Absolute Quantification and Spatial Mapping of Hyaluronic Acid in Histological Tissue Sections
Cristina Quílez, Jorge González-Rico, María Luisa López-Donaire, Nuria Gago-López, Arrate Muñoz-Barrutia, Diego Velasco

TL;DR
This paper introduces a new method to measure and map hyaluronic acid levels in tissues, providing detailed spatial and quantitative insights.
Contribution
The novel integration of immunofluorescence, ELISA, and image analysis enables pixel-level hyaluronic acid quantification in tissues.
Findings
The method generates spatial maps of hyaluronic acid concentration at the pixel level.
Validation across multiple tissues shows the method is robust and scalable.
The approach is applicable to other extracellular matrix components.
Abstract
We present an accessible methodology for the absolute quantification and spatial mapping of hyaluronic acid (HA) in paraffin-embedded tissues. Although HA plays critical roles in tissue hydration, organization, and disease progression, its local concentration and spatial distribution remain poorly defined due to the lack of quantitative measurement tools. By integrating immunofluorescence, ELISA, and image analysis our approach enables the generation of pixel-level HA concentration maps. This strategy bridges the gap between qualitative imaging modalities and bulk biochemical assays, offering both spatial resolution and quantitative data. The method is validated across multiple tissues and is robust, scalable, and readily applicable to other extracellular matrix components, offering a practical tool for studying tissue microenvironments in health, disease, and biomaterial design.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
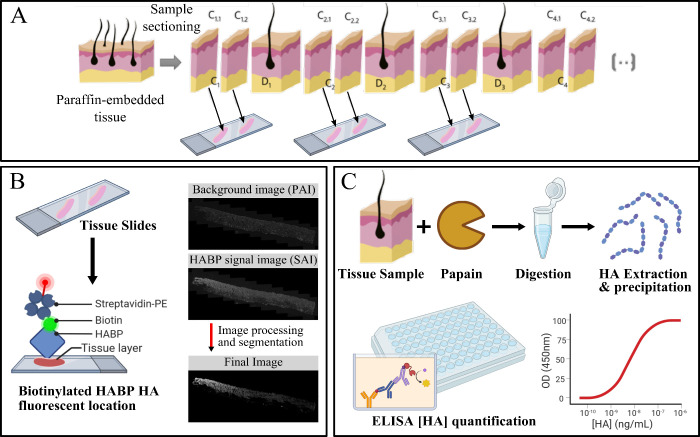 Figure 1
Figure 1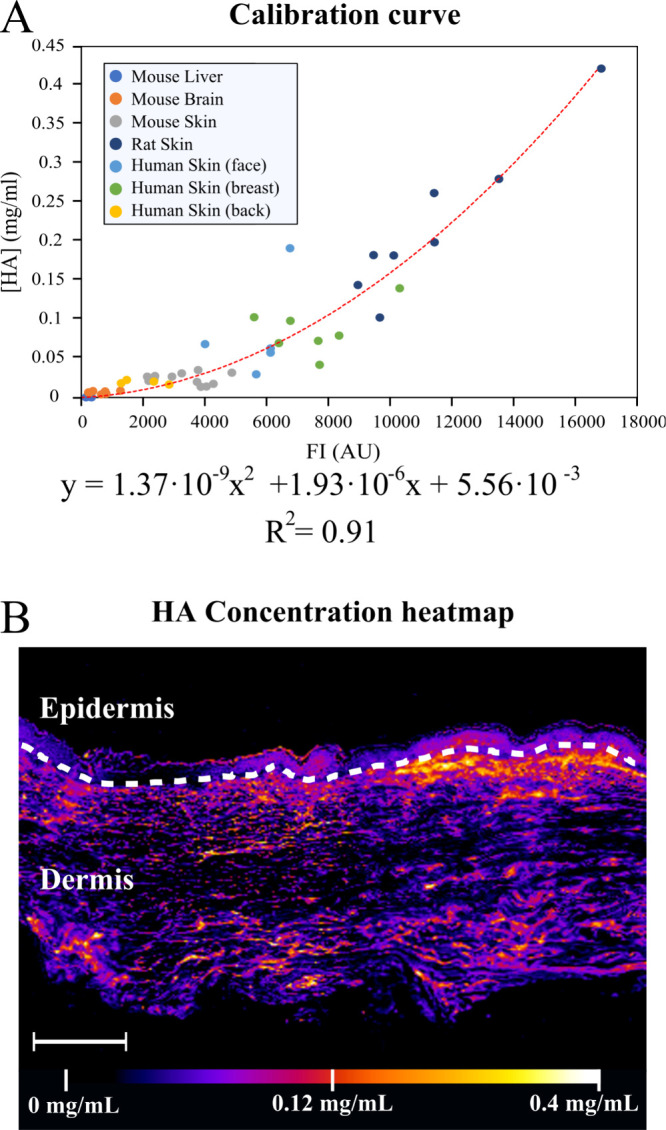 Figure 2
Figure 2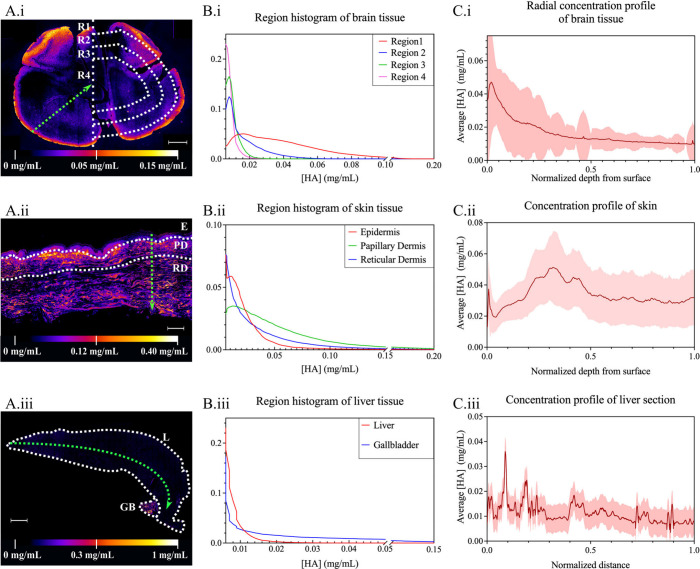 Figure 3
Figure 3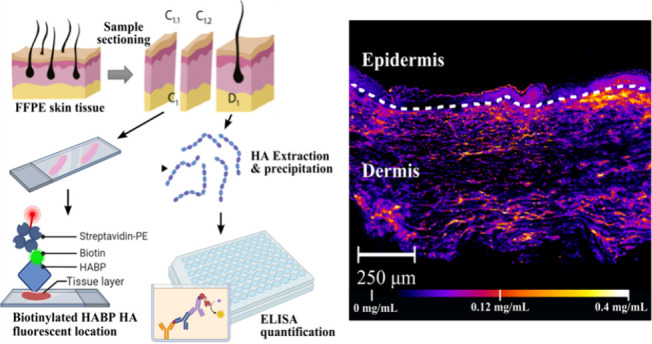 Figure 4
Figure 4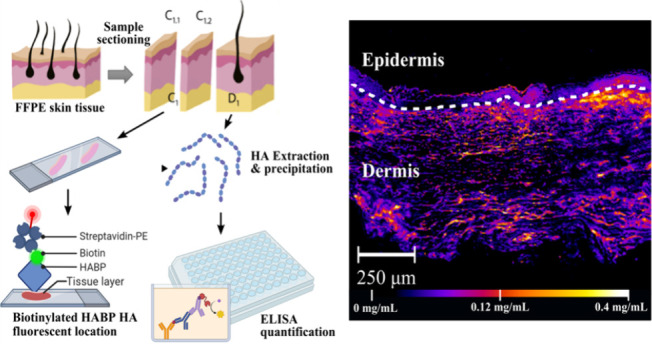 Figure 5
Figure 5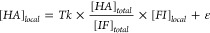 Figure 6
Figure 6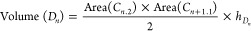 Figure 7
Figure 7- —Ministerio de Ciencia, Innovaci?n y Universidades10.13039/100014440
- —Ministerio de Ciencia, Innovaci?n y Universidades10.13039/100014440
- —Universidad Carlos III de Madrid10.13039/501100006318
- —European Regional Development Fund10.13039/501100008530
- —European Regional Development Fund10.13039/501100008530
- —Ministerio de Universidades10.13039/501100023561
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsProteoglycans and glycosaminoglycans research · Advanced Fluorescence Microscopy Techniques · Optical Imaging and Spectroscopy Techniques
The extracellular matrix (ECM) provides structural and biochemical signals that guide tissue-specific cell behaviors.? Its compositioncomprising collagens, proteoglycans, laminins, and fibronectinis uniquely structurally organized in each tissue and dynamically regulated during development, homeostasis, and disease.? A key ECM component is hyaluronic acid (HA), a nonsulfated glycosaminoglycan (GAG) abundant in soft connective tissues.? Hyaluronic acid (HA) plays key roles in hydration, tissue organization, angiogenesis, inflammation, and cell migration,? all of which are essential for both normal physiological processes and disease progression. ?,? The biological activity of HA is strongly influenced by its molecular weight and concentration,? which have been associated with cancer type and prognosis? as well as neurodevelopmental processes? and synaptic function.? Due to its multifunctional nature and ubiquitous presence, HA has also emerged as a promising biomaterial in tissue engineering, particularly for creating three-dimensional cell culture scaffolds that closely mimic the native ECM.? However, establishing HA as a complementary diagnostic marker? or effectively replicating its functions in engineered tissues requires a more comprehensive understanding of its in vivo concentration and spatial distributionparameters that remain poorly characterized. Addressing these knowledge gaps may not only clarify the role of ECM-associated HA but also facilitate its broader application as a scaffolding molecule and disease biomarker. Several approaches have previously been used to visualize or estimate HA distribution in tissues, including histochemical staining, HA-binding protein-based fluorescence, and biochemical extraction coupled with imaging readouts. ?−? ? ? ? More recently, mass spectrometry (MS)-based methods such as Matrix-Assisted Laser Desorption/Ionization (MALDI) imaging have also been applied.? Although innovative, this method is complex and limited in spatial resolution. Furthermore, the scope of the study lies uniquely on one set of ex vivo-obtained skin samples. This is due to its high complexity and specificity. While these strategies have provided valuable insights, they remain largely semiquantitative and often lack reproducibility or spatial resolution at the pixel level. In contrast, the methodology presented in this work establishes a calibration pipeline that combines HA-based fluorescence imaging with ELISA measurements from paired sections, enabling absolute quantification of HA concentration maps in paraffin-embedded tissues. This integration represents a practical advance beyond existing semiquantitative approaches, allowing reproducible, pixel-level evaluation of HA distribution across diverse tissue types. Furthermore, the method proposed in this work can be generalized to multiple sample types, taking advantage of the ubiquitous presence of HA across different tissues and species.
For this study, human, rat and mouse skin, brain, and liver biopsies of 2–3 cm^2^ were fixed using formalin-free tissue fixative (A5472, Sigma-Aldrich, USA) to prevent HA loss.? Then, tissues were embedded in paraffin (see the Supporting Information (SI)) and sectioned in a sequence of two 5 μm thickness samples (C _ n _ _.m _) used for fluorescence staining followed by a 400 μm thickness sample (D _ n _) used for tissue digestion (FigureA). These tissues were chosen to account for a broad spectrum of HA concentration as reported in the literature.? The fluorescence staining of HA within tissue sections started with the deparaffinization of the C _ n _ _.m _ samples (FigureB, SI). Next the unspecific binding sites of the tissues were blocked using 3% Phosphate-Buffered Saline (PBS) - Bovine Serum Albumin (BSA). For HA fluorescence staining, the samples were incubated first with Biotinylated Hyaluronic Acid Binding Protein (HABP-b) (AMS.HKD-BC40, AmsBio, USA), followed by secondary probe staining, with Alexa Fluor 488-Streptavidin conjugate (S11223, ThermoFisher, USA) (SI).
To accurately quantify the HA fluorescence signal, we mitigated the autofluorescence inherent to the tissue samples by acquiring images before (Primary Antibody Image (PAI)) and after (Secondary Antibody Image (SAI)) secondary antibody incubation and washing (FigureB). To prevent dehydration during image acquisition, samples were immersed in 1 × PBS. For consistency and reproducibility, fluorescence images were captured using a standardized microscope setup. Mosaic images of the whole sample were taken using a Leica Dmi8 inverted microscope with a 20× magnification dry objective with 0.4 numerical aperture using 100% laser excitation, 200 ms exposure time, and 2 × 2 camera binning. Batch effects and scanner drift were minimized by performing all imaging under fixed conditions. Instrument calibration ensured stable illumination. In addition, intensity normalization was performed to correct for potential shading. After image acquisition, the mosaic images were stitched using a 10% overlap and exported as Tagged Image Files (.tiff) for further processing. To confirm that HABP-b binding was specific to HA, tissues were treated with hyaluronidase (HAase; H1136, Sigma-Aldrich, USA) at pH 7 for 5 h at 37 °C, following the manufacturer’s protocol. Additionally, to further prove its specificity, competitive inhibition of the HABP-b with soluble HA was performed, according to previously published protocols.? Briefly, soluble HA (15MDa, NC1584217, Lifecore Biomedical, USA) at a final concentration of 0.05 mg/mL in the presence of HABP-b (at final concentration of 2.5 μg/mL) for 3 h at RT with gentle shaking before incubation in the tissue sample.
GAG extraction from skin biopsies was performed by digestion of D _ n _ sections (FigureC) with papain enzyme following the protocol by van Wijk et al.? with modifications (SI). The GAGs precipitate was resuspended in NaCl 0.9% (w/v) to quantify the HA concentration using an ELISA kit (K-4800, Echelon Biosciences, USA).
Finally, the direct relationship between the total HA of a tissue section to the fluorescence signal intensity (FI) was represented with the following eq (eq):
where [FI]_ total _ represents the total fluorescence intensity of a C _ n _ ,i _ skin section, [HA] total _ is total HA concentration of a D _ n _ skin section (in mg/mL), [FI]_ local _ and [HA]_ local _ are the fluorescence intensity and amount of HA in each pixel of a C _ n _ _,i _ skin section (in arbitrary units (A.U.) and mg/mL, respectively), Tk is a thickness normalization term, and ε reflects the residuals between predicted and observed HA values from the calibration curve (SI).
To calculate the total fluorescence intensity ([FI]_ total _), image analysis was carried out with each of the two images obtained for each C _ n _ .m _ section. The autofluorescence captured after primary antibody incubation (PAI) was subtracted from the image acquired after secondary antibody (SAI) staining. Prior to subtraction, the images were registered using the ImageJ plugin “Landmark-based Correlation” to ensure pixel-wise alignment. The subtraction was then performed using the “Calculator Plus” tool in ImageJ. The resulting image contained only the fluorescence signal attributable to HA. To correct for background signal, the average intensity of nontissue regions (background pixels) was subtracted from the entire image. The total HA-related fluorescence intensity ([FI] total _) was then computed by summing all pixel intensities within the segmented tissue area. This value was normalized by the sample volume to yield the FI concentration for the section. The entire workflow is summarized schematically in Figure S1.
For each C _ n.m _ section, the tissue volume was estimated as the product of the segmented area (in pixels) and the section thickness. Otsu thresholding was used to segment the tissue area from the background in the fluorescence images. The resulting volume estimate was used to normalize the total fluorescence intensity, yielding the IF concentration. The final unit for this value is expressed in arbitrary fluorescence units per pixel (AU/px). To generate accurate pixel-wise concentration maps across samples of varying thickness, a normalization factor Tk (SI) was applied to account for differences in section thickness. For each D _ n _ section (used for biochemical quantification), volume was calculated assuming the shape of the section is approximately continuous and free of abrupt morphological changes. Thus, the volume (eq) was estimated using the average area of the two adjacent thin sections C _ n. _ 2 and C _ n. _ +1,1, multiplied by the known thickness (h _ D _ n _ _), the thick section (FigureA):
By correlating the concentration values obtained from the total fluorescence intensity measured in C _ n.m _ through imaging (FI) and biochemical quantification (via ELISA in D _ n _), a calibration curve was constructed to convert pixel-level fluorescence intensity into absolute HA concentration (FigureA). This relationship yielded a robust polynomial regression fit (R^2^ = 0.91), allowing the conversion of fluorescence intensity at each pixel into absolute HA concentration values. Residual analysis demonstrated that the second-order polynomial regression provided a more adequate fit to the data than the linear model (Figure S2). Using this calibration, quantitative spatial maps of HA distribution were generated for tissue sections (FigureB). To validate the specificity of the staining, HAase digestion experiments confirmed that the HA signal is selectively lost upon enzymatic degradation of HA (Figure S3A–D). Additionally, sections incubated only with the secondary antibody or with competitive inhibition of the HABP-b with HABP-b confirmed the absence of nonspecific binding to endogenous biotin or other ECM components (Figure S3E,F)
This methodology was applied to assess HA distribution across tissues with known HA presence. In mouse brain sections, HA concentration displayed marked spatial heterogeneity (FigureA(i)). To facilitate analysis, arbitrary radial regions were defined, and pixel-level HA concentrations were represented as density plots (FigureB(i)). These reveal that HA levels were highest near the cortex, HA concentration maps revealed a decreasing gradient from the cortical surface inward (FigureC(i)). This observation is consistent with enrichment of HA in perineuronal nets and other extracellular specializations (9,10,22). Similarly, human (FigureA(ii)) and rat skin (Figure S4) sections show a heterogeneous distribution of HA. Manual segmentation into epidermis, papillary dermis, and reticular dermis reveal a clear separation between the epidermis and deeper layers, with the reticular dermis consistently exhibiting higher HA levels (FigureB(ii)). ?,? HA concentration declines gradually with depth (FigureC(ii)). While the relative spatial pattern is consistent, absolute values varied across anatomical sites (Figure S5) and individuals (Figure S6). In contrast, mouse liver sections exhibit a uniform absence of HA (FigureA(iii)), whereas the adjacent gallbladder showed prominent HA accumulation (FigureB(iii), FigureC(iii)).?
Finally, to assess reproducibility, both intraslide and interslide variability were quantified. Technical replicates (duplicate sections from the same block) consistently showed intraslide coefficients of variation (CVs) of approximately 25%, while interslide biological replicates (different individuals per group) yielded CVs in the range of 44.00 ± 17.20% (95% CI) Table S1. Intraclass correlation coefficients (ICCs) further confirmed good reproducibility (ICC = 0.904 ± 0.041% (95% CI)) within samples Table S2.
By enabling absolute quantification, this approach permits direct comparison of HA concentrations across tissues, individuals, and experimental conditions. This capability extends beyond qualitative or relative analyses, facilitating reproducible benchmarking of HA abundance in both physiological and pathological contexts. In skin, HA exhibits a differential distribution aligned with well-defined anatomical structures. The higher HA levels observed in the reticular dermis compared to the epidermis are consistent with the established distribution of HA within the extracellular matrix. ?,? In contrast, the organization of brain regions is considerably more complex.? In this study, brain data are presented using a simplified spatial representation to validate the methodology. Nonetheless, this approach can be applied in future investigations specifically designed to examine the biological relevance of HA distribution across distinct brain areas. Such studies, incorporating anatomically guided segmentation and targeted experimental designs, will enable a more detailed assessment of region-specific HA organization and its functional implications. Regarding the distribution and organization of HA in mouse liver sections, the sharp contrast in HA concentration between liver and gallbladder regions is noteworthy. The biological relevance of this difference likely relates to the viscoelastic and hydrating properties of HA, which support mechanical resilience, lubrication, and volume regulation. Such characteristics are consistent with the gallbladder’s physiological role as a distensible reservoir for bile storage and release.? The ability to quantify these tissue-specific patterns highlights the potential of absolute HA mapping to connect extracellular matrix composition with organ function.
The findings from this study provide a framework for determining absolute HA concentrations at defined spatial locations within diverse tissue types. This capability is critical for several applications: (i) the development of physiologically relevant in vitro models that better mimic native tissue microenvironments; (ii) a more precise understanding of the role of HA in disease progression, including its associations with cancer prognosis and treatment outcomes; and (iii) the potential use of HA distribution as a complementary diagnostic or prognostic biomarker in various pathologies. Notably, the method has been optimized for HA, a particularly challenging molecule to quantify due to its nonimmunogenic and nonproteinaceous nature, as it is a polysaccharide rather than a protein and lacks stable epitopes for conventional immunochemical detection. In addition, HA is highly dynamic and susceptible to endogenous enzymatic and oxidative degradation, ?−? ? a factor especially relevant in our study since the analyzed tissues were obtained from deceased donors. Precise post-mortem intervals and fixation times were not available for human donor tissues, and therefore, some degree of HA degradation prior to fixation cannot be excluded. This factor is especially relevant given the short half-life of HA in skin and other tissues. ?−? ? Nevertheless, because the calibration curve is derived from paired sections of the same tissue block, any degradation occurring prior to fixation is internally accounted for, ensuring robust relative quantification and reliable spatial mapping. Furthermore, several methodological limitations should be acknowledged. First, our workflow was optimized for paraffin-embedded tissues, which ensures broad applicability in pathology and provides superior structural preservation compared with frozen sections.? While HABP-based staining is also compatible with frozen samples, systematic benchmarking of the calibration workflow in that context remains to be performed. In this regard, fixation parameters such as temperature, blocking conditions, and incubation times must be carefully controlled to prevent HA degradation,? and complete paraffin removal is essential to ensure optimal accessibility for HABP binding. ?,? No antigen-retrieval steps are required, as HA is a polysaccharide rather than a protein.? Second, HABP affinity may vary with HA chain length or chemical modifications, ?,? potentially affecting staining intensity. Third, the present method provides absolute concentration maps but does not capture HA molecular weight distribution, which is critical for bioactivity. ?,? Future refinement will involve validating frozen tissue compatibility, cross-validating HABP binding across HA size variants, and integrating molecular weight–sensitive analytical assays. While this study focused on healthy tissues to establish and validate the workflow, applying the method to pathological contexts in future workssuch as tumor microenvironments or fibrotic tissueswill help reveal biologically relevant differences in HA distribution. Combining molecular weight–sensitive techniques with analyses of annotated human and animal cohorts will allow systematic assessment of how donor factors and ECM remodeling (e.g., fibrosis or edema) influence HA spatial organization and quantitative mapping. ?,?
To place these findings in context, our results should be interpreted in light of previous efforts to visualize HA in tissues, many of which relied on histochemical, biochemical, or semiquantitative fluorescence methods. ?−? ?,? While these approaches provided valuable qualitative insights, they were limited by relative measurements, restricted reproducibility, and a lack of pixel-level resolution. By integrating HABP-based fluorescence imaging with ELISA calibration from paired sections, the methodology presented here offers an accessible workflow for absolute HA quantification in standard paraffin-embedded samples. This advance enables reproducible comparisons of HA concentrations across tissues, individuals, and experimental conditions.
Recent developments in extracellular matrix (ECM) imaging and quantificationsuch as second harmonic generation (SHG) microscopy for collagen and MS imaging for proteoglycanshave substantially advanced our understanding of ECM organization and dynamics. ?−? ? ? However, these techniques fail to provide both absolute quantitative and highly spatially resolved information across a broad range of ECM components, particularly glycosaminoglycans such as HA, whose nonprotein, highly hydrated nature makes them difficult to detect and quantify with label-free or MS-based modalities. Despite inherent challenges in HA detection, our HABP-based workflow enables absolute, spatially resolved quantification of HA, yielding robust quantitative data that can be readily integrated with measurements of other ECM components, including collagen I, fibronectin, or elastin. Importantly, the methodology relies only on standard laboratory equipmentsuch as a fluorescence microscopeand does not require specialized instrumentation, making the workflow broadly accessible and scalable for histological studies. Overall, this pipeline provides a practical and reproducible tool for in situ spatial quantification of ECM components in paraffin-embedded tissues, extending the ECM imaging repertoire and supporting comprehensive analyses of tissue organization, remodeling, and function.
Supplementary Material
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Mouw J. K.Ou G.Weaver V. M.Extracellular matrix assembly: A multiscale deconstruction Nature Reviews Molecular Cell Biology 20141577178510.1038/nrm 390225370693 PMC 4682873 · doi ↗ · pubmed ↗
- 2Hu M.Ling Z.Ren X.Extracellular matrix dynamics: tracking in biological systems and their implications J. Biol. Eng.202216111310.1186/s 13036-022-00292-x 35637526 PMC 9153193 · doi ↗ · pubmed ↗
- 3Fraser J. R. E.Laurent T. C.Laurent U. B. G.Hyaluronan: Its nature, distribution, functions and turnover J. Intern Med.19972421273310.1046/j.1365-2796.1997.00170.x 9260563 · doi ↗ · pubmed ↗
- 4Salwowska N. M.Bebenek K. A.Żądło D. A.Wcisło-Dziadecka D. L.Physiochemical properties and application of hyaluronic acid: a systematic review J. Cosmet Dermatol.2016154520610.1111/jocd.1223727324942 · doi ↗ · pubmed ↗
- 5Gupta R. C.Lall R.Srivastava A.Sinha A.Hyaluronic Acid: Molecular Mechanisms and Therapeutic Trajectory [Internet]Frontiers in Veterinary Science.20196110.3389/fvets.2019.0019231294035 PMC 6603175 · doi ↗ · pubmed ↗
- 6Knudson W.Tumor-associated hyaluronan. Providing an extracellular matrix that facilitates invasion Am. J. Pathol.1996148617218669457 PMC 1861627 · pubmed ↗
- 7Karjalainen J. M.Tammi R. H.Tammi M. I.Eskelinen M. J.Agren U. M.Parkkinen J. J.Reduced level of CD 44 and hyaluronan associated with unfavorable prognosis in clinical stage I cutaneous melanoma Am. J. Pathol.200015739576510.1016/S 0002-9440(10)64608-110980134 PMC 1885706 · doi ↗ · pubmed ↗
- 8Rosales P.Vitale D.Icardi A.Sevic I.Alaniz L.Role of Hyaluronic acid and its chemical derivatives in immunity during homeostasis, cancer and tissue regeneration Semin Immunopathol.20244651510.1007/s 00281-024-01024-739240397 · doi ↗ · pubmed ↗