Sex and Age Differences in Decision-Making Under Risk by Wild Balinese Long-Tailed Macaques (Macaca fascicularis fascicularis): A Field Experimental Study
Caleb Bunselmeyer, Noëlle Gunst, I Nengah Wandia, Robert J. Williams, Elsa Addessi, Jean-Baptiste Leca

TL;DR
Wild macaques show varied risk preferences influenced by age and sex, with juveniles adapting choices based on past outcomes.
Contribution
The study reveals how age and sex interact to shape risk preferences in wild macaques, highlighting individual variability.
Findings
Macaques showed no group-level preference for safe or risky choices but had strong individual differences.
Male juveniles and older adults were more risk-prone than younger males, while adult females were more risk-prone than juvenile females.
Juveniles adapted their choices based on previous outcomes, following a win–stay strategy.
Abstract
Most past research on risk preferences in primates focused on captive animals and neglected individual differences. We studied how 33 wild Balinese long-tailed macaques of different ages and sexes make risky choices. The animals were given a choice between a small reward they would always get and a larger reward they would get only sometimes. Overall, the macaques did not prefer either the safe or the risky option. However, individuals differed greatly. Some macaques often chose the risky option, others preferred the safe one, and many showed no clear preference. These differences were strongly linked to age, sex, and what happened in previous trials. Age and sex worked together in shaping risk preferences. Among males, juveniles and older adults were more risk-prone than younger adults. Among females, adults were more risk-prone than juveniles. Juveniles were also flexible in their…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
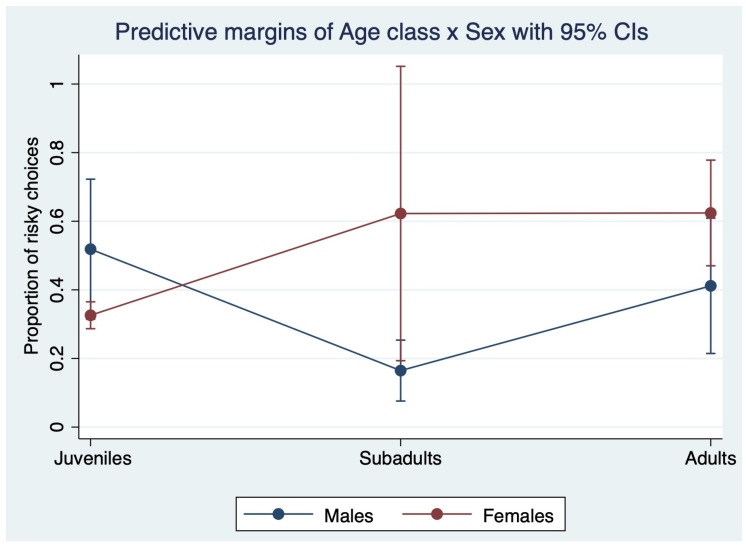 Figure 1
Figure 1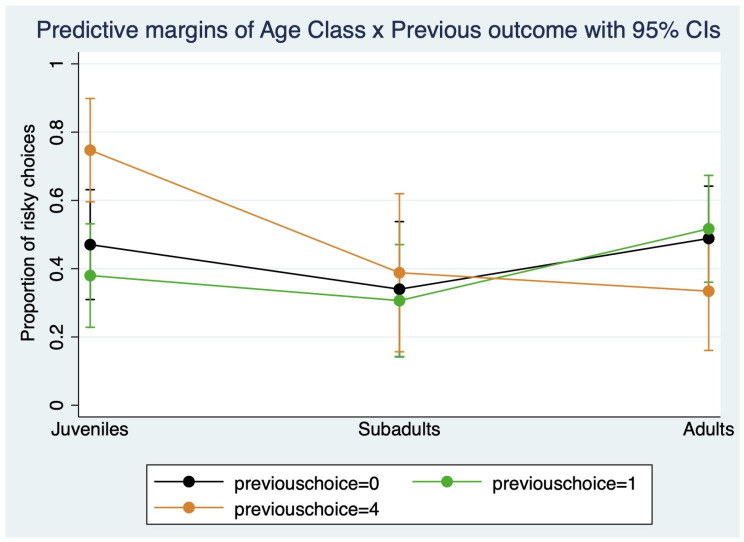 Figure 2
Figure 2- —Major Grant from the Alberta Gambling Research Institute
- —Natural Sciences and Engineering Research Council of Canada
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPrimate Behavior and Ecology · Hemispheric Asymmetry in Neuroscience · Memory and Neural Mechanisms
1. Introduction
Risk, defined as variability in the rate of gains [1], pervades the daily life of wild animals. Due to the unpredictable nature of their environments, animals face risks on a daily basis across diverse contexts such as foraging, seeking mating opportunities, and avoiding predators [2]. In decisions involving risk, individuals typically choose between a safe option (offering a small but certain reward) and a risky alternative (offering a potentially larger yet uncertain reward), with known probabilities assigned to each outcome [3,4]. Numerous comparative studies have examined risk preferences across various primate species with different feeding ecologies. Species usually exploiting more variable food sources, including rhesus macaques (Macaca mulatta), capuchin monkeys (Sapajus spp.), chimpanzees (Pan troglodytes), and orangutans (Pongo abelii), exhibited a risk-prone attitude when faced with probabilistic payoff variations. Conversely, species reliant on more stable resources or inhabiting harsh environments, such as bonobos (Pan paniscus), gorillas (Gorilla gorilla), and lemurs (Lemur catta, Eulemur mongoz, Varecia rubra) displayed risk-averse tendencies [5,6]. Nonetheless, more recent research revealed that this overarching pattern is not always consistent. Rhesus macaques, capuchin monkeys, and chimpanzees exhibit varying risk preferences—from risk-seeking to risk-averse—depending on the task and contextual factors [4,7,8,9,10,11,12]. This variability suggests that observed inter-specific differences in risk preferences might instead reflect interindividual variation within species.
Sex and age represent two relevant demographic factors that may shape risk preferences, though experimental evidence is still limited. Regarding sex differences, several studies report that male chimpanzees and capuchin monkeys exhibit greater risk tolerance compared to females. In the wild, male chimpanzees were more frequently observed hunting colobus monkeys (Procolobus spp.), a potentially hazardous behavior that only sometimes results in success [5,13]. Similarly, in a population of sanctuary-living chimpanzees tested in a risky-choice task, males demonstrated greater risk preferences than females [10]. This pattern is echoed in captive male capuchin monkeys tested in an analogous task [14]. Nevertheless, other studies have not found significant sex differences in risk preference within these species [3,4,15,16], with failures not evidently attributable to sample size limitations [5].
Regarding age differences, in sanctuary-living chimpanzees tested in a risky-choice task, adolescent and young adults exhibited greater risk proneness compared to older adults [10,17]. This developmental pattern, characterized by higher risk proneness during adolescence followed by a more conservative approach during adulthood, closely mirrors the well-documented trend observed in humans [18]. This parallel suggests that heightened risk proneness during adolescence may be linked to a higher necessity of exploring the environment, competing for social status, and/or dispersing from the natal group. However, a study on probability inference in rhesus macaques [19] reported comparable risk preferences between younger and older individuals. A more nuanced pattern emerged from a recent study on capuchin monkeys [12], revealing a three-way interaction among task contingencies, age, and past experience. Specifically, older monkeys demonstrated a higher propensity to take risks compared to younger ones when learning outcome probabilities through experience, with a lower age-related risk proneness in individuals with greater prior experience in risky-choice tasks.
Risk preferences have been extensively investigated in captive non-human primates through paradigms involving learned associations, intuitive outcomes and exchange tasks, respectively reviewed in [5]. In the first paradigm, subjects first underwent a training phase during which they learned the expected values of a certain option and a probabilistic alternative, followed by repeated choices between these options. The certain option yielded a constant payoff, whereas the risky option provided a reward that varied probabilistically in quantity or quality [3,4]. In the second paradigm, subjects could choose between an upside-down cup concealing a small certain reward, and multiple upside-down cups, only one of which hid a larger reward with a given probability [16,20]. In the third paradigm, subjects exchanged a lower-value food item with a human experimenter to obtain a larger reward under probabilistic conditions [21,22]. These controlled experimental approaches provided valuable insights, yet it remains unclear whether their findings extend to wild populations. Previous research has extensively examined probabilistic inferences [19], foraging decision-making [23], and predatory risk perception [24] in wild non-human primates, but no studies to date have investigated decision-making under risk in wild populations using paradigms comparable to those applied in captive settings.
We tested a sample of 33 Balinese long-tailed macaques (Macaca fascicularis fascicularis) from a free-ranging, urban-dwelling, habituated, provisioned population with a diverse age and sex composition, employing a risky-choice task. Subjects were tested opportunistically by presenting each individual with a series of choices between a safe option (a single upside-down colored cup consistently concealing a single food item) and a risky option (four upside-down colored cups, one of which concealed four food items). To confirm that choices in the main task reflected an understanding of the contingencies rather than chance, control trials were conducted in which macaques observed the baiting process, making clear which risky cup contained the larger reward.
Long-tailed macaques are a good primate model for studying decision-making under risk because they combine several advantages that are rare to find in another species. First, in terms of behavioral ecology, they are opportunistic foragers, frequently facing unpredictable situations in the wild that require balancing risk against potential payoff [25,26,27]. Second, in terms of cognitive flexibility, they can adapt rapidly to a changing environment [28]; they perform well in experimental tasks involving categorization [29]; they show varying degrees of inequity aversion with different cost–benefit ratios [30]; and they display relatively long waiting abilities in exchange task [22]. Third, they show temperamental characteristics and stable personality variations that are relevant to investigating their risk preferences [31,32].
Because our study population was provisioned with food daily, it was buffered from major temporal and spatial variability in food abundance. Still, studies showed that long-tailed macaque groups relying on food provided by humans also change their movement and feeding patterns depending on human presence and activities (e.g., food offerings during religious ceremonies, visitors bringing food to the site), which could be considered a proxy for unpredictable food availability [33]. Moreover, all our studied subjects routinely and spontaneously engage in token-mediated bartering interactions with humans that typically involve the following three steps: (1) a monkey steals a valued object from a visitor to the temple (e.g., glasses, hats, jewelry); (2) the monkey waits for one of the temple staff to provide a food offering in exchange for the object; (3) when the monkey deems that the food offering is sufficient, it provides the object to the temple staff who then returns it to the visitor [34]. Such unusual behavioral sequences involve risk-sensitive decisions pertaining to food procurement.
According to the foraging hypothesis, which posits that ecological pressures are crucial drivers of cognitive evolution in primates [6], the ecological and behavioral characteristics of long-tailed macaques would lead us to expect a risk-prone attitude when individuals choose between a safe option and a risky option with the same expected value. However, to our knowledge, the only study that has investigated the risk preference of long-tailed macaques reported a neutral risk attitude in two captive individuals tested using a repeated information-sampling task, in which they could interact with an automatically operating food dispenser [35]. Consequently, precise predictions based on the existing literature are difficult to formulate, and the present study should therefore be regarded as exploratory. Nevertheless, by drawing on a larger sample of wild individuals, the findings of this study will contribute to further elucidating the factors influencing decision-making under risk within the Primate order.
2. Materials and Methods
2.1. Ethical Note
All subjects were well habituated to humans and tested opportunistically. Macaques could participate voluntarily by approaching the experimenter and stop participating at any time by just moving away. The research was non-invasive, and all tests were motivated by positive reinforcement. The study was conducted in accordance with the Indonesian Ministry of Research and Technology (research permits: 253/SIP/IV/FR/10/2022 and 325/SIP/IV/FR/5/2024), the Provincial Government of Bali and the local district authorities. It was approved by the institutional Animal Welfare Committee of the University of Lethbridge (Protocol number 2201).
2.2. Study Population and Site
We studied a population of free-ranging, urban-dwelling, habituated, and provisioned Balinese long-tailed macaques living within and around the Uluwatu Temple (8°49′ S, 115°05′ E), located in a dry agricultural landscape of southern Bali, Indonesia. This is a Hindu temple complex used by Balinese communities for daily religious ceremonies. It is also one of the most famous tourist spots on the island, visited by 1.5 million tourists in 2015 [36]. A population of long-tailed macaques has lived in this anthropogenic habitat for decades [37]. At the time of the study, the population totaled approximately 400 individuals and was composed of five neighboring groups with overlapping home ranges [36]. The monkeys have been provisioned by the temple staff sporadically since 1999, and regularly since 2010 with various fruits and vegetables widely distributed across the site [38]. They also receive fruits and peanuts from tourists.
2.3. Subjects
We tested 33 individually identified subjects, including six adult females (>4 years old), seven adult males (>6 years old), three subadult females (3–4 years old), eight subadult males (4–6 years old), two juvenile females (1–3 years old), and seven juvenile males (1–4 years old). Even though the exact age and genealogy of studied subjects were not known, we determined age–sex classes by visual assessment of body size and proportion, dental development, and sexual maturity features (see [39] for the categorization of age and sex classes in Balinese long-tailed macaques). Individual identification was achieved by using a combination of natural physical markers, such as facial features, body size and proportions, fur coloration and patterns, as well as long-lasting scars, wounds, and deformities.
2.4. General Procedure
Testing was carried out during two field seasons (November 2022–January 2023 and June–August 2024) and occurred between 8:00 a.m. and 6:00 p.m. The subjects were tested opportunistically whenever the experimenter encountered one or more individuals and at least one individual was willing to approach the experimental apparatus. Throughout the study, the food item unit used as a reward was one slice of sweet banana, locally known as Pisang raja. A conditioning phase preceded experimental trials for macaques that had not interacted with experimental apparatus, wherein a food item was placed on the board in view, and the macaque was allowed to freely approach the board and take the food item (see paragraph 2.5 for a description of the experimental apparatus).
The study consisted of an Experimental phase and a Control phase. In both phases, one experimenter positioned on the ground and in front of an approaching macaque a wooden board on which there were (a) a single upside-down colored cup always covering one food item and (b) four upside-down differently colored cups, one of which covered four food items. Then, the experimenter waited until the macaque chose one of the cups by either touching or lifting it. During the choice procedure, the experimenter did not look at any cup to prevent unintentional cues. A second experimenter video recorded all trials.
2.5. Experimental Apparatus
In both Experimental and Control phases, the apparatus was a wooden board (measuring 60 × 10 cm). Attached to the board was a layer of poster board on which five dowel rods (measuring 30 cm) were evenly spaced across the board. Upside-down cups were positioned on the dowel rods to allow the macaques to lift them up to obtain the food item. The cups were all attached to the dowel rod (measuring 60 cm) by dental floss to allow the experimenter to lift all five cups at the beginning and end of each trial. The board was placed on the ground, in front of the experimental subject. In both phases, there were a single upside-down, black-striped light blue cup always covering one food item and four upside-down differently colored cups, one of which covering four food items. Due to material availability in Bali, in the 2022 field season we used four green cups, whereas in the 2024 field season we used four red cups. Each cup measured 8.9 cm in diameter and 12.7 cm high. To introduce uncertainty in the Experimental phase, the experimenter concealed baiting from the experimental subject’s view by means of an opaque screen. In the 2022 field season, cardboard was used for the screen and in the 2024 field season a wooden screen was used.
2.6. Experimental Phase
In the Experimental phase, macaques were presented with a series of choices between a safe option (a single upside-down, black-striped light blue cup always covering one food item) and a risky option (four upside-down green or red cups, one of which covering four food items). The safe option was consistently placed at either end of the row, with its order counterbalanced across trials. The expected value (EV) was the same for both options. For the risky option, there was a 0.25 probability (P) of receiving four food items (V). Thus, the EV was 0.25 × 4 = 1, which matched the EV of the safe option, yielding one food item with a probability of 1.
Before baiting the cups, the experimenter lifted the five cups to show the macaque each upside-down cup was empty. The experimenter then showed the macaque subject the potential food amounts associated with each option (one and four food items, respectively) in their open hands. After the subject had looked at both food amounts, the experimenter placed those under the respective upside-down cups. The experimenter placed the single food item under the cup representing the safe option in full view of the subject, whereas they concealed the baiting procedure of the four food items, which were placed under one of the four cups, by means of an opaque wooden or cardboard screen. The hiding position of the four food items was pseudo-randomized across trials. Once the screen covering the risky option was removed, the subject could select their preferred cup by touching or lifting it. After each rewarded choice, there were two possible scenarios. In the first one (which occurred most of the time), the subject took the food reward off the board after lifting the cup. In the second scenario, if the subject did not take the food reward, the experimenter provided it. The experimenter also revealed the content of the non-chosen option, allowing the subjects to see where the food items were located. There was an inter-trial interval of about 15 s. Supplementary Material Video S1 shows a typical experimental session during which the subject chose the risky option, and Supplementary Materials Video S2 shows a typical experimental session during which the subject chose the safe option (https://osf.io/yh8mr/files/95umw, 10 February 2026).
We usually carried out one experimental session per day. Most subjects (N = 27) participated in one session, three subjects in two sessions, two subjects in three sessions, and one subject in five sessions. Each session had an average of 11.93 ± 0.21 trials (minimum 9 trials–maximum 13 trials). Further details are reported in Table 1.
2.7. Control Phase
The Control phase aimed to assess whether macaques consistently chose the cups covering the largest food option when they could observe the baiting procedure. This ensured that their choices in the Experimental phase were influenced by the uncertainty of obtaining the better payoff rather than by other factors. The only difference between the Control phase and the Experimental phase was that, in the Control phase, the experimenter baited also the risky option in full view of the subject. This ensured that macaques could see, before making their choice, under which cup the four food items were placed.
We carried out the Control phase after the Experimental phase on 24 out of the 33 macaques, which participated in the Experimental phase during the 2024 field season. Most subjects (N = 22) participated in one session, one subject in two sessions, and one subject in three sessions. Each session included a single choice between the single upside-down, black-striped light blue cup covering one food item and the four upside-down red cups covering four food items.
2.8. Data Analysis
To assess whether macaques significantly preferred either option above chance level, we employed the Binomial test at the individual level and the single-sample Wilcoxon signed-ranks test at the population level.
Next, to evaluate whether sex, age, trial number, and previous trial outcome (zero, one, or four food items) were related to risky choices, we implemented a random-effects logistic regression model. The identity of each subject was included as a random effect. Interactions were analyzed using the Wald test; non-significant interactions were removed from the model before repeating the analysis. The significance level was set at p < 0.05. We conducted all analyses using Stata 17.0.
3. Results
3.1. Experimental Phase
The average proportion of choices for the risky option did not differ significantly from random choice (mean ± SE: 0.41 ± 0.29, z = −1.662, p = 0.097). As reported in Table 1, at the individual level, six macaques were risk-prone, 15 macaques were indifferent between options, and 12 macaques were risk-averse. Specifically, two adult females were risk-prone and four were indifferent (overall prone: z = 3.481, p exact < 0.001); one adult male was risk-prone, three were indifferent, and three were averse (overall indifferent: z = −1.949, p exact = 0.064); two subadult females were risk-prone, one was averse (overall indifferent: z = 1.333, p = 0.243); two subadult males were indifferent and six were averse (overall averse: z = −8.803, p exact < 0.001); one juvenile female was averse and the other one was indifferent (overall averse: z = −2.667, p exact = 0.011); one juvenile male was risk-prone, five were indifferent, one was averse (overall indifferent: z = 0.863, p exact = 0.450).
As shown in Figure 1 and Figure 2, there were significant interactions between age and sex (Chi square = 11.23, p = 0.004, df = 2), and between age and previous trial outcome (Chi square = 15.91, p = 0.003, df = 4); however, the triple interaction between age, sex and previous trial outcome was not significant (Chi square = 2.82, p = 0.588, df = 4).
Among males, adults (marginal mean ± SE: 0.41 ± 0.10) and juveniles (0.52 ± 0.10) showed a significantly higher risk preference than subadults (0.16 ± 0.04) (adults vs. subadults: coeff. = 1.495, z = 2.39, p = 0.017; juveniles vs. subadults: coeff. = 2.031, z = 3.23, p = 0.001); among females, adults (0.62 ± 0.08) were significantly more risk-prone than juveniles (0.33 ± 0.02) (coeff. = 1.484, z = 3.53, p < 0.001).
Juveniles chose the risky option significantly more after having previously chosen a risky option producing a four-food-item outcome (0.75 ± 0.08) compared to a risky option with no food outcome (0.47 ± 0.08) or a safe option (0.38 ± 0.08) (risky with four-food-item outcome vs. no food outcome: coeff. = 1.495, z = 3.01, p = 0.003; risky with four-food-item outcome vs. safe: coeff. = 1.957, z = 3.09, p = 0.002). There were no other statistically significant comparisons.
3.2. Control Phase
When the subjects could observe the baiting procedure, and thus were aware of which risky cup concealed the larger food amount, they selected the cup covering four food items in 0.81 ± 0.08 of the trials, on average.
4. Discussion
In the present study, we tested a sample of 33 Balinese long-tailed macaques from a free-ranging, urban-dwelling, habituated, provisioned population in a risky-choice task. Overall, macaques exhibited indifference between the safe option (yielding one food item) and the risky option (corresponding to a 25% chance of receiving four items). However, we found important interindividual differences related to age–sex interactions. Specifically, adult and juvenile males were more risk-prone than younger adult males, and adult females were more risk-prone than juvenile females. Additionally, regardless of sex, juvenile macaques were sensitive to the outcome of their prior choices compared to adults. More specifically, they showed a “win–stay” strategy, by selecting the risky option more often after having previously chosen a “winning” risky option compared to a “losing” risky option or a safe option.
Macaques’ indifference between the safe and the risky options may be due either to a lack of understanding of the contingencies of the task or to economic rationality, as the two options had the same expected value. The control trials performed on a subset of subjects, in which macaques yielded 81% accuracy in identifying rewarded cups when they could witness the baiting procedure, dismissed the possibility that they did not understand the task. Thus, it seems more likely that, on average, this population of long-tailed macaques showed genuine risk neutrality. Notably, at the individual level, indifference between options was the pattern most frequently observed, notwithstanding striking interindividual differences. To our knowledge, the only other study investigating attitude towards risk in two long-tailed macaques tested in a repeated information-sampling task showed a neutral risk preference [35].
However, the risk neutrality of long-tailed macaques contrasts with the risk-seeking behavior observed in most of the captive studies on rhesus macaques carried out so far [8,9,40]; but see [7,41]. Several factors may account for the different risk preferences of the two species. First, rhesus and long-tailed macaques, while phylogenetically closely related [42], show some differences in social tolerance and temperament: rhesus macaques are slightly more despotic than long-tailed macaques [43,44], and rather aggressive and unsociable towards humans, whereas long-tailed macaques are more cautious and fearful [32]. Additionally, when tested in a battery of inhibitory control tasks, rhesus macaques showed worse performance than long-tailed macaques ([45]; see also [46] for delay of gratification in long-tailed macaques and [47] for a critical discussion), which may explain their higher risk preferences, although there are mixed results regarding the association between risk preferences and delay preferences in both human and non-human primates [48,49]. Second, captive rhesus macaques are usually presented with repeated situations whose contingencies must be learned by experience, which produce a systematic bias towards risk-proneness [50,51]. In contrast, we tested wild long-tailed macaques with a risky-choice paradigm in which the probability distribution was visually described by the four cups, only one of which contained the larger reward. Third, captive rhesus macaques tested in risky-choice tasks are usually food-deprived to increase their motivation, and thus in a low energetic state, leading to higher risk proneness according to risk-sensitivity theory [1]. By comparison, the long-tailed macaques sampled in our study were not only free to forage, but they were also provisioned, and this may have increased their energetic state, leading them to be overall indifferent between options rather than risk prone as captive rhesus macaques. Finally, whereas captive studies are usually carried out on a few individuals, here we tested a rather large population of macaques and, despite the overall risk neutrality, we found notable interindividual differences. This variability suggests that the observed interspecific differences in risk preference may at least in part reflect interindividual variation within species.
In the present study, adult and juvenile males showed a more pronounced risk-seeking attitude than younger adult males, and adult females were more risk-prone than juvenile females. These findings are not consistent with the few reports on the effect of age on risk attitude in non-human primates, and particularly studies in chimpanzees showing that adolescents and young adults exhibited greater risk proneness than older adults [10,17]. There may be different motives beyond the higher risk preference of juvenile and adult males, which were overall risk neutral, compared to younger adult males, which were risk-averse. Juveniles’ choices may be guided by impulsive tendencies and low awareness of the potentially negative consequences of their behavior [52]. In contrast, adult males may be bolder than younger males because of their generally higher explorative and competitive attitude in social, sexual, and foraging contexts [53,54]. Similarly, the higher risk proneness of adult females, as compared to juvenile females, contrasts with the typical females’ risk-aversion behavior reported in the literature [5]. There are at least two related explanations for this deviation from the usual pattern. First, adult females’ reproductive burden is likely to increase their energy needs compared to immature females [55]. Second, according to the abundance/risk hypothesis [56], food provisioning of our studied macaque population may have favored risk proneness among adult females. Interindividual differences were, however, conspicuous, and it cannot be excluded that a few individuals have driven the group effect.
Finally, regardless of sex, juvenile macaques were sensitive to the outcome of their prior choices, showing a short-term “win–stay” strategy, i.e., selecting the risky option more often after having just chosen, in the previous trial, a “winning” risky option rather than a “losing” risky option or a safe option. Win–stay/lose–shift-like heuristics are often observed in non-human primates and other animals tested in risky-choice tasks [57]; reviewed in [39]. However, this is the first study, to our knowledge, in which juveniles weighted their recent positive outcomes more so than older individuals, possibly because the rewarding risky choice reinforced their exploratory tendencies. Although the risky-choice task was novel for all macaques, the different past experiences with ecological contingencies of juvenile and older individuals may have played a role, leading older individuals to adopt choice strategies less influenced by recent positive outcomes. This hypothesis is supported by the observation that the win–stay/lose–shift heuristic is not consistent across different species and populations of the same species. Some studies identified sensitivity to previous choice outcomes in rhesus macaques tested with risky-choice tasks [58,59], whereas others reported interindividual differences [60]. Additionally, when tested with the same paradigm, bonobos modulated their risky choices based on previous outcome, whereas chimpanzees did not [57] (see also [61] for similar negative findings in another risky-choice paradigm). Similarly, two colonies of tufted capuchin monkeys exposed to slightly different feeding schedules were differently sensitive to previous risky-choice outcomes [3,14]. Taken together, these findings indicate that reliance on recent positive outcomes varies across primate species and populations, likely reflecting differences in life history and ecological features.
While the present study offers important insights into risky decision-making in non-human primates, a specific methodological limitation should be acknowledged. Because our data collection was based on the spontaneous and voluntary participation of free-ranging individuals in the proposed choice task, our results may have been biased via a possible self-selection of study subjects. Indeed, several field experimental studies conducted in wild primates have shown that personality traits lead to systematic interindividual differences in task engagement and learning in natural environments. Specifically, bolder, more active, and more exploratory individuals are more likely to approach, interact with, and solve human-induced problems than their shyer and generally more inhibited counterparts [62,63,64]. Even though this issue is inherent to most field experiments, we cannot rule out an oversampling of bolder monkeys in our study, which could have impacted the generalizability of our results on risk preferences. For example, such a self-selection process could explain the preponderance of male subjects among juveniles and subadults (Table 1), that is, age categories in which sex differences in boldness and risk-taking are often more pronounced than in older individuals, particularly in macaques [65,66].
5. Conclusions
In conclusion, this study provided the first examination of risk preferences in free-ranging Balinese long-tailed macaques, extending laboratory paradigms on risky choices to field settings, and thus bridging a critical gap in achieving a more complete understanding of risk attitude in a more ecologically valid setting. Our findings revealed that this urban-dwelling population exhibited overall risk neutrality in a descriptive risky-choice task. This contrasts with the risk-prone tendencies of captive rhesus macaques and highlights the potential influence of ecological context, provisioning, and species-specific features on decision-making under risk. Interindividual variability driven by age–sex interactions underscores that risk preferences are potentially affected by a complex interplay of factors, such as developmental stage, reproductive demands, and experience, which all shape risk attitude, and highlight the importance of considering demographic factors when characterizing species-typical risk preferences. These findings call for future research on decision-making under risk in a longitudinal perspective in a wide variety of captive and free-ranging non-human primate populations, possibly experiencing different degrees of food availability, to provide a framework for reconciling inter- and intra-specific variability in non-human primate attitude towards risk.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Kacelnik A. Bateson M. Risky theories—The effects of variance on foraging decisions Am. Zool.19963640243410.1093/icb/36.4.402 · doi ↗
- 2Stevens J.R. Stephens D.W. The adaptive nature of impulsivity Impulsivity: The Behavioral and Neurological Science of Discounting Madden G.J. Bickel W.K. American Psychological Association Washington, DC, USA 2010361387
- 3De Petrillo F. Ventricelli M. Ponsi G. Addessi E. Do tufted capuchin monkeys play the odds? Flexible risk preferences in Sapajus spp.Anim. Cogn.20151811913010.1007/s 10071-014-0783-724993065 · doi ↗ · pubmed ↗
- 4Heilbronner S.R. Rosati A.G. Stevens J.R. Hare B. Hauser M.D. A fruit in the hand or two in the bush? Divergent risk preferences in chimpanzees and bonobos Biol. Lett.2008424624910.1098/rsbl.2008.008118364305 PMC 2610056 · doi ↗ · pubmed ↗
- 5De Petrillo F. Rosati A.G. Variation in primate decision-making under uncertainty and the roots of human economic behaviour Philos. Trans. R. Soc. B 20213762019067110.1098/rstb.2019.0671 PMC 781543433423637 · doi ↗ · pubmed ↗
- 6Rosati A.G. Foraging cognition: Reviving the ecological intelligence hypothesis Trends Cogn. Sci.20172169170210.1016/j.tics.2017.05.01128625354 · doi ↗ · pubmed ↗
- 7Eisenreich B.R. Hayden B.Y. Zimmermann J. Macaques are risk-averse in a freely moving foraging task Sci. Rep.201991509110.1038/s 41598-019-51442-z 31636348 PMC 6803699 · doi ↗ · pubmed ↗
- 8Farashahi S. Azab H. Hayden B.Y. Soltani A. On the flexibility of basic risk attitudes in monkeys J. Neurosci.2018384383439810.1523/JNEUROSCI.2260-17.201829626169 PMC 5932645 · doi ↗ · pubmed ↗