Jaceidin Suppresses Melanoma Metastasis by Modulating ERK, JNK Pathways and Extracellular Matrix Degradation Proteins
Mu‐Kuei Shieu, Hui‐Ju Yang, Chia‐Chieh Lin, Min‐Yun Kao, Hsin‐Yu Ho, Yu‐Sheng Lo, Yi‐Ching Chuang, Yuan‐Ting Yang, Ming‐Ju Hsieh

TL;DR
Jaceidin, a flavonoid compound, shows anti-melanoma effects by inhibiting cancer cell metastasis through specific molecular pathways.
Contribution
This study reveals jaceidin's novel anti-metastatic effects on melanoma cells via ERK, JNK pathways and extracellular matrix proteins.
Findings
Jaceidin significantly inhibits metastasis in HMY-1 and A2058 melanoma cell lines.
Jaceidin modulates ERK and JNK phosphorylation and suppresses epithelial-mesenchymal transition proteins.
Jaceidin reduces expression of MMP-2 and cathepsins A, which degrade extracellular matrix.
Abstract
Metastatic melanoma, marked by its poor prognosis and frequent recurrence, is a particularly aggressive skin cancer. Jaceidin, a compound derived from flavonoids abundant in common fruits and vegetables, has attracted interest for its potential anti‐cancer properties. Nevertheless, the specific effects of jaceidin on melanoma cells remained unclear before this study. Here, we examined the impact of jaceidin on two metastatic melanoma cell lines, HMY‐1 and A2058. Our results demonstrate that jaceidin exerts a significant anti‐metastatic effect against both cell lines. This inhibitory action involved modulating the phosphorylation of extracellular signal‐regulated kinase and c‐Jun N‐terminal kinase, as well as suppressing proteins associated with epithelial‐mesenchymal transition. Furthermore, jaceidin reduced the expression of extracellular matrix degradation proteins MMP‐2 and…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
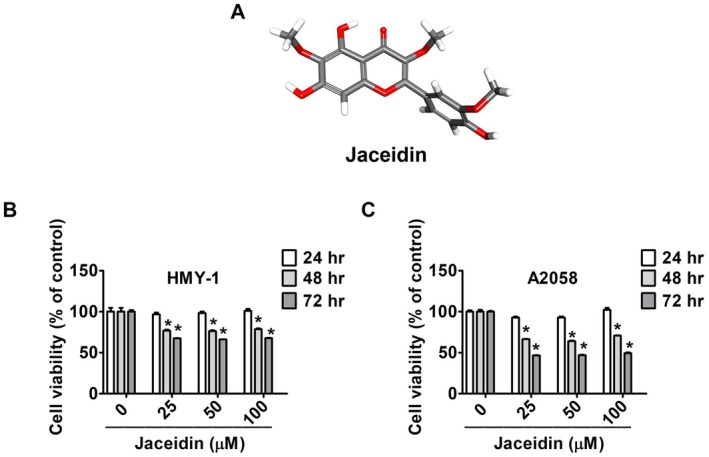 FIGURE 1
FIGURE 1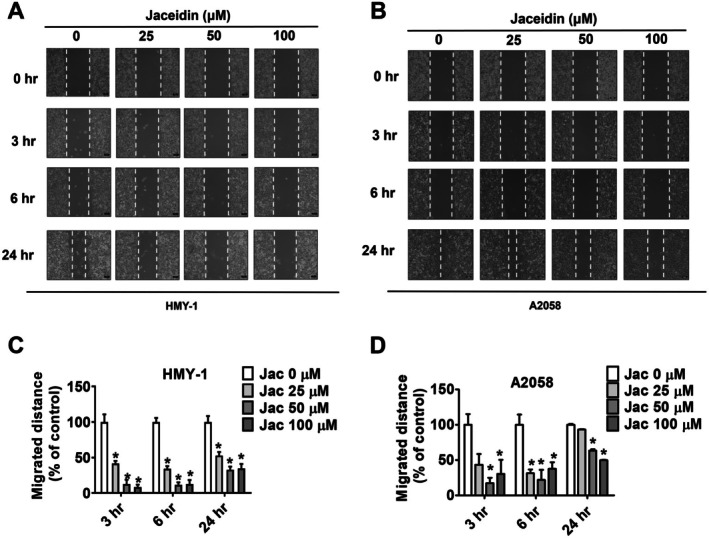 FIGURE 2
FIGURE 2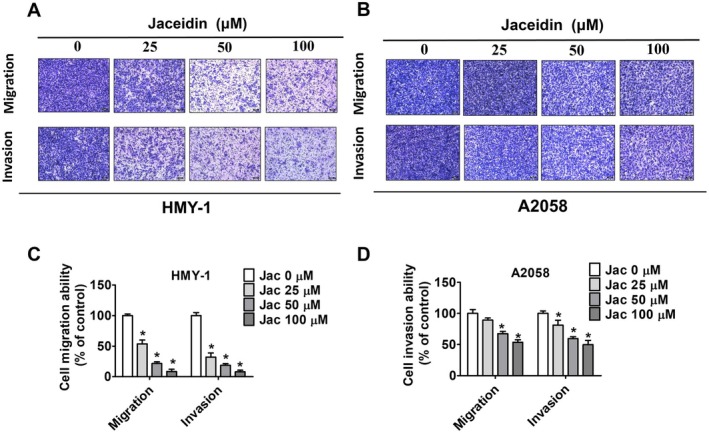 FIGURE 3
FIGURE 3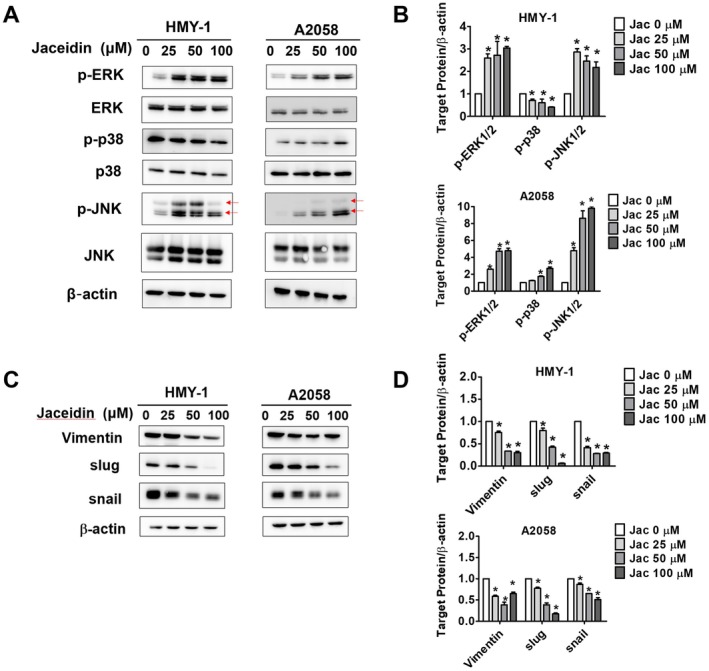 FIGURE 4
FIGURE 4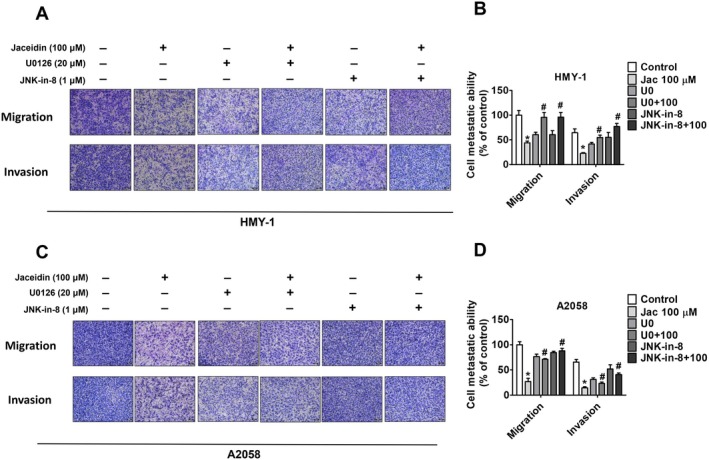 FIGURE 5
FIGURE 5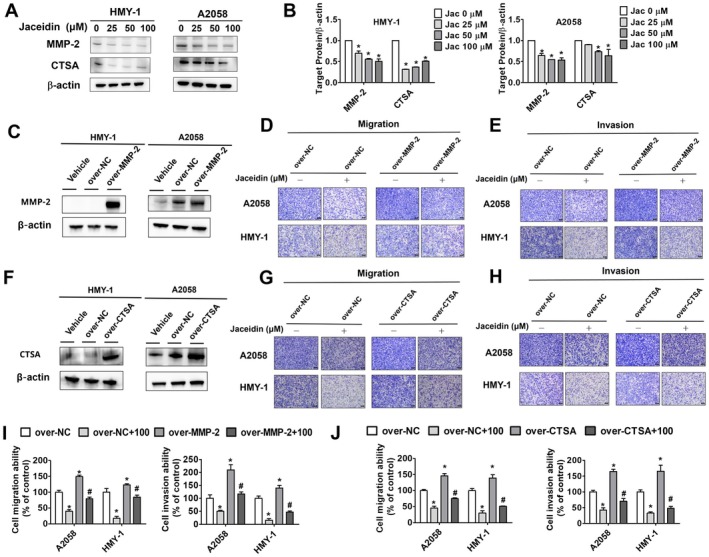 FIGURE 6
FIGURE 6- —Changhua Christian Hospital10.13039/501100007632
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsFlavonoids in Medical Research · Melanoma and MAPK Pathways · melanin and skin pigmentation
Introduction
1
Among skin cancer, cutaneous melanoma has remained the most significant global health concerns for decades. In 2024, it is estimated that 100,000 new cases will be diagnosed in the United States alone [1]. Additionally, the annual incidence of melanoma has been steadily increasing in most countries, according to the World Health Organization (WHO) database [2]. Various risk factors have been identified in the development of melanoma, including over‐exposure to ultraviolet, family history of the disease, genetic predisposition and ethnicity, with white individuals being particularly at risk [3, 4]. Furthermore, melanoma can be diagnosed at any age, with cases reported in both neonates and the elderly [5].
While most of the melanoma tumour are classified as primary stages (I and II) [6], they can typically be treated effectively through wide local excision [7], with survival rate extending to 10 years or longer [8]. In contrast, once advanced stage of melanoma (III and IV) was diagnosed, it becomes a significant challenge for both clinicians and patients due to its highly metastatic rate and recurrence. According to previous studies, before the systematic treatment was invented, the median survival of advanced melanoma only lasted for 6 months [9]. Fortunately, recent years have seen the emergence of several new treatment options, including the application of immunotherapy (e.g., Cytotoxic T‐lymphocyte‐associated antigen‐4, Programmed Cell Death Protein 1, or Programmed Cell Death Ligand 1), target therapy (e.g., BRAF inhibitors) or chemotherapy. While these therapies have led to significant improvements in survival rates [10], the adverse effects, including high toxicity, remain a considerable concern [11, 12]. Furthermore, the following financial cost during therapeutic course has become a burden for both patients and their families [13].
Fortunately, increasing research efforts are being directed towards identifying new therapeutic pathways to combat melanoma. One such focus is the epithelial–mesenchymal transition (EMT), which plays a critical role in melanoma progression and is closely associated with enhanced cell motility and invasiveness [14]. For instance, one study demonstrated that green tea polyphenols can inhibit the proliferation of A875 and SK‐MEL‐1 melanoma cells by downregulating EMT‐related gene expression [15, 16], Another study found that long pentraxin‐3, a component of the fibroblast growth factor family, suppresses the proliferation of A375 and A2058 melanoma cells through similar downregulation of EMT markers [17].
In the present study, we focused on an organic compound called jaceidin as our main target against melanoma cells. Jaceidin is a member of the flavonoid family, which is commonly found in fruits and vegetables [18]. Flavonoids can be categorized into several subclasses [19], with some derivatives documented to exhibit anti‐cancer properties, particularly in inhibiting invasion and migration in various cancer cells [20]. For example, in gastric cancer cells, a compound called jaceosidin was shown to suppress cell migration by activating reactive oxygen species‐mediated signalling pathways [21]. In cervical cancer cells, a synthetic compound, WYC02‐9, was found to promote apoptosis and inhibit cell migration by activating the mitogen‐activated protein kinase 14 pathway [22]. One previous study highlighted jaceidin's ability to suppress ascites carcinoma progression in mice, resulting in a significant reduction in tumour weight [23]. Furthermore, jaceidin and other flavonoid derivatives have been shown to exert anti‐carcinogenic properties by inducing cell apoptosis in breast cancer [24].
To date, however, the effects of jaceidin on melanoma cells remain unexplored. Therefore, this study aims to investigate the potential chemotherapeutic effects of jaceidin on the invasion and migration of malignant melanoma cells.
Material and Methods
2
Cell Culture
2.1
Two melanoma cell lines, HMY‐1 (BRAF wild type) and A2058 (BRAF mutant), were selected as the target cells for this study. These cell lines were derived from metastatic lymph nodes of a 62‐year‐old male and a 43‐year‐old male [25, 26]. Both lines were obtained from the Japanese Collection of Research Bioresources (Tokyo, Japan). The cells were cultured in Dulbecco's Modified Eagle Medium (Gibco BRL, Grand Island, NY, USA), supplemented with 10% foetal bovine serum and 1% penicillin/streptomycin. The cells were then incubated at 37°C in a 5% CO_2_ atmosphere under humidified conditions.
Chemical Treatments
2.2
Jaceidin (C18H16O8, molecular weight: 360.31, ≥ 96% purity) was purchased from ChemFaces (Wuhan, China). A stock solution of jaceidin (100 μM) was prepared using dimethyl sulfoxide (DMSO) and stored at −20°C for future use. Based on the required treatment doses, the stock solution was serially diluted to prepare working solutions. The final concentration of DMSO in all treatments was maintained at less than 0.1%.
MTT Assay
2.3
To assess cell viability, melanoma cells were cultured in 24‐well plates and treated with 25, 50, or 100 μM jaceidin for 24 h. Untreated cells served as the control group. After incubation, the MTT assay (3‐(4,5‐dimethylthiazol‐2‐yl)‐2,5‐diphenyltetrazolium bromide) was performed to evaluate cell viability. DMSO was then added to dissolve the purple crystals, and absorbance was measured using a BioTek spectrophotometer (Winooski, VT, USA) at 570 nm.
Wound Closure Assay
2.4
Two melanoma cell lines (HMY‐1 and A2058) were seeded into 12‐well culture plates and incubated overnight to reach 90% confluence. After creating a wound using a tip, the cells were incubated with different concentrations of jaceidin (25, 50, 100 μM) for 3, 6 and 24 h. Wound photographs and the crawling distance of cells were recorded under a microscope.
Cell Migration and Invasion Assay
2.5
To analyse cell migration and invasion, transwell assays were performed using transwell inserts. The cells were treated with different concentrations of jaceidin (25, 50 and 100 μM) for 24 h. After 24 h of incubation, the cells were fixed with methanol for 10 min, then dried. The cells were stained with 10× Giemsa for 4 h, and the non‐migrating cells on the upper surface of the membrane were removed. The number of migrating cells was then counted under a 100X microscope. For the invasion assay, the procedure was repeated with a 10‐fold dilution, and the steps were the same as the migration assay, and diluted Matrigel gel (25 mg/50 mL; 60 μL; BD Biosciences) was coated on the upper transwell at 37°C for overnight.
Western Blot Assay
2.6
For Western blot analysis, protein samples were extracted using lysis buffer. The samples were then separated by 10% polyacrylamide gel electrophoresis and transferred to polyvinylidene fluoride (PVDF) membranes (Millipore Corporation, Milford, MA). The membranes were blocked with milk for 1 h, followed by incubation with primary antibodies for 24 h. Afterward, the membranes were incubated with secondary antibodies at room temperature for 1 h. Finally, protein bands were visualized using the ImageQuant fluorescence biomolecule imaging system.
Statistical Analysis
2.7
Statistical analysis was conducted using SigmaPlot v12.5 (Systat Software; Palo Alto, CA, USA). A p < 0.05 was considered statistically significant. One‐way analysis of variance (ANOVA) followed by Tukey's multiple comparison test was used to compare the treated cells with the control group.
Results
3
Effect of Jaceidin on Cell Viability in Melanoma Cell Lines
3.1
Figure 1A shows the chemical structure of jaceidin. To investigate the cytotoxicity of jaceidin, the HMY‐1 and A2058 melanoma cell lines were treated with different concentrations (25, 50 and 100 μM) of jaceidin for 24, 48 and 72 h. MTT assays were performed as shown in Figure 1B,C. DMSO treated cells served as the control group. The results indicated that jaceidin concentrations of 25, 50 and 100 μM did not exhibit cytotoxicity after 24 h of treatment. However, significant cytotoxicity was observed after 48 and 72 h of treatment, in comparison to the DMSO treated group. These findings suggest that jaceidin effectively inhibits the viability of melanoma cells, with prolonged exposure leading to enhanced cytotoxic effects.
*Toxicity of jaceidin against melanoma cells. (A) Chemical structure of jaceidin. (B, C) MTT assay of HMY‐1 and A2058 cells treated with different concentrations of jaceidin (0, 25, 50 and 100 μM) for 24, 48 and 72 h. Data are mean ± SD of three independent experiments. p < 0.05, compared with vehicle.
Effect of Jaceidin on Cell Motility in Melanoma Cell Lines
3.2
To evaluate cell motility, a wound healing assay was performed. As described earlier, HMY‐1 and A2058 cells were treated with 25, 50 and 100 μM of jaceidin for 3, 6 and 24 h, respectively. As shown in Figure 2, jaceidin reduced the migration distance of both melanoma cell lines in a dose‐dependent manner.
*Jaceidin inhibited horizontal migration of melanoma cells. (A, C) HMY‐1 cells treated with indicated concentrations of jaceidin (0, 25, 50 and 100 μM) and photographed at 0, 3, 6 and 24 h to measure distance migrated. (B, D) A2058 cells treated with indicated concentrations of jaceidin (0, 25, 50 and 100 μM) and photographed at 0, 3, 6 and 24 h to measure distance migrated. Data are mean ± SD of three independent experiments. p < 0.05, compared with vehicle.
Effect of Jaceidin on Invasion and Migration in Melanoma Cell Lines
3.3
In our study, the effects of jaceidin on cell invasion and migration were assessed using the transwell assay. As shown in Figure 3, jaceidin treatment significantly inhibited both invasion and migration in both melanoma cell lines, with a particularly notable reduction observed in HMY‐1 cells. Overall, these findings suggest that jaceidin exerts anti‐metastatic effects on malignant melanoma cell lines.
*Jaceidin inhibited migration and invasion through melanoma cells (A, C) migration and invasion assays for HMY‐1 cells (B, D) migration and invasion assays for A2058 cells. Cells stained with Giemsa staining buffer were counted to analyse cell migration and invasion. Data are mean ± SD of three independent experiments. p < 0.05, compared with vehicle.
Jaceidin Reduces Protein Expression in the MAPK Pathways of Melanoma Cell Lines
3.4
The MAPK pathway is well‐known for its strong association with cell proliferation and migration [27]. To explore the signalling pathways involved in the effects of jaceidin on cell migration, three key MAPK pathways were selected as targets: extracellular signal‐regulated kinase (ERK), p38 and c‐Jun N‐terminal kinase (JNK) [19]. In this study, as the concentration of jaceidin increased, the phosphorylation of ERK and JNK pathways significantly increased in both melanoma cell lines. In contrast, the phosphorylation of the p38 pathway decreased in HMY‐1 cells, with no statistically significant change observed in the A2058 cell line (Figure 4A,B). As previously mentioned, EMT plays a crucial role in enhancing cell mobility and invasion [28, 29]. Therefore, we examined the expressions of EMT‐related proteins, including vimentin [30], slug [31] and snail [32]. Our results showed that jaceidin reduced the expression of these proteins (Figure 4C,D). These findings suggest that jaceidin inhibits melanoma cell metastasis by modulating the phosphorylation of ERK, JNK, as well as downregulating EMT‐related proteins.
*MAPK pathways involved in jaceidin‐induced metastasis regulation in melanoma cells. (A, B) After jaceidin treatment, total and phosphorylated ERK, p38 and JNK levels in HMY‐1 and A2058 cells were measured using Western blot assay with β‐Actin as an internal control. (C, D) Expressions of EMT‐related proteins (vimentin, slug and snail) were measured using Western blot assay with β‐Actin as an internal control. Data are mean ± SD of three independent experiments. p < 0.05, compared with vehicle; #p < 0.05, compared with jaceidin only.
Effects of Jaceidin co‐Treatment With U0126 and JNK‐IN‐8 on Melanoma Cell Lines
3.5
To further investigate the involvement of the ERK and JNK pathways in cell migration and invasion, small molecule inhibitors—U0126 (an ERK inhibitor) and JNK‐IN‐8 (a JNK inhibitor)—were used. As shown in Figure 5, the results demonstrated that, in HMY‐1 cells, co‐treatment with jaceidin, U0126 and JNK‐IN‐8 significantly enhanced migration and invasion compared to the control groups. Similarly, in A2058 cells, the co‐treatment produced comparable results. Taken together, these findings suggest that jaceidin inhibits melanoma cell metastasis by upregulating both the ERK and JNK pathways.
*ERK and JNK pathway participated in jaceidin‐induced metastasis regulation. (A, B) HMY‐1 and (C, D) A2058 cells treated with ERK inhibitor (U0126) and JNK inhibitor (JNK‐IN‐8), respectively, for 1 h. Cells were cotreated (or not) with 100 μM jaceidin for 24 h and subjected to migration and invasion assays. Data are mean ± SD of three independent experiments. p < 0.05, compared with vehicle; #p < 0.05, compared with jaceidin only.
Effects of Jaceidin on MMP and Cathepsin A Expression in Melanoma Cells
3.6
The degradation of the extracellular matrix (ECM) is a critical step in tumour metastasis [33], with both matrix metalloproteinases (MMPs) [34] and cathepsin family proteins [35] playing vital roles in this process. Among the MMP family, MMP‐2 and MMP‐9 are widely recognized as cancer biomarkers [36]. As shown in Figure 6A,B, treatment with varying concentrations of jaceidin significantly inhibited the expression of MMP‐2 and CTSA in both melanoma cell lines. The down‐regulation of MMP‐2 and CTSA indicates that both have vital roles in the antimetastatic effects of jaceidin. In Figure 6C,F, MMP‐2 and CTSA levels were up‐regulated through transfection of the MMP and CTSA plasmids. In addition, cells cotreated with over‐expression of MMP‐2 and CTSA, respectively, exhibited higher motility compared with the control group (Figure 6D,H), and the motility was inhibited after treatment with jaceidin (Figure 6I,J). Overall, the results indicated that CTSA and MMP‐2 took part in the antimetastatic properties of jaceidin in both melanoma cell lines.
*Jaceidin regulated cell metastasis by downregulating MMP‐2 and CTSA in melanoma cells. (A, B) MMP‐2 and CTSA were measured using Western blot assay after jaceidin treatment (0, 25, 50, or 100 μM), with β‐Actin as internal control. (C, D, E, I) Cells were transfected with pEGFP (empty vector) or MMP‐2 overexpression plasmid following treatment with or without jaceidin (100 μM), with β‐Actin as internal control. (F, G, H, J) Cells were transfected with pEGFP (empty vector) or CTSA overexpression plasmid following treatment with or without jaceidin (100 μM). Migration and invasion assays were used to analyse HMY‐1 and A2058 cells. p < 0.05, compared with vehicle; #p < 0.05, compared with jaceidin only.
Discussion
4
Natural products have been used as therapeutic options since ancient times [37], often under the umbrella of Traditional Chinese Medicine (TCM) or folk medicine in Asia, with many having been documented for thousands of years. Even today, numerous natural compounds continue to be explored for their medicinal potential [38]. These phytochemicals have shown distinct anti‐tumour effects against various cancers [39, 40]. In this study, we found that jaceidin, a flavonoid compound, effectively inhibited the growth of two metastatic melanoma cell lines, HMY‐1 and A2058. Our results showed that jaceidin significantly reduced melanoma cell viability. Furthermore, wound healing and transwell assays indicated that jaceidin's effects were associated with the upregulation of the ERK and JNK pathways, along with reduced expression of EMT and MMP‐related proteins. While few studies have explored the synergistic effects of jaceidin, as mentioned previously [23]. To the best of our knowledge, this is the first study to demonstrate that jaceidin inhibits cell migration and invasion in melanoma cell lines.
Among the initial steps of tumour progression, EMT is a dynamic process in which epithelial cells lose their epithelial characteristics and transform into mesenchymal cells [41]. Mesenchymal cells contribute to the promotion of tumour growth [14, 42]. This phenomenon can occur in both epithelial and non‐epithelial cancers, including melanoma [43]. As members of the transcriptional factor family, slug and snail have been shown to induce EMT in various cancer cells [44]. In ovarian cancer cells, they have been associated with increased radio resistance and chemo resistance to paclitaxel and cisplatin [45]. In B16 melanoma cells, studies have found that snail and slug are key regulators of nodal‐induced EMT in mice [46], where overexpression of nodal enhances tumour migration. Our study suggests that jaceidin is involved in downregulating the expression of EMT‐related proteins.
Considering the upstream signalling pathways in the regulation of cells, the MAPK pathway plays a crucial role in regulating various cellular processes, including cell proliferation, migration and apoptosis in mammalian cells [27, 47, 48]. We discovered that jaceidin inhibits the invasion and migration of A2058 and HMY‐1 melanoma cells by upregulating both the ERK and JNK signalling pathways. Interestingly, current BRAF inhibitors, such as vemurafenib, dabrafenib and encorafenib [49], are being used in combination with immunotherapy to target advanced stages of melanoma [50]. These inhibitors primarily target the ERK pathway, effectively inhibiting cell proliferation and improving survival outcome [51]. However, resistance mechanisms against BRAF inhibitors have been described, with reactivation of the MAPK pathway noted after approximately 12 months of treatment [52, 53]. This resistance has been specifically observed in BRAF‐mutant A2058 cell lines [54, 55]. Although our study highlights contrasting mechanisms by inhibiting melanoma cell migration through activation of the ERK and JNK pathways, other natural products have also yielded similar results. In B16F10 melanoma cells, theaflavin inhibited migration and invasion via the upregulation of JNK, p53 and downregulation of the ERK pathway [56]. Another natural compound, Gomisin A, also inhibited migration and invasion through activation of the MAPK, ERK and JNK pathways in B16F10 and A375 cell lines [57]. Conversely, Taxifolin reduced migration in B16F10 and A375 melanoma cells by inhibiting the JNK pathway [58]. Overall, it may somehow offer an alternative or combination approach for treating metastatic melanoma cells.
As previously mentioned, the activity of MMPs and cathepsin family proteins plays a crucial role in tumour growth, local invasion and metastasis [59, 60]. The degradation of the ECM is closely linked to MMP expression, which is highly correlated with tumour progression across various melanoma subtypes [61]. For instance, overexpression of MMP‐1, MMP‐2 and MMP‐13 has been observed in nodular melanoma, a subtype characterized by aggressive growth and early metastasis [62]. In uveal melanoma, patients with positive expression of MMP‐2 and MMP‐9 have been shown to exhibit lower survival rates compared to the control group [63]. Furthermore, other studies have reported higher concentrations of MMP‐2 in metastatic melanoma (stage IV) compared to primary melanoma subgroups (stage I) [64, 65]. Taken together, these findings highlight the increased detection rates of MMP‐2 in melanoma cells.
On the other hand, when considering cathepsin proteases, the cathepsin family can be classified into aspartic proteases (CTSD, E), serine proteases (CTSA, G) and cysteine proteases (CTSB, C, F, H, K, L, O, S, V, X and W) [66]. Among these lysosomal proteases, CTSA has been shown to correlate with tumour development. For example, in lung adenocarcinoma A549 cells, knockdown of CTSA decreased invasion and migration, thereby influencing cell cycle regulators [67]. In prostate cancer, elevated levels of CTSA expression have been observed in comparison to normal prostate tissues, and suppression of CTSA genes leads to inhibition of tumour growth [68]. Similarly, previous studies have demonstrated that malignant melanoma lesions exhibit higher expression of lysosomal CTSA compared to normal pigmented nevi [69, 70]. Other studies have also suggested that UV overexposure leads to the release of cathepsins from melanoma cells into the surrounding environment [71]. In the present study, jaceidin significantly attenuates the expressions of both MMP‐2 and CTSA, both of which serve as prognostic biomarkers for predicting patient outcomes.
Conclusion
5
The present study revealed that jaceidin significantly reduces the motility, migration and invasion of two metastatic melanoma cell lines (HMY‐1 and A2058). It has been found to be activated by phosphorylation of the ERK and JNK pathways. Additionally, jaceidin reduces the expression of ECM degradation proteins MMP2 and CTSA. Taken together, this study identifies jaceidin as a potential anti‐migratory agent that could be used clinically to improve metastatic melanoma prognosis.
Author Contributions
Ming‐Ju Hsieh: conceptualization, writing – original draft, writing – review and editing.
Funding
The authors have nothing to report.
Conflicts of Interest
The authors declare no conflicts of interest.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1R. L. Siegel , A. N. Giaquinto , and A. Jemal , “Cancer statistics, 2024,” Cancer Journal Clinicians 74, no. 1 (2024): 12–49, 10.3322/caac.21820.38230766 · doi ↗ · pubmed ↗
- 2G. De Pinto , S. Mignozzi , C. La Vecchia , F. Levi , E. Negri , and C. Santucci , “Global Trends in Cutaneous Malignant Melanoma Incidence and Mortality,” Melanoma Research 34 (2024): 265, 10.1097/CMR.0000000000000959.38391175 PMC 11045545 · doi ↗ · pubmed ↗
- 3M. Rastrelli , S. Tropea , C. R. Rossi , and M. Alaibac , “Melanoma: Epidemiology, Risk Factors, Pathogenesis, Diagnosis and Classification,” In Vivo 28, no. 6 (2014): 1005–1011.25398793 · pubmed ↗
- 4Y. Qian , P. Johannet , A. Sawyers , J. Yu , I. Osman , and J. Zhong , “The Ongoing Racial Disparities in melanoma: An Analysis of the Surveillance, Epidemiology, and End Results Database (1975–2016),” Journal of the American Academy of Dermatology 84, no. 6 (2021): 1585–1593, 10.1016/j.jaad.2020.08.097.32861710 PMC 8049091 · doi ↗ · pubmed ↗
- 5C. Stefanaki , L. Chardalias , E. Soura , A. Katsarou , and A. Stratigos , “Paediatric Melanoma,” Journal of the European Academy of Dermatology and Venereology 31, no. 10 (2017): 1604–1615, 10.1111/jdv.14299.28449284 · doi ↗ · pubmed ↗
- 6C. Garbe , T. Amaral , K. Peris , et al., “European Consensus‐Based Interdisciplinary Guideline for Melanoma. Part 1: Diagnostics: Update 2022,” European Journal of Cancer 170 (2022): 236, 10.1016/j.ejca.2022.03.008.35570085 · doi ↗ · pubmed ↗
- 7S. E. Orme and M. D. Moncrieff , “A Review of Contemporary Guidelines and Evidence for Wide Local Excision in Primary Cutaneous Melanoma Management,” Cancers (Basel) 16 (2024): 895, 10.3390/cancers 16050895.38473257 PMC 10930506 · doi ↗ · pubmed ↗
- 8J. E. Gershenwald , R. A. Scolyer , K. R. Hess , et al., “Melanoma staging: Evidence‐Based Changes in the American Joint Committee on Cancer Eighth Edition Cancer Staging Manual,” CA: a Cancer Journal for Clinicians 67, no. 6 (2017): 472–492, 10.3322/caac.21409.29028110 PMC 5978683 · doi ↗ · pubmed ↗