VviAMT4;1 Is a High-Affinity Ammonium Transporter in Table Grape
Huilin Xiao, Matthew Shi, Yanwen Tang, Rui Yuan, Zhizhong Song, Meiling Tang

TL;DR
This study identifies a high-affinity ammonium transporter in table grape roots, which helps in understanding nitrogen uptake in fruit trees.
Contribution
The novel contribution is the identification and characterization of VviAMT4;1 as a high-affinity ammonium transporter in table grape.
Findings
VviAMT4;1 is highly expressed in grape roots and is upregulated under ammonium depletion.
VviAMT4;1 has a Kₘ of 49.58 μmol·L−1 and Vₘₐₓ of 3.29 μmoles·min−1·μg−1 cells, indicating high-affinity ammonium transport.
The transporter also mediates weak methyl amine uptake and is inhibited by endogenous ammonium.
Abstract
Ammonium transporters (AMTs) are a class of membrane-associated proteins that play crucial roles in the uptake and transport of ammonium (NH4+ or NH3). In this study, an ammonium transporter-encoding gene VviAMT4;1 was isolated and identified from table grape ‘Yanpu No.2’. Notably, the expression level of VviAMT4;1 varied significantly across different organs or tissues of ‘Yanpu No.2’, and the highest expression level was detected in the roots of both tissue-cultured seedlings and 5-year-old mature trees. Expression of VviAMT4;1 was significantly up-regulated under NH4+ depletion throughout the whole of tissue-cultured seedlings. Yeast mutant functional complementation indicates that the recombinant strain pYES2-VviAMT4;1/31019b restored growth under different pH conditions. 15N isotope-labeled uptake kinetics analysis demonstrated that VviAMT4;1 is a typical high-affinity ammonium…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1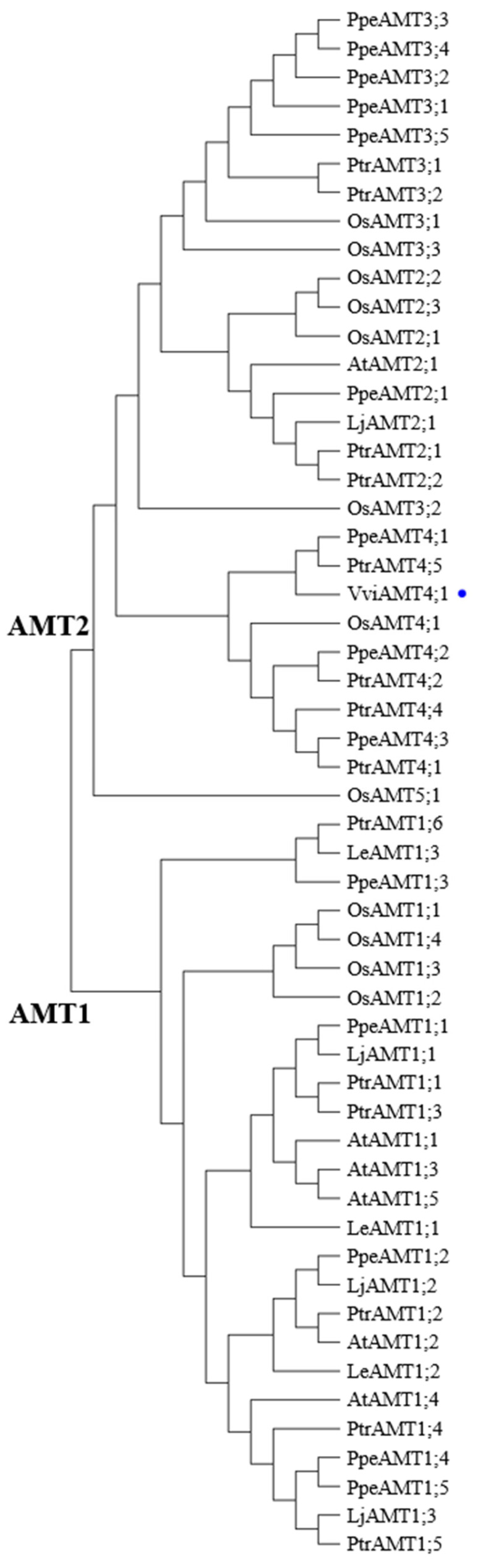 Figure 2
Figure 2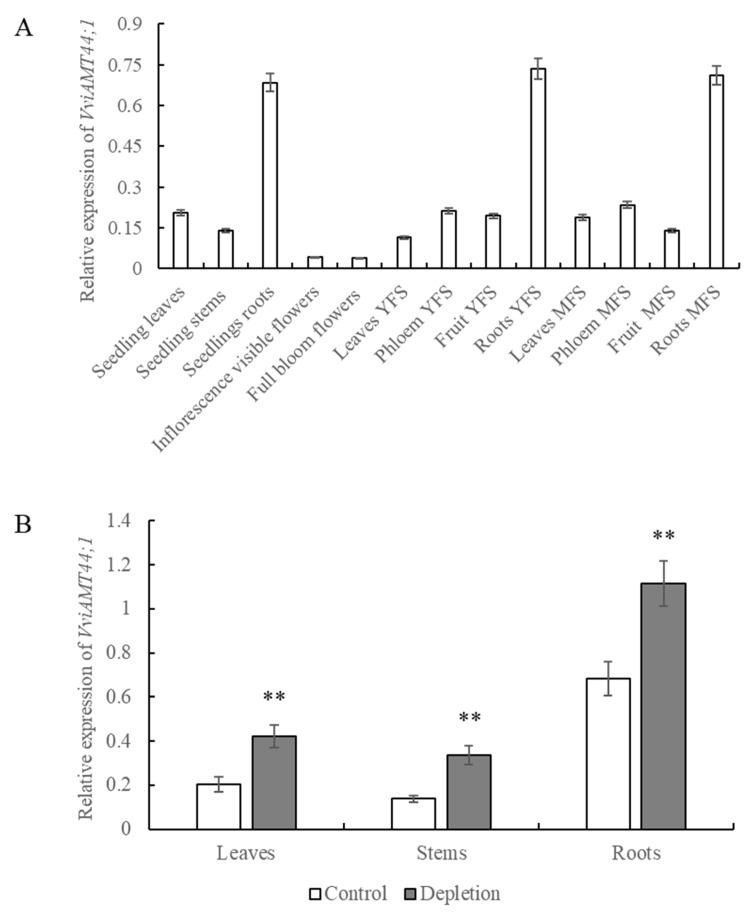 Figure 3
Figure 3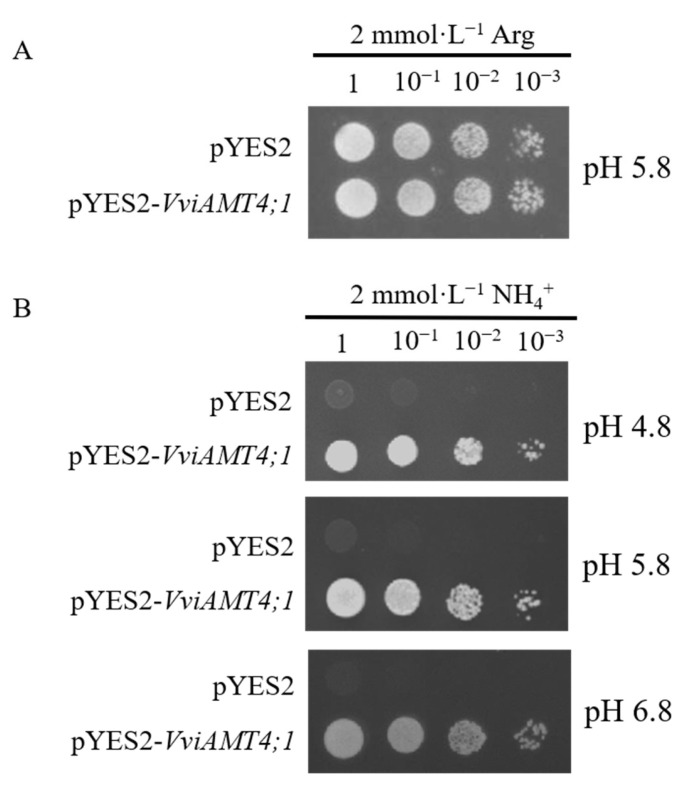 Figure 4
Figure 4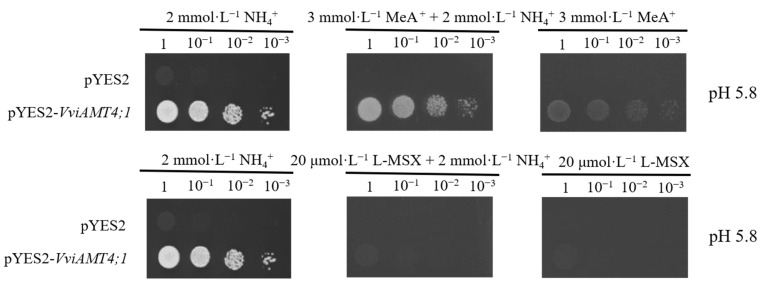 Figure 5
Figure 5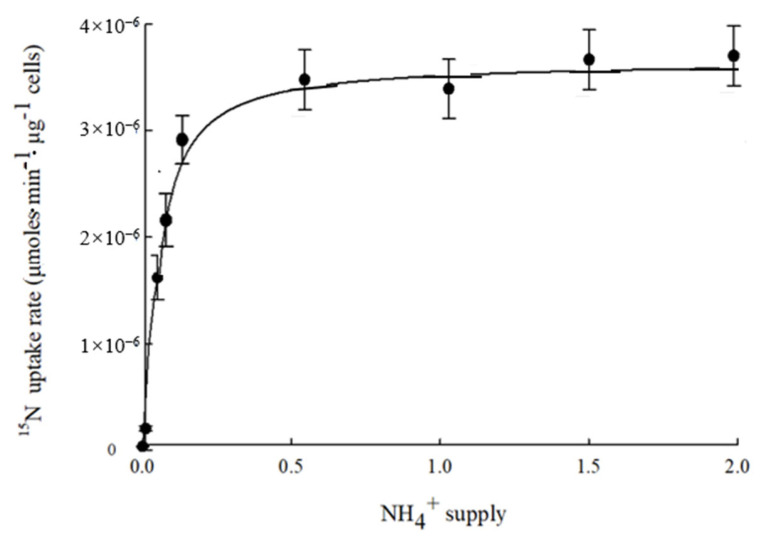 Figure 6
Figure 6- —Key R&D Program of Shandong Province—Innovation Capability Enhancement Project for Technology-based Small- and Medium-sized Enterprises
- —Major Project of Science and Technology of Shandong Province
- —China Agriculture Research System of MOF and MARA
- —China Scholarship Council Fund
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant nutrient uptake and metabolism · Ion Transport and Channel Regulation · Plant responses to water stress
1. Introduction
Ammonium serves as a primary nitrogen source for plants and exhibits significant absorption advantages under low-nitrogen conditions (soil NH_4_^+^ concentration < 1 mmol·L^−1^), with its transmembrane transport efficiency exceeding that of nitrate by 3–5 times. However, when the intracellular ammonium concentration exceeds a critical threshold (typically >5 mmol·L^−1^), it triggers harmful effects such as proton imbalance and membrane potential disorder [1,2]. The precise regulation of this “dual effect” relies on the functional synergy of the ammonium transporter (AMT) family [3,4,5].
In plants, AMT transporters are mainly located on the plasma membrane, containing 10 or 11 typical conserved transmembrane regions, and transport ammonium via a high- or low-affinity transport system [3,4,5,6]. Plant AMTs are divided into two major sub-groups, AMT1 and AMT2, which share similar topological structures and mediate ammonium absorption and transport functions in the form of trimers [3,5]. Since the cloning of AtAMT1;1 in Arabidopsis thaliana [7], dozens of AMT1 subgroup homologous genes have been identified in annual plants such as tomato (Solanum lycopersicum) [8,9], rice (Oryza sativa) [10,11,12], alligatorweed (Alternanthera philoxeroides) [13], and wheat (Triticum aestivum) [14].
Common methods for functional research include T-DNA insertion gene mutation, electrophysiological technology, and yeast heterologous expression systems [3,9,11,12,13,14,15]. Research shows that OsAMT1;1 in rice exhibits a constitutive expression pattern, with expression in both the aerial parts and roots. In contrast, OsAMT1;3 displays root-specific expression characteristics. The expression of both OsAMT1;1 and OsAMT1;3 is induced by nitrogen starvation but repressed via glutamine-mediated feedback [10]. Using electrophysiological technology and complementation of functional yeast mutant, Yang et al. [11] verified that OsAMT1;1 is an ammonium transporter with relatively low affinity for ammonium, with a affinity constant (Km) of 110–129 μmol·L^−1^. Hao et al. [12] demonstrated that OsAMT1;3 is a typical high-affinity ammonium transporter, with a Km of approximately 32 μmol·L^−1^ and weak permeability to methylammonium (MeA^+^).
The AMT transporters play a key role in regulating ammonium uptake and maintaining nitrogen homeostasis in plants. Research on the agronomic traits of plant AMT transporters is primarily focused on the AMT1 subgroup, while studies on AMT2 agronomic traits and their regulation of ammonium uptake and metabolism remain relatively scarce. Studies using Arabidopsis T-DNA insertion or transposon mutants indicate that AtAMT1;1 and AtAMT1;3 play a dominant role in high-affinity ammonium absorption. The AtAMT1;1 single mutant results in a 30% reduction in high-affinity ammonium absorption in plants [16], while the transposon insertion mutant of AtAMT1;2 leads to a 28% reduction in high-affinity ammonium absorption in roots. The double mutant of AtAMT1;1 and AtAMT1;3 significantly inhibits plant growth, with a 70% decline in ammonium absorption capacity [15,17]. Nonetheless, AtAMT1.1, AtAMT1.2, and AtAMT1.3 together contribute approximately 95% of ammonium uptake. Homologs of these AMT proteins in rice have been reported to mediate ammonium uptake in roots and are highly expressed under conditions of excess ammonium supply [18].
Currently, plant AMT2 transporters are characterized as NH_3_ transporters [19,20]. In Arabidopsis, AtAMT2 functions as a channel-like NH_3_ flux transporter [19]. In maize, ZmAMT3;1 mediated high-affinity ammonium transport, with the substrate NH_4_^+^ being accessed, but actually transporting NH_3_ [20]. However, the biological functions of AMT2 transporters in fruit trees are essentially unexplored. In peach, PpeAMT3;4 is primarily expressed in roots and significantly down-regulated by ammonium toxicity. A ^15^N-labeled ammonium uptake assay in yeast cells revealed that PpeAMT3;4 is a typical high-affinity ammonium transporter [6]. In pear, PbAMT2 is expressed in all tested organs with the highest level detected in roots, whereas PbAMT3 is primarily expressed in leaves [21].
As a globally important perennial vine crop, grape (Vitis vinifera L.) exhibits significant cultivar-specific characteristics in nitrogen metabolism. Fruit quality parameters (e.g., soluble solids, anthocyanin content) show a strong positive correlation with nitrogen supply, and over 60% of flavor precursor compounds in wine are regulated by nitrogen nutrition [22,23]. In this study, VviAMT4;1 was isolated from ‘Yanpu No.2’ and proved to be a typical high-affinity AMT2 subgroup ammonium transporter. Favorably, this work provides a theoretical foundation for elucidating the biological functions of AMT2 transporters in fruit trees.
2. Materials and Methods
2.1. Plant Material and Growth Conditions
The table grape cultivar ‘Yanpu No.2’ was used as experimental material. The tested vines were 5-year-old mature ‘Yanpu No.2’ trees provided by Yantai Academy of Agricultural Sciences, Shandong Province, and were cultivated under consistent management conditions.
Referring to the grape developmental stage classification methods [24,25], fruit samples were collected at the young fruit stage (35 days post-anthesis, DPA) and maturity (120 DPA). Flowers at the inflorescence visible stage and full bloom, as well as new leaves (fully expanded leaves at the 4th–6th internodes of new shoots) and roots (new white absorbing roots, diameter < 2 mm), were collected at different fruit developmental stages. For each sample, 3 biological replicates were selected, with 4 clusters randomly harvested from each plant. For each cluster, 30 fruits of uniform maturity, free from mechanical damage or disease, were randomly selected.
Tissue-cultured ‘Yanpu No.2’ seedlings were germinated on half-strength MS solid medium (pH 5.8) for 1 month before being transferred to half-strength MS liquid solution in plastic incubators within a growth chamber [26,27]. For ammonium depletion (–NH_4_^+^) treatment, NH_4_Cl was omitted from the MS medium. The nutrient solution was changed every other day.
All collected samples were immediately frozen in liquid nitrogen and stored at −80 °C for subsequent analysis.
2.2. Isolation and Cloning of VviAMT4;1
Using the coding protein sequences of the peach PpeAMT3;4 and PpeAMT4;1 genes as the reference sequences [6], a potential grape AMT gene was retrieved from the Phytozome grape genome database and named VviAMT4;1. The following primers were designed: forward primer 5′-ATGGCGTCCCTGGACTGCTC-3′ (20 bp sequence starting from the initiation codon) and reverse primer 5′-TTAACAATGACGGCACATGG-3′ (20 bp sequence preceding the termination codon). Total RNA was extracted from the roots of ‘Yanpu No.2’ tissue-cultured seedlings, and first-strand cDNA was synthesized using the PrimeScript™ RT Reagent Kit (TaKaRa, Dalian, China) as the template. The VviAMT4;1 gene was amplified using PrimeSTAR™ HS DNA Polymerase (TaKaRa, Dalian, China) and sent to Sangon Biotech (Shanghai, China) Co., Ltd. for sequencing verification.
2.3. Phylogenetic Tree Construction of Plant AMT Homologues
Amino acid sequence alignment of AMT homologous proteins, including table grape VviAMT4;1, Prunus persica PpeAMT3;4 and PpeAMT4;1, Lotus japonicus LjAMT2;1, Populus trichocarpa PtrAMT4;1, Oryza sativa OsAMT2;1 and OsAMT4;1, and Arabidopsis thaliana AtAMT2;1, was conducted using ClustalX_2.0.13 software. Amino acid sequences of AMT homologues were collected from table grape, S. lycopersicum, P. persica, L. japonicus, P. trichocarpa, O. sativa, and A. thaliana. A phylogenetic tree of plant AMT homologues was constructed using the maximum likelihood method in MEGA 15.0, with a bootstrap test performed using 1000 replicates to assess the confidence of the tree.
2.4. Quantitative Real Time PCR (qRT-PCR)
Using the online NCBI/Primer-BLAST server, sequence-specific primers for VviAMT4;1 were designed (forward primer: 5′-CACCGTATGGACCAGGCTTAT-3′; reverse primer: 5′-GGCATACGCTTCCTCTCCAT-3′). Grape Ubiquitin (GenBank No. MH114011) and Tubulin (GenBank gene ID: 100246164) were used as internal reference genes [24,25,26,27,28,29,30]. The expression patterns of VviAMT4;1 in different tissues at various developmental stages were analyzed using SYBR Green (TaKaRa, Dalian, China) on an ABI 7500 Real-Time PCR System (ABI, New York, NY, USA). The PCR reaction was performed under the following conditions: 95 °C for 2 min, 1 cycle; 95 °C for 30 s, 60 °C for 30 s and 72 °C for 30 s, 28 cycles; 72 °C for 10 min, 1 cycle. Each reaction was performed with four biological replicates. The Ct values obtained from the ABI 7500 PCR system were normalized using the internal reference genes, and relative expression levels were calculated using the 2^−ΔΔCt^ method [6,13,24,25,26,27,28,29,30].
2.5. Functional Complemention of VviAMT4;1 in Yeast Mutant
The recombinant plasmid pYES2-AMT4;1 was constructed by cloning the CDS of the VviAMT4;1 gene into the pYES2 vector [11,12,13,26]. This process utilized the forward primer 5′-GAGAGGTACCATGGCGTCCCTGGACTGCTC-3′ (Kpn I underlined) and reverse primer 5′- GAGAGTCGACTTAACAATGACGGCACATGG -3′ (Sal I underlined). The resulting recombinant plasmid was subcloned into the triple-mutant yeast strain 31019b (MATa mepl△ mep2△::LEU2 mep3△::KanMX2 ura3) [11,12,13,26]. The empty vector pYES2 and the recombinant vector pYES2-AMT4;1 were separately transformed into 31019b via electroporation. Then, 1 mL of pre-chilled 1 mol·L^−1^ sorbitol solution was immediately added, and the cells were allowed to recover at 30 °C for 2 h before being plated onto yeast selection medium (0.17% YNB + 2 mmol·L^−1^ arginine + 2% D-galactose + 2% agar, pH 5.8). The plates were incubated at 30 °C in the dark for 72 h. Single colonies were picked and inoculated into liquid selection medium, followed by incubation at 30 °C with shaking at 200 rpm for 36 h. Positive recombinant yeast clones were confirmed by PCR. The positive strains were cultured in YNB liquid selection medium until the OD_600_ reached 1.0. Tenfold serial dilutions (10^0^, 10^−1^, 10^−2^, 10^−3^) were prepared, and 5 μL of each dilution was spotted onto solid test plates containing either 2 mmol·L^−1^ arginine or 2 mmol·L^−1^ NH_4_Cl as the sole nitrogen source (other components were the same as the screening medium).
To examine the effects of methyl amine (MeA^+^) or L-methionine sulfoximine (L-MSX) on the ammonium uptake activity of VviAMT4;1, 3 mmol·L^−1^ MeA^+^ or 20 μmol·L^−1^ L-MSX was added to the solid medium with 2 mmol·L^−1^ NH_4_Cl as the sole nitrogen source [11,12,13]. The plates were incubated at 30 °C in the dark for 72 h, and colony growth was recorded using a gel imaging system. Parallel control experiments were performed with the empty vector-transformed strain (pYES2/31019b).
2.6. Kinetic Analysis of 15N Isotope Tracer-Labeled Ammonium Uptake
Following the methods previously reported by Yang et al. [11] and Guo et al. [13], the ammonium uptake kinetics of VviAMT4;1 were analyzed. Yeast cells were cultured in YNB liquid medium containing ^15^N isotope-labeled ^15^NH_4_Cl, with NH_4_^+^ concentrations set at 0, 0.01, 0.025, 0.05, 0.1, 0.25, 0.5, 1, and 2 mmol·L^−1^. The ^15^N-labeled ^15^NH_4_^+^ content was measured using a FLASH EA-DELTA V Elemental Analyzer-Mass Spectrometer (Flash-2000 Delta V ADVADTAGE, Thermo Fisher Scientific, Norristown, PA, USA), and the kinetic parameters Km (affinity constant) and Vmax (maximum uptake rate) were derived from curve fitting [11,12,13].
2.7. Statistical Analysis
Graphs were generated using Origin 12.0 software, and significant differences were analyzed using Student’s t-test in SPSS 13.0 software (SPSS Chicago, IL, USA), with details provided in the legends.
3. Results
3.1. Isolation of VviAMT4;1 from Table Grape
Using the peach AMT3;4 protein sequence as a reference [6], a homologous protein (VIT_200s1818g00010) was identified in the Phytozome grape genome, which was named VviAMT4;1. InterProScan prediction revealed the presence of an ammonium transporter functional domain, suggesting it might be a potential ammonium transporter protein. The VviAMT4;1 gene was further amplified from ‘Yanpu No.2’ roots, with a coding sequence (CDS) of 1338 bp encoding 445 amino acids (Figure 1). Gene structure analysis indicated that VviAMT4;1 has two introns, with lengths of 168 bp and 95 bp, respectively. Amino acid sequence alignment showed 64.39% identity between VviAMT4;1 and plant AMT homologues from P. persica, L. japonicus, P. trichocarpa, O. sativa, and A. thaliana. In addition, these plant AMT homologues contain 11 typical transmembrane domains (Figure 1).
3.2. Phylogenetic Tree Construction and Analysis
Phylogenetic tree analysis revealed that plant AMT transporters are classified into two major subgroups: AMT1 and AMT2. Notably, VviAMT4;1 was tightly clustered with peach PpeAMT4;1 and poplar PtrANT4;5, which belong to the AMT2 subgroup. The clustering pattern suggests that AMT2 subgroup transporters, particularly those of perennial tree species, share a relatively close evolutionary distance (Figure 2).
3.3. Expression Profiles of VviAMT4;1
Both grape Ubiquitin (GenBank No. MH114011) and Tubulin (GenBank gene ID: 100246164) were evaluated as internal reference genes, and similar expression trends were obtained. In this study, results are presented using Ubiquitin as the internal reference gene (Figure 3). Tissue-specific expression analysis revealed that the VviAMT4;1 gene was most highly expressed in the roots of both mature ‘Yanpu No.2’ grapevines and tissue-cultured seedlings. In contrast, its expression levels were relatively low in other tissues or organs, including leaves and stems of seedlings, as well as flowers, leaves, phloem, and fruits of mature vines (Figure 3A).
Moreover, expression of VviAMT4;1 was strongly induced by NH_4_^+^ depletion. After 48 h of NH_4_^+^ starvation, the transcript levels of VviAMT4;1 increased in all tested tissues or organs (leaves, stems, and roots) of the tissue-cultured seedlings (Figure 3B).
3.4. Functional Determination of VviAMT4;1 in Yeast Mutant
The yeast strain 31019b failed to grow normally in a medium containing less than 5 mmol·L^−1^ NH_4_^+^ as the sole nitrogen source [11,12,13]. On yeast-selective medium with 2 mmol·L^−1^ arginine (Arg) as the sole nitrogen source (pH 5.8), both the recombinant strain (pYES2-VviAMT4;1/31019b) and the empty vector-transformed strain (pYES2/31019b) exhibited normal growth. However, on the screening medium containing 2 mmol·L^−1^ NH_4_Cl as the sole nitrogen source, the empty vector-transformed strain pYES2/31019b could not grow, whereas the recombinant strain pYES2-VviAMT4;1/31019b restored growth under varying pH conditions (Figure 4). These findings suggest that VviAMT4;1 can mediate ammonium uptake in yeast cells. Meanwhile, we speculate that VviAMT4;1 may recognize NH_4_^+^ but could be implicated in NH_3_ transport.
MeA^+^ is an analog of NH_4_^+^ and significantly affects NH_4_^+^ uptake [11,12,13]. On the screening medium supplemented with both 3 mmol·L^−1^ MeA^+^ and 2 mmol·L^−1^ NH_4_Cl, the empty vector-transformed strain (pYES2/31019b) failed to grow, whereas the recombinant vector-transformed strain (pYES2-VviAMT4;1/31019b was able to grow but exhibited weakened growth compared to the control screening medium containing 2 mmol·L^−1^ NH_4_Cl as the sole nitrogen source. On yeast screening medium containing 3 mmol·L^−1^ MeA^+^ as the sole nitrogen source, the empty vector-transformed strain pYES2/31019b failed to grow, whereas the recombinant strain pYES2-VviAMT4;1/31019b exhibited weak growth (Figure 5). L-MSX is a glutamine analog that inhibits NH_4_^+^ assimilation [12,13]. On yeast screening medium containing 2 mmol·L^−1^ NH_4_Cl plus 20 μmol·L^−1^ L-MSX, or containing 20 μmol·L^−1^ L-MSX as the sole nitrogen source, neither the recombinant strain pYES2-VviAMT4;1/31019b nor the empty vector-transformed strain pYES2/31019b could grow (Figure 5).
3.5. 15N-Labeled Ammonium Uptake Kinetics Analysis
Km reflects the affinity between the AMT transporter and its substrate and Vmax reflects the catalytic efficiency of AMT transporters [11,12,13]. The ^15^N isotope tracer analysis revealed that VviAMT4;1-mediated ammonium uptake exhibited a Km of 49.58 ± 4.66 μmol·L^−1^ and a Vmax of 3.29 μmoles·min^−1^·μg^−1^ cells (Figure 6), indicating that VviAMT4;1 is a typical high-affinity ammonium transporter.
4. Discussion
Ammonium is one of the primary forms of nitrogen utilized by plants. When soil nitrogen is deficient, ammonium becomes the preferred absorption form for most plant species [1,2,3,5]. In particular, AMT transporters mediate the uptake and intracellular transport of ammonium (NH_4_^+^ or NH_3_), playing a critical role in plant growth and development. The AMT family comprises over a dozen members divided into two major subgroups, AMT1 and AMT2, with the majority of their gene functions remaining uncharacterized. Current research primarily focuses on functional analyses of the AMT1 subgroup in annual model plants, like Arabidopsis and rice [3,4,5,6,7,8,9,10,11,12]. Due to their perennial growth habit and specific lignified stems, woody and liana plants exhibit distinct nitrogen absorption and transport mechanisms compared to herbaceous species, suggesting differences in the physiological functions and regulatory mechanisms of their AMT homologs [31,32]. Nevertheless, the biological functions of AMT2 transporters in perennial woody and liana fruit crops remain largely unclear.
In model plants, multiple AMT1 subgroup homologues, such as Arabidopsis AtAMT1;3 [33], rice OsAMT1;1 [10,11], OsAMT1;3 [10,12], and Alternanthera philoxeroides ApAMT1;1 [13], are predominantly highly expressed in roots, mediating ammonium uptake or transport in root cells [10,11,12,32]. Interestingly, an AMT2 subgroup-encoding gene, VviAMT4;1, was cloned from ‘Yanpu No.2’. This gene was highly expressed in roots of both tissue-cultured seedlings and 5-year-old mature trees, suggesting its potential role in ammonium uptake and transport in cells of table grape roots. Similarly, PpeAMT4;3 was also highly expressed in peach roots [6] and PbAMT2 was highly expressed in pear roots [21]. Simultaneously, VviAMT4;1 shares the closest evolutionary relationship with peach PpeAMT4;1 and poplar PtrAMT4;5, implying that AMT2 subgroup transporters of perennial trees may possess similar biological functions, which requires further validation. Furthermore, VviAMT4;1 exhibits lower expression in leaves, stems, and fruits, indicating its comprehensive involvement in NH_4_^+^ allocation and transport in above-ground tissues or organs.
In plants, AMT2 family members are functionally recognized as NH_3_ transporters [19,20]. In this study, yeast functional complementation and ^15^N isotope tracer kinetic assays revealed that VviAMT4;1 is a high-affinity ammonium transporter that may be involved in NH_3_ transport, which needs further biological verification. These findings are in accordance with Arabidopsis AtAMT2 [19] and maize ZmAMT3;1 [20]. Together, these findings support the proposition that NH_4_^+^ is firstly recruited by the binding site and then transported across the membrane in the form of uncharged NH_3_ by dehydrogenation in AMT2 transporters.
Notably, MeA^+^ impacts ammonium uptake [11,12,13]. This study demonstrates that MeA^+^ as a substrate analog inhibits ammonium uptake and utilization in yeast cells, consistent with findings in rice OsAMT1;1 [11] and OsAMT1;3 [12] and A. philoxeroides ApAMT1;1 [13]. These findings suggest that VviAMT4;1 may play a role in nitrogen nutrition dynamics and efficient utilization in table grape roots. In addition, when ammonium and MeA^+^ coexist, VviAMT4;1 mediates preferential ammonium utilization, ensuring weak growth of yeast mutants. However, when MeA^+^ is the sole nitrogen source, VviAMT4;1 can also facilitate trace MeA^+^ uptake. The addition of L-MSX abolished VviAMT4;1-mediated ammonium uptake, aligning with results from rice OsAMT1;3 [12] and alligatorweed ApAMT1;1 [13]. Since L-MSX is a glutamine analog primarily inhibiting glutamine synthetase [12,13], it is speculated that ammonium absorbed by plant AMTs may participate in glutamine synthesis. L-MSX blocks this pathway, limiting ammonium assimilation and thereby suppressing VviAMT4;1 function. These findings indicate that VviAMT4;1 may be subject to feedback inhibition by intracellular ammonium accumulation.
Favorably, this study provides genetic resources for elucidating the biological functions of AMT2 transporters in table grapes and lays a theoretical foundation for exploring the molecular mechanisms of VviAMT4;1 in efficient nitrogen utilization.
5. Conclusions
VviAMT4;1 was isolated from table grape ‘Yanpu No.2’. It was highly expressed in roots and showed up-regulated expression under NH_4_^+^ depletion. VviAMT4;1 is a typical high-affinity ammonium transporter that may recognize NH_4_^+^ but actually transports NH_3_. Both MeA^+^ and L-MSX reduced the growth of yeast mutants transformed with pYES2-VviAMT4;1. The VviAMT4;1-mediated ammonium uptake process may be subject to feedback inhibition by endogenous NH_4_^+^ enrichment.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Cruz C. Bio A.F.M. Dominguez-Valdivia M.D. Aparicio-Tejo P.M. Lamsfus C. Martins-Loução M.A. How does glutamine synthetase activity determine plant tolerance to ammonium?Planta 20062231068108010.1007/s 00425-005-0155-216292661 · doi ↗ · pubmed ↗
- 2Liu Y. von Wirén N. Ammonium as a signal for physiological and morphological responses in plants J. Exp. Bot.2017682581259210.1093/jxb/erx 08628369490 · doi ↗ · pubmed ↗
- 3Ludewig U. Neuhäuser B. Dynowski M. Molecular mechanisms of ammonium transporter and accumulation in plants FEBS Lett.20075812301230810.1016/j.febslet.2007.03.03417397837 · doi ↗ · pubmed ↗
- 4Mc Donald T.R. Ward J.M. Evolution of electrogenic ammonium transporters (AM Ts)Front. Plant Sci.2016735210.3389/fpls.2016.0035227066024 PMC 4814505 · doi ↗ · pubmed ↗
- 5Straub T. Ludewig U. Neuhäuser B. The kinase CIPK 23 inhibits ammonium transport in Arabidopsis thaliana Plant Cell 20172940942210.1105/tpc.16.0080628188265 PMC 5354196 · doi ↗ · pubmed ↗
- 6You S.H. Wang Y.Q. Li Y.J. Li Y.H. Tan P. Wu Z. Shi W.J. Song Z.Z. Cloning and functional determination of ammonium transporter Ppe AMT 3; 4 in peach Biomed. Res. Int.202021473671710.1155/2020/2147367 PMC 773237533344631 · doi ↗ · pubmed ↗
- 7Ninnemann O. Jauniaux J.C. Frommer W.B. Identification of a high affinity NH 4+ transporter from plants EMBO J.1994133464347110.1002/j.1460-2075.1994.tb 06652.x 8062823 PMC 395249 · doi ↗ · pubmed ↗
- 8Ludewig U. von Wiren N. Frommer W.B. Uniport of NH 4+ by the root hair plasma membrane ammonium transporter Le AMT 1; 1J. Biol. Chem.2002277135481355510.1074/jbc.M 20073920011821433 · doi ↗ · pubmed ↗