The Oncogenic Role of Long Non-Coding RNA NEAT1 in Head and Neck Squamous Cell Carcinoma: From Molecular Mechanisms to Clinical Implications
Yuanxin Shi, Bin Chen, Guohui Bai

TL;DR
This paper reviews how the long non-coding RNA NEAT1 contributes to head and neck cancer progression and its potential as a biomarker and therapeutic target.
Contribution
The paper provides a comprehensive review of NEAT1's molecular mechanisms and clinical implications in HNSCC.
Findings
NEAT1 promotes HNSCC progression by acting as a ceRNA for miRNAs like miR-125b-5p and miR-34a-5p.
High NEAT1 expression correlates with metastasis and poor survival in HNSCC patients.
Nanoparticle-based gene silencing offers a potential therapeutic strategy to target NEAT1.
Abstract
Head and neck squamous cell carcinoma (HNSCC) is an aggressive cancer with limited treatment options and poor survival rates. This review focuses on a specific long non-coding RNA NEAT1 (LncRNA NEAT1), which is often found at high levels in HNSCC. We summarize how LncRNA NEAT1 acts as a key driver of cancer progression—promoting tumor growth, spread, and resistance to therapy—by interacting with multiple microRNAs and activating several cancer-related signaling pathways. Clinically, high LncRNA NEAT1 expression is linked to advanced disease stage, metastasis, and worse patient outcomes, indicating its potential as a diagnostic and prognostic biomarker. Although directly targeting LncRNA NEAT1 therapeutically remains challenging, emerging approaches such as nanoparticle-based gene silencing offer promising strategies. Understanding the role of LncRNA NEAT1 may help advance precision…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
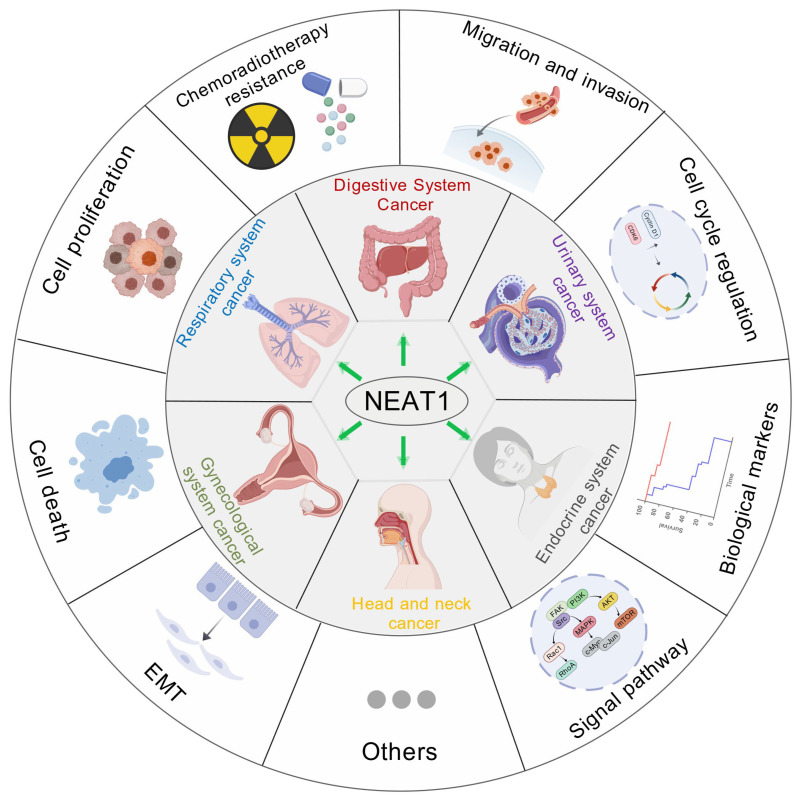 Figure 1
Figure 1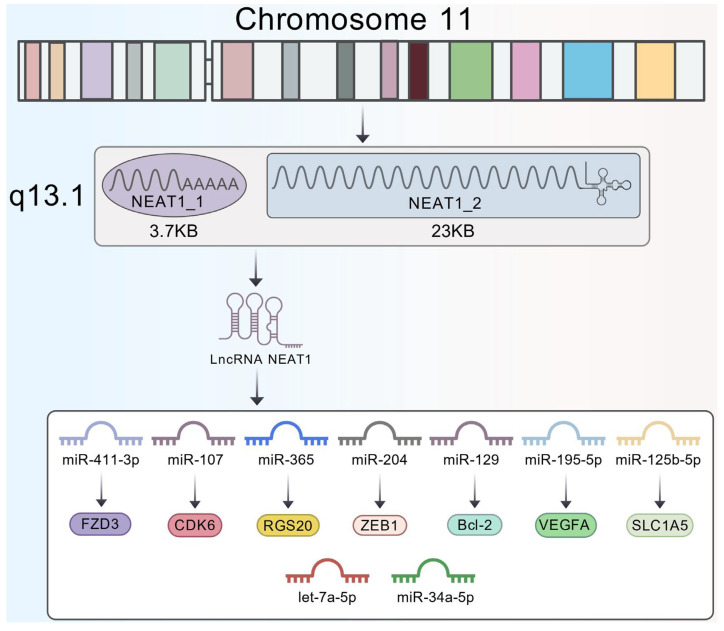 Figure 2
Figure 2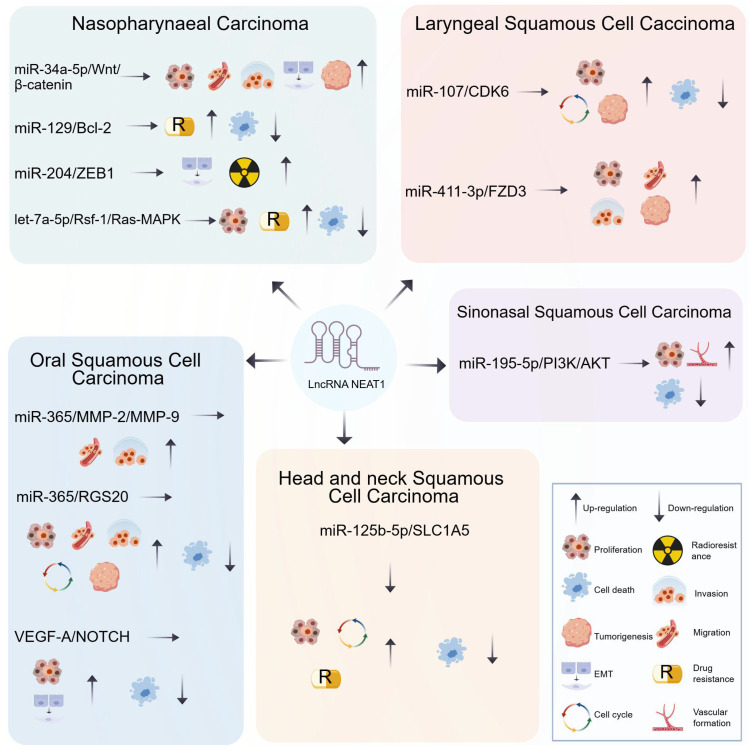 Figure 3
Figure 3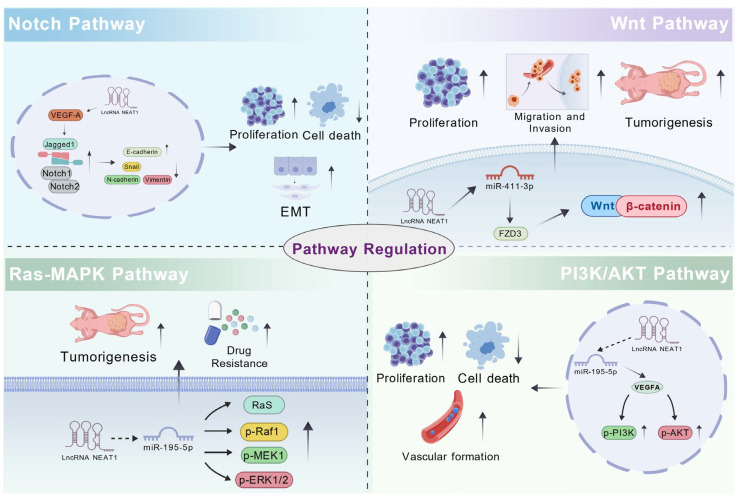 Figure 4
Figure 4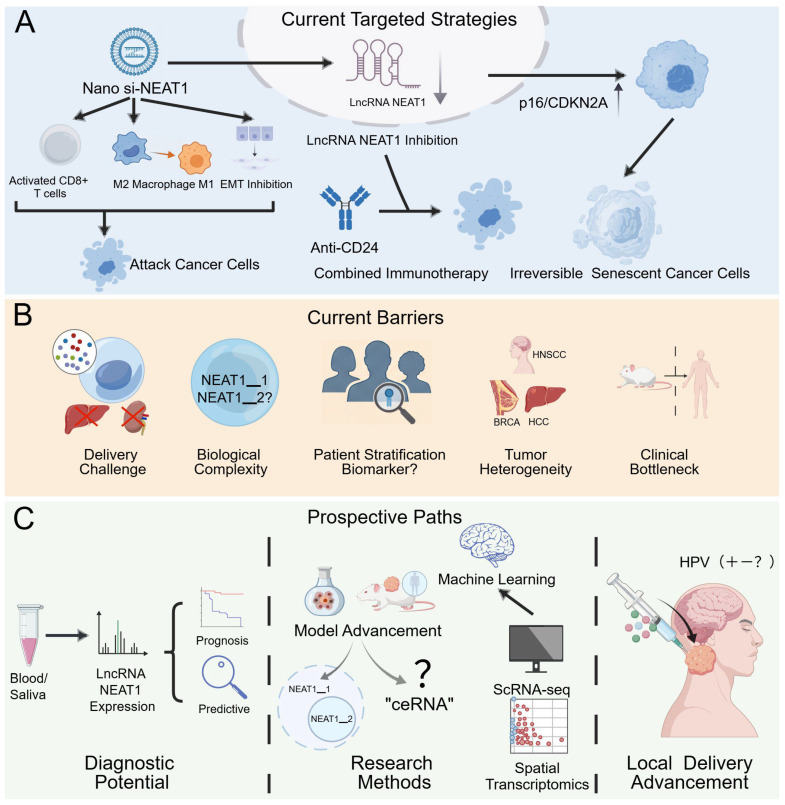 Figure 5
Figure 5- —Guizhou Province Science and Technology Program
- —Key Discipline Construction Project of Higher Education Institutions in Guizhou Province
- —Talent Base Construction Project for the R&D of Medical Biomaterials in Guizhou Province and Zunyi City
- —Guizhou Province Key Laboratory Platform Project for Oral Disease Research of Ordinary Higher Education Institutions
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCancer-related molecular mechanisms research · Cancer-related gene regulation · Autophagy in Disease and Therapy
1. Introduction
Head and Neck Squamous Cell Carcinoma (HNSCC) ranks as the sixth most common cancer globally and remains a clinically challenging disease due to its high rates of local recurrence and distant metastasis. Its incidence continues to rise, with projections indicating a 30% increase by 2030, corresponding to approximately 1.08 million new cases annually [1]. Advanced HNSCC often involves structures of the head and neck region, including the nasal cavity and pharynx, leading to impaired chewing and swallowing functions, facial disfigurement, and speech disorders. These deficits profoundly diminish patients’ quality of life [2]. Moreover, the five-year survival rate for HNSCC remains among the lowest of all solid malignancies and has stagnated at around 50% over the past three decades [3]. Beyond direct mortality, HNSCC survivors exhibit a notably high suicide rate of 63.4 per 100,000—the second highest among all cancer survivors, after pancreatic cancer (86.4 per 100,000), and significantly exceeding that of other cancer survivors (23.6 per 100,000). Psychological distress and reduced quality of life are considered major contributing factors [4]. The high mortality of HNSCC stems from two key challenges: (1) the lack of reliable biomarkers for early diagnosis, and (2) the absence of specific molecular targets to guide precision therapy and improve prognosis [5]. Furthermore, squamous cell carcinomas originating from different anatomical subsites within the head and neck region demonstrate distinct epidemiological characteristics and tumor progression mechanisms, reflecting the heterogeneity and complexity of the disease [6]. This variability further complicates the development of effective treatment strategies, underscoring the need for in-depth investigation into the molecular mechanisms and potential biomarkers of HNSCC.
Non-coding RNAs (ncRNAs) represent a class of RNA molecules that do not encode proteins, such as miRNAs, lncRNAs, circRNAs, and others. They play critical roles in the epigenetic, transcriptional, and post-transcriptional regulation of gene expression, and are involved in a wide range of biological processes including cell differentiation, development, metabolism, and disease pathogenesis [7,8]. In cancer, ncRNAs function as key regulators, acting either as oncogenes to promote malignant phenotypes—such as proliferation, invasion, metastasis, and drug resistance—or as tumor suppressors that inhibit cancer progression. These molecules contribute to tumorigenesis through diverse mechanisms, including miRNA-mediated mRNA degradation, lncRNA-involved epigenetic regulation, circRNA-based ceRNA networks, as well as modulation of the tumor microenvironment and immune responses [9,10]. Moreover, ncRNAs demonstrate considerable clinical potential as diagnostic and prognostic biomarkers, and as therapeutic targets in cancer [11,12].
In head and neck squamous cell carcinoma (HNSCC), lncRNAs can act as oncogenes or tumor suppressors by influencing tumor proliferation, invasion, and metastasis via mechanisms such as epigenetic modification, transcriptional regulation, and competitive binding to miRNAs. As such, they are regarded as potential diagnostic biomarkers and therapeutic targets [13]. Recent studies have identified a number of differentially expressed lncRNAs—including PVT1, LINC00491, MALAT1, HOXA11-AS, HCG22, and LINC01133—that participate in HNSCC progression through various molecular pathways [14,15,16,17,18,19].
LncRNA NEAT1 (Nuclear Enriched Abundant Transcript 1), one of the earliest identified long non-coding RNAs, is predominantly localized in the nucleus where it plays a critical role in the formation and organization of paraspeckles. Studies have demonstrated that LncRNA NEAT1 is commonly upregulated in a variety of solid tumors [20]. lncRNA NEAT1 is significantly up-regulated in a wide range of cancer types, including those of the head and nervous systems (glioma, oral squamous cell carcinoma, laryngeal squamous cell carcinoma, nasopharyngeal carcinoma) [21,22,23,24], digestive system (gastric cancer, colon cancer, esophageal cancer, liver cancer, and pancreatic cancer) [25,26,27,28,29], respiratory system (lung adenocarcinoma, lung squamous cell carcinoma, non-small cell lung cancer) [30,31,32], urinary system (renal cell carcinoma, bladder cancer, prostate cancer) [33,34,35], gynecological system (cervical cancer, ovarian cancer, and endometrial cancer) [36,37,38], endocrine system (thyroid cancer, breast cancer) [39,40], as well as other systems (retinoblastoma, melanoma) [41,42].
In these cancer types, lncRNA NEAT1 promotes tumor development through diverse molecular mechanisms. These include its role as a miRNA sponge, where it sequesters miRNAs and prevents them from mediating the degradation of target genes, thereby regulating multiple signaling pathways—such as VEGF, Notch, and Wnt/β-catenin—to facilitate tumor cell proliferation, migration, and angiogenesis [43,44]. Additionally, lncRNA NEAT1 participates in the regulation of apoptosis and cell cycle progression, influences cancer stem cell properties, and modulates immune cell functions within the tumor microenvironment [45,46]. Notably, under hypoxic conditions, lncRNA NEAT1 expression is induced by HIF-1α, further driving malignant progression [47,48]. Clinically, elevated lncRNA NEAT1 expression is closely associated with poor patient outcomes, supporting its utility as a diagnostic and prognostic biomarker in various cancers [49] (Figure 1).
In summary, the clinical management of head and neck squamous cell carcinoma faces multiple challenges, including tumor heterogeneity, therapy resistance, and a lack of precise biomarkers. A deeper understanding of the role of lncRNAs, particularly lncRNA NEAT1, in tumor biology is crucial for elucidating its oncogenic mechanisms and discovering novel diagnostic and therapeutic targets for HNSCC.
Future research will require the integration of multidisciplinary approaches and cutting-edge technologies. Utilizing single-cell and spatial transcriptomics will enable the precise mapping of lncRNA NEAT1 expression across distinct cellular compartments within the tumor and its microenvironment [50]. Patient-derived organoid models will provide a physiologically relevant platform to dissect lncRNA NEAT1’s functional roles and test targeted interventions in a controlled yet complex setting [51]. Furthermore, applying machine learning algorithms to multi-omics data will be instrumental in deciphering the comprehensive lncRNA NEAT1 regulatory network, predicting its crosstalk with signaling pathways and the tumor immune microenvironment, and ultimately building robust prognostic or predictive models [52]. This concerted effort aims to advance the development of precision therapy and improve patient outcomes.
2. The Structural Characteristics of Long Non-Coding RNA NEAT1
LncRNA NEAT1 (Nuclear Enriched Abundant Transcript 1) is located on chromosome 11q13.1 and gives rise to two major isoforms with distinct structures and functional specializations: the shorter NEAT1_1 (∼3.7 kb) and the longer NEAT1_2 (∼23 kb) [53]. These two isoforms are generated through alternative transcription termination and processing. The shorter transcript, NEAT1_1, is produced via canonical polyadenylation and termination. In contrast, the NEAT1_2 isoform is transcribed when RNA polymerase II reads through the termination signal; its 3ʹ-end, generated by RNase P cleavage, is stabilized by a triple-helix structure rather than a canonical poly(A) tail [54].
Although NEAT1_1 is a linear transcript that does not participate in paraspeckle formation, recent studies have demonstrated that it is not functionally inert [55]. In glioblastoma, the expression ratio of NEAT1_1 to NEAT1_2 is dysregulated, with NEAT1_1 being more prominently upregulated. Deletion of the poly(A) signal of lncRNA NEAT1 using CRISPR–Cas9 significantly reduced NEAT1_1 expression while increasing that of NEAT1_2. This imbalance led to enhanced assembly of nuclear paraspeckles (increased in number), promoted glioma cell migration, and widespread transcriptomic alterations—including upregulation of gene pathways associated with cell migration, adhesion, and polarity. These effects were predominantly driven by the increase in NEAT1_2 rather than the decrease in NEAT1_1, and could be reversed by NEAT1_2-specific knockdown [56]. NEAT1_2 possesses a unique structural feature: inverted repeats at its 3ʹ-end enable the formation of a triple-stranded RNA structure that confers self-stability, allowing it to serve as the core architectural scaffold for the nuclear subdomain known as the paraspeckle. This closed circular topology provides NEAT1_2 with high stability, enabling it to form a regulatory hub within the nucleus—analogous to a membrane-bound organelle—by recruiting various RNA-binding proteins (e.g., SFPQ, NONO) and participating in essential processes such as transcriptional regulation, stress response, RNA editing, and genome stability maintenance [55,57,58,59].
LncRNA NEAT1 plays a pivotal oncogenic role in tumor progression. It promotes cancer cell proliferation, invasion, metastasis, and chemotherapy resistance, largely through dysregulating key signaling pathways such as Wnt/β-catenin and PI3K/AKT [60,61]. In head and neck squamous cell carcinoma (HNSCC), lncRNA NEAT1 promotes tumor progression through multiple regulatory axes—such as miR-411-3p/FZD3 [62], miR-365/RGS20 [22], miR-107/CDK6 [23], and miR-204/ZEB1 [63]—mediating diverse oncogenic mechanisms and highlighting its potential as a therapeutic target in HNSCC (Figure 2). Importantly, it should be noted that most functional studies in HNSCC to date have not distinguished between the two major isoforms (NEAT1_1 and NEAT1_2). Thus, the mechanisms summarized in the following sections primarily reflect the collective functions attributed to NEAT1 in the existing literature.
3. The Mechanism by Which lncRNA NEAT1 Regulates the Progression of HNSCC
3.1. Regulation of Cell Proliferation and Death
The dynamic imbalance between cell proliferation and death serves as a fundamental driver of malignant transformation and cancer progression. Uncontrolled proliferation, mediated by dysregulation of signaling pathways such as Cyclin D-CDK4/6-Rb and PI3K/AKT, enables sustained tumor growth [64,65], while evasion of apoptosis—through mechanisms like BCL-2 overexpression or p53 inactivation—ensures aberrant cell survival [66,67]. This dual dysregulation not only accelerates tumorigenesis and metastasis but also underlies resistance to conventional therapies. Therapeutic approaches targeting the proliferation death axis involve two main strategies: inhibiting cell proliferation using agents such as CDK4/6 inhibitors (e.g., palbociclib) and EGFR inhibitors (e.g., cetuximab) [68,69], and restoring programmed cell death via BCL-2 antagonists (e.g., venetoclax) or ferroptosis inducers (e.g., erastin) [70,71]. These processes are further modulated by the tumor microenvironment (TME), where immune cells such as T lymphocytes can induce apoptosis through the Fas/FasL pathway, and cancer-associated fibroblasts (CAFs) secrete growth factors like HGF and IGF-1 to promote proliferation and suppress apoptosis [72,73]. A comprehensive therapeutic strategy should therefore target both tumor-intrinsic mechanisms and the dynamic crosstalk between cancer cells and their microenvironment. Simultaneously targeting both processes—for instance, combining CDK4/6 inhibitors to restore cell-cycle control with BH3 mimetics to reactivate apoptosis—represents a promising direction in precision oncology [74,75].
Studies have shown that lncRNA NEAT1 is highly expressed across various subtypes of HNSCC, its upregulation is correlated with advanced tumor stage and lymph node metastasis, suggesting its tumor-promoting function [62]. Through the competitive endogenous RNA (ceRNA) mechanism.
lncRNA NEAT1 facilitates cell growth by sequestering multiple microRNAs and dysregulating their downstream targets, These include miR-34a-5p (via the WNT/β-catenin axis) [76], let-7a-5p (via the RASSF1/Ras-MAPK axis), miR-411-3p (via FZD3) [62], miR-107 (via CDK6) [23], miR-365 (via RGS20) [22,77], miR-195-5p (via VEGFA) [78], and miR-125b-5p (via SLC1A5) [79]. Moreover, lncRNA NEAT1 enhances cell survival and chemoresistance by regulating the miR-125b-5p/SLC1A5 axis, which promotes glutamate transport and metabolic reprogramming [79]. Downregulation of lncRNA NEAT1 has been shown to increase oxidative stress, promote apoptosis and autophagy, and thereby suppress tumor proliferation [79]. Regarding the suppression of cell death, lncRNA NEAT1 also inhibits cell death programs by targeting several regulatory axes, such as miR-195-5p (via VEGFA) [78], let-7a-5p (via the RASSF1/Ras-MAPK axis) [24], miR-107 (via CDK6) [23], miR-125b-5p (via SLC1A5) [79], miR-365 (via RGS20) [75], and miR-129 (via Bcl-2) [80]. Notably, its sequestration of miR-195-5p also promotes angiogenesis, thereby providing nutritional support for tumor progression [78]. Furthermore, high lncRNA NEAT1 expression is associated with altered immune cell infiltration patterns in tumor tissues, suggesting its role in modulating immune evasion within the TME, which may indirectly promote HNSCC cell survival and proliferation [79] (Figure 3).
Through its function as a competing endogenous RNA (ceRNA), lncRNA NEAT1 forms a finely-tuned core network that critically regulates the “proliferation-death” balance in HNSCC cells. However, it remains unclear whether hierarchical or synergistic relationships exist among these distinct miRNA axes, and how lncRNA NEAT1 interacts with specific cell types within the tumor microenvironment to indirectly influence this equilibrium. Future investigations should validate these mechanisms in more physiologically relevant models, such as organoid co-culture systems, and explore whether targeting lncRNA NEAT1 can selectively reverse the dysregulation of proliferation and apoptosis in cancer cells while minimizing toxicity to normal tissues [81].
3.2. The Role in Epithelial–Mesenchymal Transition
Epithelial–mesenchymal transition (EMT) plays a pivotal role in the development and progression of head and neck squamous cell carcinoma (HNSCC). EMT is a biological process in which epithelial cells lose their characteristic features and acquire mesenchymal traits, a transition closely associated with enhanced invasiveness and metastatic potential of tumor cells [82,83]. Studies have demonstrated that in HNSCC, EMT not only facilitates tumor cell migration and invasion but also correlates with poor clinical outcomes, including lymph node metastasis and higher pathological grades [84,85].
In-depth analysis of the role of EMT in HNSCC reveals its underlying mechanisms and clinical implications. On one hand, EMT promotes the transition of tumor cells to a mesenchymal phenotype by regulating multiple signaling pathways, such as the transforming growth factor-β (TGF-β) pathway, thereby enhancing their migratory and invasive capabilities [1,86]. Moreover, the morphological and molecular changes during EMT may impair immune recognition and attack of tumor cells, further facilitating immune escape [87]. On the other hand, key factors such as TWIST1 and IGF2BP1 have been found to regulate the EMT process, and their expression levels are closely associated with patient prognosis, providing new targets for individualized therapy [88,89].
Among them, lncRNA NEAT1 plays a key role in HNSCC: it promotes EMT by modulating the VEGF-A/Notch axis, characterized by down-regulation of E-cadherin and up-regulation of N-cadherin, vimentin, and Snail [90]. Additionally, lncRNA NEAT1 can sequester microRNAs such as miR-34a-5p (via WNT/β-catenin) and miR-204 (via ZEB1), thereby relieving their repression on target genes or directly enhancing the EMT program [63,76], ultimately influencing tumor cell metastatic and invasive processes (Figure 3).
Accumulating evidence supports the consensus that lncRNA NEAT1 acts as a pleiotropic regulator driving the EMT program in HNSCC. It functions by sequestering multiple EMT-suppressive miRNAs, thereby relieving their repression on key EMT-related transcription factors including ZEB1 and Snail. Nevertheless, several important questions remain. For instance, are these distinct lncRNA NEAT1/miRNA axes universally activated across all HNSCC subtypes, or are they selectively engaged under specific genetic backgrounds or microenvironmental cues? Furthermore, whether lncRNA NEAT1-mediated EMT is associated with full reversibility (i.e., mesenchymal-to-epithelial transition) and what role it plays in maintaining cancer stem-like properties require further clarification.
3.3. Regulation of Invasion and Migration
During the progression of head and neck squamous cell carcinoma (HNSCC), the migratory and invasive capabilities of tumor cells represent critical determinants of malignancy. Accumulating evidence indicates that tumor cells employ diverse mechanisms to enhance their mobility and invasiveness, ultimately leading to more severe clinical outcomes.
Long non-coding RNAs (lncRNAs) have emerged as important regulatory factors in modulating tumor cell migration and invasion. These molecules influence gene expression through multiple mechanisms, such as functioning as miRNA sponges or directly regulating target genes, thereby actively participating in HNSCC pathogenesis [91]. Furthermore, lncRNAs may enhance the migratory capacity of tumor cells by modulating the epithelial–mesenchymal transition (EMT) process, which is critical for the invasive behavior of HNSCC [92,93]. Current studies indicate that lncRNA NEAT1 is upregulated in metastatic HNSCC cells compared to primary cancer cells [94]. Furthermore, multiple reports have detected higher expression levels of lncRNA NEAT1 in tissues from cervical lymph node metastases and advanced-stage HNSCC, a finding further supported by meta-analyses [23,76,77].
Across various subtypes of head and neck squamous cell carcinoma (HNSCC), lncRNA NEAT1 enhances cellular migration and/or invasion through multiple miRNA/target gene signaling axes. These include the miR-34a-5p/WNT/β-catenin axis [76], the miR-125b-5p/SLC1A5 axis [79], the miR-411-3p/FZD3 axis [62], the miR-365/RGS20 axis [22], and the miR-365/MMP-2/MMP-9 axis [77]. These mechanisms collectively underscore the multi-pathway synergy of lncRNA NEAT1 in regulating the invasive and metastatic processes of HNSCC (Figure 3).
In summary, lncRNA NEAT1 drives the migration and invasion of HNSCC cells through an overlapping and redundant ceRNA network involving miRNAs. This constitutes a robust and well-supported consensus mechanism. The functional convergence of these axes may reflect the network robustness evolved by tumor cells to sustain metastatic competence. However, how these mechanisms are precisely sequenced or coordinated during the spatiotemporal dynamics of invasion and metastasis—such as from local invasion to intravasation, circulatory survival, extravasation, and colonization—remains to be elucidated. Furthermore, whether lncRNA NEAT1 influences migration by more directly regulating physical processes, such as cytoskeletal dynamics or cell–matrix adhesion, awaits further experimental investigation.
3.4. Regulation of Signaling Pathways
The regulation of signaling pathways is critically important in tumor initiation and progression. Dysregulation of several classic signaling cascades—such as PI3K/AKT, RAS/MAPK, Wnt/β-catenin, and Notch—constitutes a central mechanism driving malignant transformation in cancer cells. Abnormal activation or inhibition of these pathways collectively promotes tumor progression and metastasis by modulating key cellular processes, including proliferation, apoptosis, metabolism, invasion, migration, angiogenesis, and tumor microenvironment remodeling [95,96,97,98,99]. Therefore, elucidating the regulatory networks and interactions of specific signaling pathways in tumors not only aids in uncovering the mechanisms of tumor pathogenesis but also facilitates the development of novel molecularly targeted therapeutic strategies.
Specifically, lncRNA NEAT1 contributes to HNSCC progression by orchestrating the activation of multiple oncogenic signaling pathways. It activates the Wnt/β-catenin pathway through the miR-411-3p/FZD3 axis [62]. In parallel, it promotes PI3K/AKT signaling via the miR-195-5p/VEGFA axis, indicated by increased p-PI3K and p-AKT levels [78]. Additionally, lncRNA NEAT1 upregulates VEGF-A to stimulate the Notch pathway [90], while also activating the Ras-MAPK cascade through the let-7a-5p/Rsf-1 axis, evidenced by elevated expression of Ras, p-Raf1, p-MEK1, and p-ERK1/2 [24]. Together, these coordinated mechanisms mediated by lncRNA NEAT1 drive the malignant phenotype and advance tumor progression in HNSCC (Figure 4).
Collectively, lncRNA NEAT1 functions not as a regulator of a single pathway but as a signaling “cross-hub.” Through its ceRNA activity, it concurrently modulates multiple core oncogenic pathways. This multi-pathway regulatory capacity likely underlies the pleiotropic pro-tumorigenic functions of lncRNA NEAT1. Nevertheless, several important questions remain: whether substantial cross-talk exists among these lncRNA NEAT1-activated pathways; whether lncRNA NEAT1 itself is subject to feedback regulation by downstream effectors of these pathways; and, in different HNSCC subtypes or microenvironmental contexts, which pathway serves as the predominant effector of lncRNA NEAT1-driven oncogenesis.
3.5. Diagnostic and Prognostic Biomarkers
Despite accumulating evidence suggesting the potential of long non-coding RNA NEAT1 (lncRNA NEAT1) as a diagnostic and prognostic biomarker in various common malignancies, a systematic summary of its role in head and neck squamous cell carcinoma (HNSCC) remains lacking. For instance, Meta-analyses have indicated that elevated lncRNA NEAT1 expression is associated with poor prognosis in patients with multiple cancer types and may serve as a predictor of clinicopathological features [100]. Furthermore, Shieh et al. demonstrated that high lncRNA NEAT1 expression correlates with unfavorable survival outcomes in HNSCC patients, indicating poorer prognosis even after radiotherapy and chemotherapy [101]. Similarly, Yuan et al., using Mendelian randomization and eQTL analysis, revealed that the lncRNA NEAT1 rs3741384 GG genotype predicts adverse prognosis in HNSCC patients, suggesting that genetic variations in lncRNA NEAT1 may influence patient survival [102].
In terms of diagnostic value, lncRNA NEAT1 has also been recognized as a diagnostic biomarker in several human cancers [103]. In HNSCC, tumor tissues exhibit significantly higher levels of lncRNA NEAT1 compared to adjacent non-tumor tissues (3.041 ± 0.709-fold, p < 0.01). LncRNA NEAT1 expression is significantly correlated with tumor T stage, cervical lymph node metastasis, and clinical stage in HNSCC, with higher expression observed in T3–T4 tumors, lymph node metastases, distant metastases, and advanced clinical stages [23,104,105].
Notably, multiple studies in HNSCC have consistently reported that upregulated lncRNA NEAT1 expression is strongly associated with adverse clinical outcomes (Table 1), further supporting its reliability as a prognostic indicator and potential therapeutic target. Moreover, the diagnostic utility of lncRNA NEAT1 in HNSCC has been confirmed by several investigations (Table 1), highlighting its considerable potential as a clinical diagnostic biomarker.
A clear consensus has emerged from current research: lncRNA NEAT1 is highly expressed in HNSCC tissues, and its expression level shows a significant positive correlation with advanced tumor stage, lymph node metastasis, and poor prognosis. These findings strongly support its potential as a promising diagnostic and prognostic biomarker. However, most evidence is derived from tissue samples; therefore, its stability and diagnostic performance in liquid biopsy sources such as plasma or saliva require validation in prospective, large-scale cohorts [107]. Another major challenge lies in establishing a universally applicable and standardized detection threshold (cut-off value). Finally, integrating lncRNA NEAT1 with other established markers (e.g., HPV status, PD-L1) or emerging non-coding RNAs into a multi-molecular signature may offer greater predictive value than a single biomarker alone, representing a critical direction for future research [108].
3.6. Resistance to Therapeutic Agents and Strategies Targeting lncRNA NEAT1
Chemoresistance represents a major cause of treatment failure and poor prognosis in patients with head and neck squamous cell carcinoma (HNSCC) [109]. Cisplatin-based chemotherapy regimens have become a cornerstone of systemic treatment for HNSCC [110]. However, the intrinsic or acquired resistance of tumor cells significantly limits its clinical efficacy. Therefore, elucidating the underlying molecular mechanisms of chemoresistance in HNSCC and identifying potential targets to reverse drug resistance hold considerable scientific and clinical value for improving patient outcomes [111].
In recent years, the role of non-coding RNAs (ncRNAs) in tumorigenesis, progression, and therapy resistance has garnered increasing attention. Through involvement in complex regulatory networks, ncRNAs influence multiple biological processes—such as cell proliferation, apoptosis, epithelial–mesenchymal transition (EMT), and tumor microenvironment remodeling—thereby modulating drug resistance in cancers [7,112,113]. Multiple studies have revealed that lncRNA NEAT1 is significantly upregulated in HNSCC and closely associated with adverse pathological features and poor prognosis, suggesting its role as a key molecule promoting malignant progression. Notably, lncRNA NEAT1 plays an important role in regulating therapeutic sensitivity in HNSCC. For example, high expression of lncRNA NEAT1 significantly reduces the survival rate of HNSCC patients, even after radiotherapy or chemotherapy [101]. Furthermore, existing studies indicate that lncRNA NEAT1 promotes migration, invasion, and cisplatin resistance in HNSCC cells via the miR-125b-5p/SLC1A5 axis [79]; influences cisplatin resistance and tumorigenicity in nude mice through regulation of let-7a-5p [24]; and modulates EMT and radioresistance in HNSCC via the miR-204/ZEB1 axis [63] (Figure 5A).
These findings suggest that lncRNA NEAT1 may exert pleiotropic regulatory functions across different drug treatment contexts. LncRNA NEAT1 shows promise not only as a biomarker for predicting therapeutic response but also as a potential target for enhancing the efficacy of existing regimens and developing novel intervention strategies.
Targeting lncRNA NEAT1 has emerged as a promising novel direction in cancer therapeutics. LncRNA NEAT1 is frequently overexpressed in multiple cancer types and promotes tumor progression through diverse mechanisms, including participation in DNA repair, modulation of the immune microenvironment, and regulation of cellular senescence. Current targeting strategies primarily involve the following four approaches: One key strategy utilizes nanoparticle-based delivery of siRNA or shRNA to specifically silence lncRNA NEAT1. For example, Chen et al. employed layered double hydroxide (LDH) nanoparticles to deliver si-NEAT1 (LDH@si-NEAT1), which activated CD3+CD8+ T cells, inhibited immune evasion in breast cancer cells, polarized M2-type TAMs towards an M1 phenotype, suppressed EMT, and remodeled the immunosuppressive microenvironment in mouse models, thereby synergizing with anti-PD-1 therapy to enhance antitumor immunity [114]. Additionally, combining lncRNA NEAT1 inhibition with existing immune checkpoint inhibitors—such as anti-CD24 antibodies—has been shown to significantly suppress tumor growth and counteract immune escape [115]. Another intriguing study in liver cancer demonstrated that inhibiting lncRNA NEAT1 relieves its repression of senescence-associated pathways (e.g., CDKN2A/p16), driving cancer cells into irreversible senescence. This finding provides a novel rationale for exploiting “therapy-induced senescence” as a potential treatment strategy [116] (Figure 5A).
However, it is important to note that all the findings summarized above remain at the preclinical stage, and most are not based on HNSCC-specific models. LncRNA NEAT1-targeted therapy has not yet entered clinical trials for any cancer, and its translation faces multiple practical hurdles: (1) Delivery and specificity challenges: lncRNA NEAT1 is predominantly localized in the nucleus, necessitating efficient, specific, and safe delivery of siRNA/ASOs into the nuclear compartment—a key technical hurdle for RNA interference. Current nanodelivery systems still require optimization in terms of targeting efficiency, organ selectivity, and immunogenicity. Off-target effects could lead to unpredictable toxicity in normal tissues [117]. (2) Biological complexity of the target: lncRNA NEAT1, particularly NEAT1_2, serves as the functional core of nuclear paraspeckles, which are involved in normal stress responses, RNA processing, and the maintenance of genome stability. Global inhibition of lncRNA NEAT1 may disrupt these fundamental physiological processes, posing long-term safety risks [118]. (3) Unclear patient stratification and beneficiary population: HNSCC exhibits high heterogeneity, and it remains unclear which subtypes or molecular subgroups would benefit most from lncRNA NEAT1 inhibition. The lack of reliable predictive biomarkers significantly increases the risk of clinical trial failure [119]. (4) Tumor heterogeneity: The function and regulatory mechanisms of lncRNA NEAT1 may vary across cancer types and even among different subtypes of the same cancer, necessitating further clarification of its context-dependent roles [120]. (5) Clinical translation bottleneck: All significant progress remains confined to preclinical studies, and rigorous human trials are required to validate both safety and efficacy (Figure 5B) [121,122].
Given these fundamental challenges, advancing lncRNA NEAT1 as a predictive biomarker may currently represent a more translationally feasible path than its direct development as a therapeutic target. Its high expression is closely associated with poor prognosis and treatment resistance, supporting its role as a prognostic indicator [101,106]. Further prospective studies could validate whether lncRNA NEAT1 expression levels—detected via liquid biopsies such as blood or saliva—can predict patient responses to specific radiotherapy or chemotherapy regimens, thereby guiding pre-treatment risk stratification and personalized therapeutic strategies. This approach entails significantly lower technical and regulatory barriers compared with developing novel RNA-targeting drugs [123] (Figure 5C).
In summary, although the role of lncRNA NEAT1 in mediating therapy resistance in HNSCC is increasingly clear, its direct targeting as a therapeutic agent remains a formidable challenge. Future research should proceed along two parallel tracks: first, rigorously evaluating the efficacy and safety of lncRNA NEAT1 inhibition in more clinically relevant HNSCC models (e.g., patient-derived organoids) and exploring strategies such as localized delivery to reduce systemic toxicity; second, prioritizing the multicenter validation of lncRNA NEAT1 as a clinical biomarker—a pathway that may currently offer a faster and more practical route toward clinical translation.
4. Conclusions
This review systematically elucidates the pleiotropic oncogenic functions of the long non-coding RNA NEAT1 (LncRNA NEAT1) in head and neck squamous cell carcinoma (HNSCC), establishing its central role in regulating critical processes such as tumor proliferation, metastasis, and therapy resistance via a complex ceRNA network (Figure 3). However, significant limitations and unresolved questions persist in the current body of research, delineating key directions for future investigation.
First, the function of lncRNA NEAT1 exhibits substantial context-dependency. HNSCC itself is highly heterogeneous in terms of anatomical origin, etiology (e.g., HPV infection status), and tumor microenvironment [124]. Although lncRNA NEAT1 displays pro-tumorigenic properties across multiple subtypes, its upstream regulatory signals, predominant downstream pathways, and clinical performance as a biomarker likely vary. For instance, A key regulatory axis involves the tumor suppressor p53, which directly binds to the NEAT1 promoter and activates its transcription. This p53-mediated induction of NEAT1 contributes to the tumor-suppressive function of p53, forming a reciprocal transcriptional regulatory network [125]. Consequently, any upstream event affecting p53 stability or function may perturb NEAT1 expression. In HPV-positive HNSCC, the viral oncoprotein E6 binds to the host ubiquitin ligase E6AP, forming a complex that specifically targets p53 for ubiquitin-proteasome-mediated degradation. This constitutes a core mechanism of HPV-driven carcinogenesis, leading directly to the loss of p53 tumor-suppressor activity [126]. From this, the following regulatory cascade can be deduced: HPV positivity → E6 overexpression → p53 degradation → impaired transcriptional activity of p53 → potential suppression or dysregulation of its downstream target gene NEAT1. Thus, in HPV-positive cases, E6-mediated p53 degradation likely reduces p53 transcriptional activity, thereby weakening its induction of NEAT1 and potentially leading to suppressed or aberrant NEAT1 expression. In contrast, in HPV-negative HNSCC—particularly those associated with smoking—TP53 frequently acquires inactivating or gain-of-function mutations due to tobacco exposure [127]. These mutations result in loss of normal p53 transcriptional activity or acquisition of oncogenic functions, which can also drive NEAT1 dysregulation. The patterns of NEAT1 dysregulation and the associated biological contexts (e.g., genomic instability, oxidative stress) thus differ fundamentally between HPV-positive and HPV-negative tumors. Notably, the biological role of NEAT1 in tumors is dynamically determined by p53 status: it exerts a tumor-suppressive role in synergy with functional p53, whereas upon p53 inactivation, NEAT1 may shift toward promoting tumorigenesis [128]. This context-dependent mechanism holds significant implications for both HPV-positive and HPV-negative head and neck squamous cell carcinomas.
Beyond its context-dependent regulation by p53, NEAT1 functions across various solid tumors as a pivotal molecular scaffold, orchestrating oncogenic gene networks by recruiting specific transcriptional or chromatin-modifying complexes. It operates through distinct modes: as a transcriptional co-regulatory platform—for example, in prostate cancer, where it directly binds transcription factor CDC5L to co-activate the oncogene AGRN [129]—and as an epigenetic regulatory hub, as seen in gastric cancer, where it mediates ALKBH5/EZH2-driven silencing of tumor suppressors [130]. Its own expression is also tightly controlled, such as by MUC1-C via PBAF-dependent chromatin remodeling [131]. This evidence underscores NEAT1’s role as a dynamic signal-integrating node, suggesting that in HNSCC, it may similarly recruit HPV-status-specific protein complexes to drive divergent downstream pathways, thereby contributing to the distinct phenotypes observed in HPV-positive and HPV-negative tumors. Currently, the specific mechanisms by which NEAT1 recruits transcriptional or chromatin-modifying complexes in HNSCC remain insufficiently explored. Therefore, future research should systematically map the NEAT1-interacting proteome in HNSCC to decode its precise, context-dependent regulatory circuits. It is also imperative to validate the association between NEAT1 expression and prognosis or therapeutic response in well-stratified clinical cohorts and corresponding experimental models (e.g., patient-derived organoids), based strictly on anatomical subsites and HPV status [119,132].
Second, the functional complexity of lncRNA NEAT1 extends far beyond our current understanding. On one hand, its two major transcript isoforms, NEAT1_1 and NEAT1_2, likely have distinct roles based on their subcellular localization, molecular interactions, and functions [55]. The long isoform NEAT1_2 is essential for the scaffold and formation of paraspeckles, a type of nuclear body involved in various regulatory processes, while the short isoform NEAT1_1 may exert functions independent of this structure [131]. Critically, current research in HNSCC largely overlooks the specific contributions of paraspeckles themselves to cancer progression. Whether the oncogenic functions attributed to lncRNA NEAT1 are predominantly mediated through paraspeckle-dependent mechanisms (e.g., sequestering proteins or RNAs within the nuclear body) or through paraspeckle-independent activities of individual isoforms remains poorly defined [131]. This gap underscores a significant limitation in the field. Most existing studies treat lncRNA NEAT1 as a homogeneous entity, obscuring which specific biological effects are mediated by which isoform. Clarifying the individual contributions and dynamic balance between NEAT1_1 and NEAT1_2 in HNSCC is pivotal for a deeper understanding of its molecular mechanisms. Therefore, future studies should prioritize experimental approaches that can distinguish between the two isoforms—such as isoform-specific knockdown or overexpression—to delineate their individual functions and potential synergy. Concurrently, methodologies that specifically disrupt paraspeckle architecture without abolishing lncRNA NEAT1 expression are needed to clarify the role of this nuclear sub-structure. Together, these lines of investigation will be essential for elucidating the precise molecular mechanisms of lncRNA NEAT1 in HNSCC and for informing the development of targeted therapeutic strategies, whether they are isoform-specific or pathway-specific.
On the other hand, the role of lncRNA NEAT1 in shaping the HNSCC immune microenvironment remains largely unexplored. Although studies in other cancers suggest lncRNA NEAT1 can induce CD8^+^ T cell exhaustion via axes such as miR-155/Tim-3 [133], it is still unknown whether and how lncRNA NEAT1 differentially regulates immune cell infiltration and checkpoint expression in the immunologically distinct contexts of HPV-positive versus HPV-negative HNSCC. Systematic investigation using models like co-culture organoids or humanized mice is warranted [134] (Figure 5C).
Similarly, numerous other lncRNAs, such as lncRNA PVT1 and lncRNA MALAT1 [14], have also been widely demonstrated to be involved in HNSCC progression. However, there are both similarities and differences in their roles. For instance, in terms of clinical significance, both lncRNA NEAT1 and lncRNA MALAT1 show potential as liquid biopsy biomarkers for early diagnosis, prognosis assessment, and treatment response prediction in HNSCC. Nevertheless, differences exist in their clinical correlations. For example, in predicting radiotherapy-related toxicities, low expression of both lncRNA NEAT1 and lncRNA MALAT1 is significantly associated with risks of anemia, hepatic dysfunction, and malnutrition. Yet their sensitivity and specificity in identifying specific toxicity types are not identical, suggesting that they may participate in the development of treatment-related adverse reactions through distinct molecular mechanisms [135]. This comparative perspective underscores the need to precisely define the unique and shared pathways governed by different oncogenic lncRNAs, including lncRNA NEAT1, within the HNSCC landscape.
Furthermore, the predominant interpretation of lncRNA NEAT1′s core mechanism—the ceRNA model—requires critical scrutiny. As a predominantly nuclear-localized lncRNA (especially NEAT1_2), the spatial logic of lncRNA NEAT1 efficiently acting as a cytoplasmic “molecular sponge” for miRNAs under physiological conditions presents a challenge [136]. Current evidence largely relies on correlative analyses and functional rescue experiments, lacking direct, quantitative validation of the stoichiometry of its interactions with miRNAs. Future studies should employ more rigorous techniques, such as crosslinking immunoprecipitation sequencing (CLIP-seq) and single-molecule imaging, to validate or refine this prevailing model [136]. We should also explore the possibility of lncRNA NEAT1 functioning through non-ceRNA mechanisms, such as transcriptional regulation or sequestration of nuclear factors (Figure 5C).
Finally, translating lncRNA NEAT1 research from the bench to the clinic necessitates the integration of novel methodologies. Single-cell and spatial transcriptomics can precisely resolve lncRNA NEAT1 expression patterns across different intratumoral cell subpopulations (e.g., cancer cells, immune cells, fibroblasts) and their impact on cellular states [137,138]. Multi-omics integrative analyses can systematically uncover the upstream genetic and epigenetic drivers of aberrant lncRNA NEAT1 expression and reconstruct its global regulatory network [139,140]. Engineered models based on patient-derived organoids provide an ideal platform for manipulating lncRNA NEAT1 expression within a three-dimensional, near-physiological context to observe its effects on tumor growth, immune interactions, and therapeutic response [141]. Furthermore, machine learning algorithms can mine high-throughput data to build prognostic or predictive models based on lncRNA NEAT1 and its associated features, thereby identifying patient subgroups most likely to benefit from targeted intervention [142,143] (Figure 5C).
In conclusion, future research on lncRNA NEAT1 should move beyond the traditional view of it as a single, homogeneous molecule and instead focus on its heterogeneity and multidimensional complexity. By leveraging cutting-edge technologies to dissect its functional mechanisms within finely stratified contexts, there is significant potential to transform lncRNA NEAT1 from a correlative marker into a diagnostic tool and precision therapeutic target with clear clinical utility.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Chen X. Zhang S. Liu C. Li G. Lu S. Wang Y. Zhang X. Huang D. Qiu Y. Liu Y. UBE 2O Promotes Progression and Epithelial-Mesenchymal Transition in Head and Neck Squamous Cell Carcinoma Onco Targets Ther.2020136191620210.2147/OTT.S 253861 PMC 733401432636643 · doi ↗ · pubmed ↗
- 2Warnakulasuriya S. Global epidemiology of oral and oropharyngeal cancer Oral Oncol.20094530931610.1016/j.oraloncology.2008.06.00218804401 · doi ↗ · pubmed ↗
- 3Ren Z.H. Hu C.Y. He H.R. Li Y.J. Lyu J. Global and regional burdens of oral cancer from 1990 to 2017: Results from the global burden of disease study Cancer Commun.202040819210.1002/cac 2.1200932067418 PMC 7163731 · doi ↗ · pubmed ↗
- 4Osazuwa-Peters N. Simpson M.C. Zhao L. Boakye E.A. Olomukoro S.I. Deshields T. Loux T.M. Varvares M.A. Schootman M. Suicide risk among cancer survivors: Head and neck versus other cancers Cancer 20181244072407910.1002/cncr.3167530335190 · doi ↗ · pubmed ↗
- 5Yu W. He X. Zhang C. Huangfu H. Transcriptomics data mining to uncover signature genes in head and neck squamous cell carcinoma: A bioinformatics analysis and RNA-sequencing based validation Am. J. Cancer Res.2023135513553038058845 PMC 10695818 · pubmed ↗
- 6Ferreira C.C. The relation between human papillomavirus (HPV) and oropharyngeal cancer: A review Peer J 202311 e 1556810.7717/peerj.1556837397013 PMC 10309048 · doi ↗ · pubmed ↗
- 7Nemeth K. Bayraktar R. Ferracin M. Calin G.A. Non-coding RN As in disease: From mechanisms to therapeutics Nat. Rev. Genet.20242521123210.1038/s 41576-023-00662-137968332 · doi ↗ · pubmed ↗
- 8Liu X. Haugh W. Zhang Z. Huang J. Emerging Role of Long, Non-Coding RNA Nuclear-Enriched Abundant Transcript 1 in Stress- and Immune-Related Diseases Int. J. Mol. Sci.202526441310.3390/ijms 2609441340362651 PMC 12072541 · doi ↗ · pubmed ↗