Possible regulation of Arabidopsis MYB93 by both SCARECROW and MPK3
Xulyu Cao, Clare Clayton, Bethany Hutton, Nancy McMulkin, Juliet C. Coates

TL;DR
This study explores how the Arabidopsis MYB93 gene is regulated by two proteins, SCARECROW and MPK3, through experiments and genetic analysis.
Contribution
The paper provides experimental evidence that MYB93 is regulated by SCARECROW and interacts with MPK3, linking these factors to plant development.
Findings
MYB93 transcript levels are elevated in the scr-3 mutant and reduced with SCR-GFP expression.
The N-terminal domain of MYB93 interacts with MPK3 in yeast.
mpk3-1 mutants show a lateral root phenotype similar to myb93-1 mutants.
Abstract
The promoter of the Arabidopsis MYB93 ( MYB93 ) transcription factor was previously identified in a large-scale screen using the SCARECROW ( SCR ) transcription factor. Independent high-throughput studies also identified MYB93 as a protein-interaction partner of the MAP kinase 3 ( MPK3 ). Here, we validate and extend those observations using RT-PCR, yeast two-hybrid assays and phenotypic analysis. MYB93 transcript levels were elevated in the scr-3 mutant and reduced upon expression of SCR-GFP, indicating regulation by SCR. In yeast, the N-terminal domain of MYB93 , but not the C-terminal region, interacted with MPK3 . We also observed that mpk3-1 mutants exhibited a lateral root phenotype similar to myb93-1 . Together, these findings support a model in which both MYB93 gene expression and MYB93 protein function are modulated by SCR and MPK3 , respectively.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
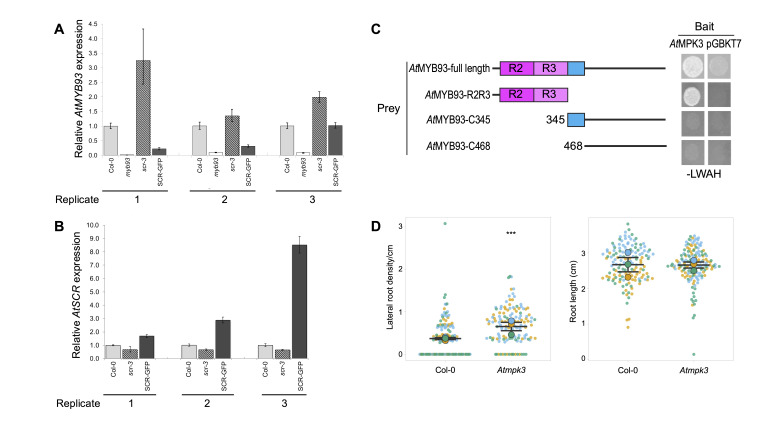 Figure 1
Figure 1|
|
|
|
|
|
AAGCTCGCAGATTTGAATAGGTG |
ATCTGTACGACCTTGCAAATGC |
|
|
GCAGATAAGCTTGGCCTGCC |
GGAGCTAATCTTTGGAGTAACCAG |
|
|
TCGTACAACCGGTATTGTGCTG |
TAACAATTTCCCGCTCTGCTG |
|
|
CGATTCTTGACCAAGATATTCCATC |
TTAGAAGATTCCCTGAGTCGCAG |
|
|
|
|
|
|
AAAGAATTCGGGAGGTCGCCTTGTTGC |
AAAGGATCCCTAAGATATAACGTTCATGAGG |
|
|
AAAGAATTCGGGAGGTCGCCTTGTTGC |
AAAGGATCCTTTCTTCTTTAGATGTGTGTTCC |
|
|
AAAGAATTCTTGATCCAGATGGGGATCG |
AAAGGATCCCTAAGATATAACGTTCATGAGG |
|
|
AAAGAATTCTCCATGCAAGGCGAAGCAG |
AAAGGATCCCTAAGATATAACGTTCATGAGG |
|
|
AAAGAATTCAACACCGGCGGTGGCC |
AAAGGATCCCTAACCGTATGTTGGATTGAGTGC |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Gene Expression Analysis · Plant-Microbe Interactions and Immunity · Polysaccharides and Plant Cell Walls
Description
The Arabidopsis MYB93 ( At1g34670 ) transcription factor is a member of the plant R2R3-MYB transcription factor family (Du et al., 2015) whose gene expression is largely restricted to root endodermal cells overlying developing lateral root primordia and is transiently upregulated during the early stages of lateral root development (Gibbs et al., 2014; Shukla et al., 2021; Voss et al., 2015). MYB93 is a negative regulator of lateral root development as myb93 mutants show increased lateral root density whilst MYB93-overexpressing plants have fewer lateral roots (Gibbs et al., 2014). MYB93 is part of a small clade of three related proteins, the S24 clade, alongside MYB92 ( At5g10280 ) and MYB53 ( At5g65230 ) (Du et al., 2015; Gibbs et al., 2014). MYB93 , MYB93 and MYB53 , alongside MYB41 ( At4g28110 ), function redundantly to regulate suberin biosynthesis in the root endodermis (Shukla et al., 2021). However, the three S24 genes do not appear to function completely redundantly during lateral root development as myb93 , but not myb92 , mutants show elevated lateral root density (Gibbs et al., 2014). In addition, only * MYB93 * expression is induced by auxin (Gibbs et al., 2014). Furthermore, via enhanced yeast one hybrid analysis, * MYB93 * (but not * MYB92 * or * MYB53 * ) is implicated in a root signaling network downstream of the endodermal cell identity gene SCARECROW ( * SCR * , At3g54220 ) (Iyer-Pascuzzi et al., 2011; Sparks et al., 2016). Finally, publicly available protein interaction data (Oughtred et al., 2019; Popescu et al., 2009; Wanamaker et al., 2017) suggests that MYB93 has a unique set of interaction partners compared to other S24 clade members. High-throughput protein microarray analysis (Popescu et al., 2009) shows that MYB93 interacts with MITOGEN ACTIVATED PROTEIN KINASE 3, ( MPK3 , At3g45640 ) while other S24 proteins do not. In contrast, MPK6 ( At2g43790 ) interacts with MYB93 , MYB92 and MYB41 (Hoang et al., 2012; Popescu et al., 2009).
To further explore the upstream regulation of MYB93 , we firstly analyzed the expression of * MYB93 * in the
- scr-3* mutant using qRT-PCR. We showed that in three separate biological repeats with similar trends,
MYB93 * levels are upregulated in scr-3 mutant seedlings and reduced in a ‘rescue' line ( * p SCR ::GFP- SCR * (Goh et al., 2016)) where * SCR * is re-introduced into a scr-3 background under the control of its own promoter ( Figure 1A, 1B). This demonstrates that SCR is a likely negative regulator of * MYB93 * expression, extending previous protein-DNA interaction studies (Iyer-Pascuzzi et al., 2011; Sparks et al., 2016). Given the highly restricted localization of * MYB93 * promoter activity (Gibbs et al., 2014), our data suggest that SCR inhibits * MYB93 * promoter activity in most endodermal cells in the root. Interestingly, in a time course transcriptome of root segments induced to form lateral root primordia, * SCR * gene expression is significantly downregulated at 9-12h, immediately before the largest upregulation of * MYB93 * at 12-15h (Voss et al., 2015), suggesting that localized * SCR * downregulation may be required for * MYB93 * induction.
To further investigate MPK3 as a potential protein regulator of MYB93 , we tested the interaction of MYB93 with MPK3 in the yeast two-hybrid system. We showed that full-length MYB93 interacts with MPK3 ( Figure 1C ). Furthermore, truncation of MYB93 showed that the N-terminus of the protein (amino acids 2-365, encompassing the R2R3-MYB domain but not the downstream unique motif (Gibbs et al., 2014)) was necessary and sufficient to mediate interaction with MPK3 ( Figure 1C ). The N-terminal region of MYB93 interacted with MPK3 in yeast but the C-terminal region of MYB93 (amino acids 115-365 or 156-365) did not ( Figure 1C ). We next investigated the impact of loss of MPK3 on root development and showed that the mpk3-1 mutant has elevated lateral root density compared to wild type, similarly to Atmyb93 ( Figure 1D ; (Gibbs et al., 2014)) but no difference in primary root length ( Figure 1D ). This would position MPK3 as a potential positive regulator of MYB93 during lateral root development, similar to the positive regulation of MYB44 ( At5g67300 ) by MPK3 and MPK6 (Nguyen et al., 2012). A previous study suggested that an mpk3 mutant does not show a lateral root phenotype (Zhu et al., 2019), although this study was performed on slightly older seedlings (10 days vs 8 days), meaning that differences occurring during early lateral root development may have been overlooked in the 10-day old seedlings. A conditional *mpk3/mpk6 * double mutant has fewer emerged lateral roots, suggesting that * MPK3 * and * MPK6 * could together promote lateral root emergence, via the auxin transporter * LAX3 * (Zhu et al., 2019). As * MYB93 * inhibits lateral root initiation as well as emergence (Gibbs et al., 2014), this suggests that * MPK3 * , on its own or together with * MPK6 * , may play contrasting roles at different stages of lateral root development. Collectively, our data suggest that MPK3 may work with MYB93 during lateral root development but not during primary root development.
In summary, we have extended previous data to show that * MYB93 * gene expression is negatively regulated by the endodermal transcription factor SCARECROW . We have also demonstrated that the R2R3-MYB domain of MYB93 interacts with the MAP kinase MPK3 . Furthermore, we have implicated MPK3 in the negative regulation of lateral root development, as a potential positive regulator of MYB93 . Thus, we provide new insights into the mechanism by which endodermal MYB93 regulates lateral root development in a very specific and localized manner.
Methods
RNA extraction and cDNA generation
Up to 100mg plant tissue from pooled 7-day old seedlings was ground in liquid nitrogen using RNAse-free ceramic pestles and mortars. RNA was extracted using an ISOLATE II Plant RNA kit (Bioline, Meridian Biosciences, Memphis, TN, USA). cDNA was generated from RNA using the SuperScript ^TM^ III first-strand synthesis system (Invitrogen, ThermoFisher Scientific, Waltham, MA, USA).
Quantitative RT-PCR (qRT-PCR).
qRT-PCR was carried out using Brilliant III ultra-fast SYBR Green low ROX qPCR master mix (600892, Aglient Technologies, Stockport, UK) using a final template concentration of 1ng/µl based on the concentration of RNA added to the cDNA synthesis reaction. The primers used were as follows.
**: **
Primers were used at a final concentration of 200nM-400nM depending on primer efficiency. Reactions were carried out on an AriaMx qPCR machine (Agilent Technologies, Stockport, UK) was used with cycling parameters of 95°C for 10 min, 40 amplification cycles of 95°C for 30 s and 60°C for 1 min. After this, a melt curve cycle (95°C 30 s) was performed, then 65°C for 30 s and ramping back to 95°C for 30 s at a ramp rate of 0.3°C every 2 s to produce a dissociation curve. Three technical repeats per plate were carried out for each sample and three biological replicates were performed overall. Cq values were normalised to *ACTIN2 * ( At3g18780 ) and * UBC21 * ( At5g25760 ) housekeeping controls and fold changes calculated using the ΔΔCt method (Livak & Schmittgen, 2001). Statistics were performed on ΔCt values of combined replicates: ANOVA followed by a Dunnet's post-hoc test.
Yeast two-hybrid assays.
MYB93 * ( At1g34670 ) full-length and three truncated cDNAs (nucleotides 1-344, 345-1378, 468-1378) were cloned into the pGADT7 vector; full-length * MPK3 * (A3g45640) cDNA was cloned into the pGBKT7 vector. The primers used had appropriate restriction sites added for cloning and the primer pairs used were as follows.
**: **
2µg of each plasmid (constructs or empty vector controls) was transformed into S. cerevisiae strain AH109 Hansen following a small-scale transformation protocol ((Clontech, 2009); Takara Biosciences, Otsu, Japan). Transformed yeast was grown on drop out medium (SD) -LW (DSCK172, Formedium, Swaffham, UK) agar plates for 2-3 days at 30˚C until transformants were observed. Individual transformed colonies were selected inoculated into 50µl of sterile distilled water, 5µl of which was then pipetted onto SD -AHLW (DSCK272 Formedium, Swaffham, UK) agar plates for 2-3 days at 30˚C to test for protein-protein interaction.
Plant genotypes, growth conditions and lateral root assays.
Arabidopsis ecotype Col-0 wild type and the myb93-1 mutant (SALK_131752, NASC ID N631752; (Gibbs et al., 2014)), mpk3-1 mutant (SALK_151594, NASC ID N869692; (Merkouropoulos et al., 2008)), scr-3 (Gallagher et al., 2004) and * p SCR ::GFP- SCR * in scr-3 (Goh et al., 2016) were grown in Levington M3 compost/vermiculite mix at 22˚C under 16h light in a glasshouse. For qPCR, root assays and magenta pot growth, seeds were sterilised for 10 minutes in 10% Parozone ^TM^ Bleach (Jeyes, Hemel Hempstead, UK) followed by 3 rinses in sterile distilled water and resuspension in 200µl distilled water. Seeds were vernalized in the dark at 4˚C for 2 days. For root assays and qRT-PCR, seeds were plated in rows at the top of half-strength Murashige and Skoog (MS) medium (M0404, Sigma-Aldrich, St Louis, Missouri, USA) pH5.6-5.8 with 1% agar. Seedlings were grown vertically for 7 days (qRT-PCR) or 8 days (root assays) and 8-day root plates were photographed. Emerged lateral roots and adventitious roots (roots emerging from the collet) were counted by eye from plates and root length was measured from photographs using the freehand drawing tool in ImageJ (https://imagej.net/ij/). Lateral root density was calculated for each seedling by dividing lateral root number by primary root length. Root data was visualised using SuperPlots ((Lord et al., 2020); https://huygens.science.uva.nl/SuperPlotsOfData/).
Statistical significance for lateral root density and primary root length between Col-0 and mpk3 was calculated using a pairwise Mann-Whitney U-test.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Du Hai Liang Zhe Zhao Sen Nan Ming-Ge Tran Lam-Son Phan Lu Kun Huang Yu-Bi Li Jia-Na 201565 The Evolutionary History of R 2R 3-MYB Proteins Across 50 Eukaryotes: New Insights Into Subfamily Classification and Expansion Scientific Reports 512045-232210.1038/srep 11037 PMC 460378426047035 · doi ↗ · pubmed ↗
- 2Gallagher Kimberly L. Paquette Alice J. Nakajima Keiji Benfey Philip N. 2004101 Mechanisms Regulating SHORT-ROOT Intercellular Movement Current Biology 14200960-98221847185110.1016/j.cub.2004.09.08115498493 · doi ↗ · pubmed ↗
- 3Gibbs Daniel J. VoßUte Harding Susan A. Fannon Jessica Moody Laura A. Yamada Erika Swarup Kamal Nibau Candida Bassel George W. Choudhary Anushree Lavenus Julien Bradshaw Susan J. Stekel Dov J. Bennett Malcolm J. Coates Juliet C. 201466 At MYB 93 is a novel negative regulator of lateral root development in Arabidopsis New Phytologist 20340028-646X 1194120710.1111/nph.1287924902892 PMC 4286813 · doi ↗ · pubmed ↗
- 4Goh Tatsuaki Toyokura Koichi Wells Darren M. Swarup Kamal Yamamoto Mayuko Mimura Tetsuro Weijers Dolf Fukaki Hidehiro Laplaze Laurent Bennett Malcolm J. Guyomarc'h Soazig 201611 Quiescent center initiation in the Arabidopsis lateral root primordia is dependent on the SCARECROW transcription factor Development 1477-912910.1242/dev.13531927510971 · doi ↗ · pubmed ↗
- 5Hoang My Hanh Thi Nguyen Xuan Canh Lee Kyunghee Kwon Young Sang Pham Huyen Trang Thi Park Hyeong Cheol Yun Dae-Jin Lim Chae Oh Chung Woo Sik 201251 Phosphorylation by At MPK 6 is required for the biological function of At MYB 41 in Arabidopsis Biochemical and Biophysical Research Communications 42210006-291X 18118610.1016/j.bbrc.2012.04.13722575450 · doi ↗ · pubmed ↗
- 6Iyer-Pascuzzi Anjali S. Jackson Terry Cui Hongchang Petricka Jalean J. Busch Wolfgang Tsukagoshi Hironaka Benfey Philip N. 2011101 Cell Identity Regulators Link Development and Stress Responses in the Arabidopsis Root Developmental Cell 2141534-580777078210.1016/j.devcel.2011.09.00922014526 PMC 3204215 · doi ↗ · pubmed ↗
- 7Livak Kenneth J. Schmittgen Thomas D. 2001121 Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method Methods 2541046-202340240810.1006/meth.2001.126211846609 · doi ↗ · pubmed ↗
- 8Lord Samuel J. Velle Katrina B. Mullins R. Dyche Fritz-Laylin Lillian K. 2020428 Super Plots: Communicating reproducibility and variability in cell biology Journal of Cell Biology 21960021-952510.1083/jcb.202001064 PMC 726531932346721 · doi ↗ · pubmed ↗