy uses other chemotherapy modalities (cyclophosphamide and fludarabine) and does not use anthracyclines or taxanes. In fact, the concomitant use of anthracyclines with CAR-T therapy is not recommended [141]. Therefore, the question arises as to how these cytostatic drugs could be incorporated (added to the lymphodepletion regimen or as bridge therapy prior to CAR-T infusion). The potential toxicity associated with the addition of these cytostatic agents must be carefully considered. Importantly, it remains to be determined whether lower, subtherapeutic doses sufficient to induce NKG2DL expression could be employed, potentially limiting treatment-related toxicity.
Another drug that is effective in TNBC is pembrolizumab (anti-PD1) [5]. A study has previously been mentioned in which the administration of anti-PD1 could increase NKG2D+ effector cells [42]. Another interesting target is the methylation of NKG2DL genes. This methylation appears to be described as one of those responsible for immune evasion in acute myeloid leukemia and agents like 5-aza-cytidine could reverse this epigenetic silencing [58]. Old drugs with newly discovered functions are also being studied, such as spironolactone, a diuretic that could increase the expression of NKG2DL in tumor cells [142]. Other drugs of interest currently in preclinical research are ADAM10/17 inhibitors and monoclonal antibodies against sNKG2DL [138].
6. Conclusions
In summary, available in silico data and published evidence indicate that NKG2DL ligands are transcriptionally expressed in BC across cell lines and tumor samples, with higher levels generally observed in biologically aggressive subtypes (especially TNBC). The frequent coexistence of NKG2DL expression with impaired NKG2D-positive effector cell function, together with multiple tumor-driven resistance mechanisms—particularly ligand shedding—highlights the complexity of immune surveillance in BC. However, this raises a starting point for the development of new therapies addressing this NKG2D/NKG2DL axis (with adoptive cell therapy taking on particular importance).
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bray F. Laversanne M. Sung H. Ferlay J. Siegel R.L. Soerjomataram I. Jemal A. Global cancer statistics 2022: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries CA Cancer J. Clin.20247422926310.3322/caac.2183438572751 · doi ↗ · pubmed ↗
- 2Harbeck N. Penault-Llorca F. Cortes J. Gnant M. Houssami N. Poortmans P. Ruddy K. Tsang J. Cardoso F. Breast cancer Nat. Rev. Dis. Primer 201956610.1038/s 41572-019-0111-231548545 · doi ↗ · pubmed ↗
- 3Perou C.M. Sørlie T. Eisen M.B. van de Rijn M. Jeffrey S.S. Rees C.A. Pollack J.R. Ross D.T. Johnsen H. Akslen L.A. Molecular portraits of human breast tumours Nature 200040674775210.1038/3502109310963602 · doi ↗ · pubmed ↗
- 4Prat A. Adamo B. Cheang M.C.U. Anders C.K. Carey L.A. Perou C.M. Molecular characterization of basal-like and non-basal-like triple-negative breast cancer Oncologist 20131812313310.1634/theoncologist.2012-039723404817 PMC 3579595 · doi ↗ · pubmed ↗
- 5Cortes J. Rugo H.S. Cescon D.W. Im S.-A. Yusof M.M. Gallardo C. Lipatov O. Barrios C.H. Perez-Garcia J. Iwata H. Pembrolizumab plus Chemotherapy in Advanced Triple-Negative Breast Cancer N. Engl. J. Med.202238721722610.1056/NEJ Moa 220280935857659 · doi ↗ · pubmed ↗
- 6Bardia A. Hurvitz S.A. Tolaney S.M. Loirat D. Punie K. Oliveira M. Brufsky A. Sardesai S.D. Kalinsky K. Zelnak A.B. Sacituzumab Govitecan in Metastatic Triple-Negative Breast Cancer N. Engl. J. Med.20213841529154110.1056/NEJ Moa 202848533882206 · doi ↗ · pubmed ↗
- 7Rugo H.S. Bardia A. MarméF. Cortés J. Schmid P. Loirat D. Trédan O. Ciruelos E. Dalenc F. Gómez Pardo P. Overall survival with sacituzumab govitecan in hormone receptor-positive and human epidermal growth factor receptor 2-negative metastatic breast cancer (TRO Pi CS-02): A randomised, open-label, multicentre, phase 3 trial Lancet Lond. Engl.20234021423143310.1016/S 0140-6736(23)01245-X 37633306 · doi ↗ · pubmed ↗
- 8Cortés J. Kim S.-B. Chung W.-P. Im S.-A. Park Y.H. Hegg R. Kim M.H. Tseng L.-M. Petry V. Chung C.-F. Trastuzumab Deruxtecan versus Trastuzumab Emtansine for Breast Cancer N. Engl. J. Med.20223861143115410.1056/NEJ Moa 211502235320644 · doi ↗ · pubmed ↗