Chloride-Transporting OsHKT1;1 Splice Variants and Their Expression Profiles Under Salinity Stress in Rice
Shahin Imran, Shuntaro Ono, Rie Horie, Maki Katsuhara, Tomoaki Horie

TL;DR
This study investigates how different versions of a rice transporter protein handle salt stress, revealing that one variant mainly transports chloride instead of sodium.
Contribution
The study reveals the chloride-transporting function of OsHKT1;1-V2 splice variant, contrasting with the sodium-selective full-length variant.
Findings
OsHKT1;1-V2 primarily transports Cl− with weak Na+ selectivity, unlike the Na+-selective OsHKT1;1-FL.
OsHKT1;1-V2 localizes to the plasma membrane and accumulates Cl− in oocytes.
Nipponbare rice has higher OsHKT1;1-FL transcripts and lower OsHKT1;1-V2 transcripts under salt stress compared to Pokkali.
Abstract
OsHKT1;1, a member of the high-affinity K+ transporter (HKT) family, plays a key role in Na+ homeostasis and salinity tolerance in rice. In our previous study, multiple potential OsHKT1;1 splicing variants were identified, as well as the full-length (FL) OsHKT1;1 transcript from the salt-tolerant rice Pokkali. However, most previous studies focused solely on the full-length protein, leaving the transport functions of splice variants largely unexamined. In this study, we focused on the splice variant OsHKT1;1-V2 and compared its function and gene expression with those of OsHKT1;1-FL. Two-electrode voltage clamp experiments using Xenopus laevis oocytes revealed that the 1st start codon of OsHKT1;1-V2 is functional to exhibit bidirectional currents in bath solutions containing NaCl. Unlike the Na+-selective feature of OsHKT1;1-FL, OsHKT1;1-V2 primarily mediated Cl− transport with weak Na+…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
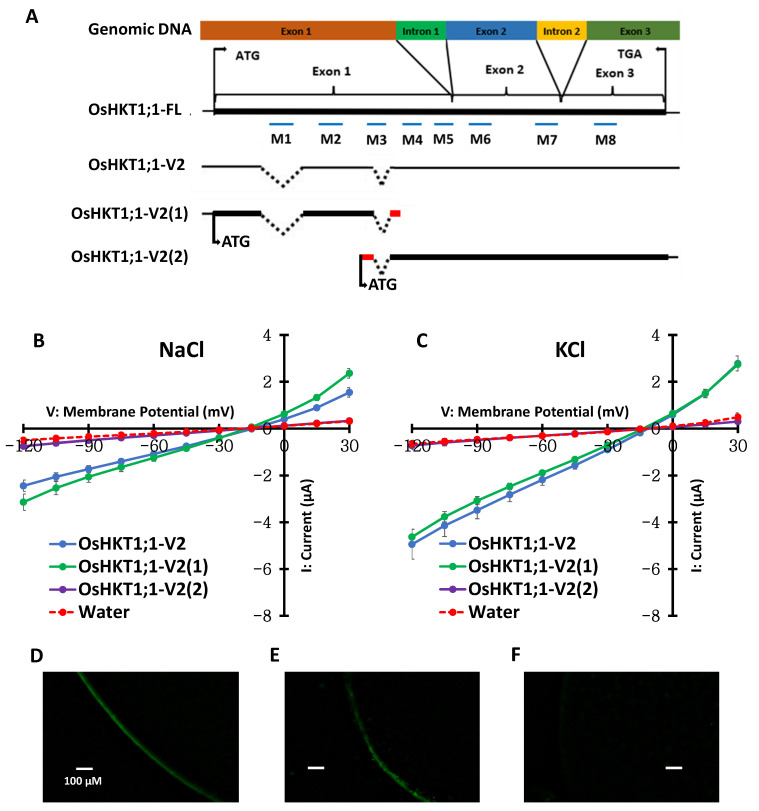 Figure 1
Figure 1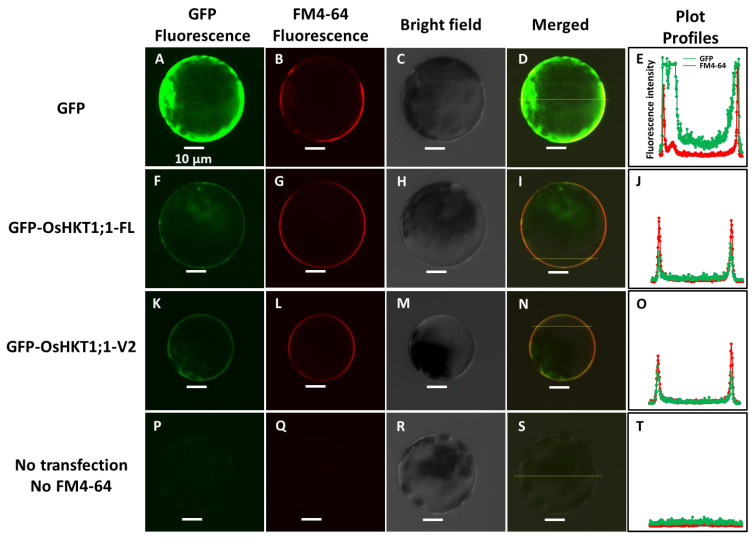 Figure 2
Figure 2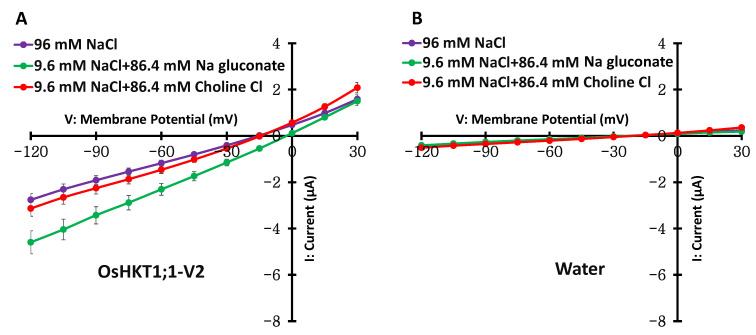 Figure 3
Figure 3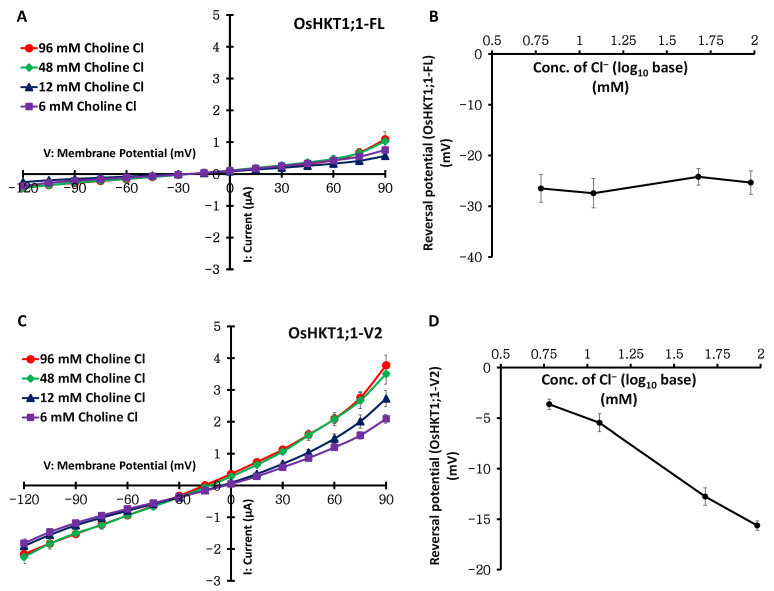 Figure 4
Figure 4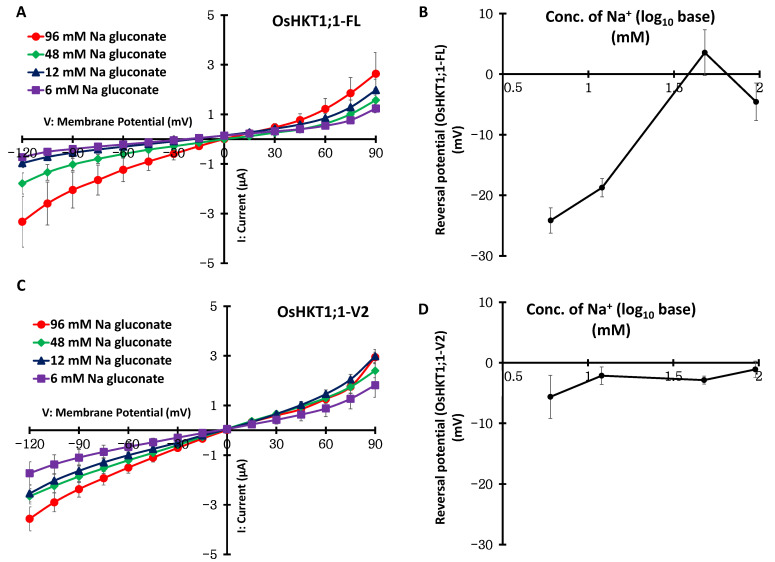 Figure 5
Figure 5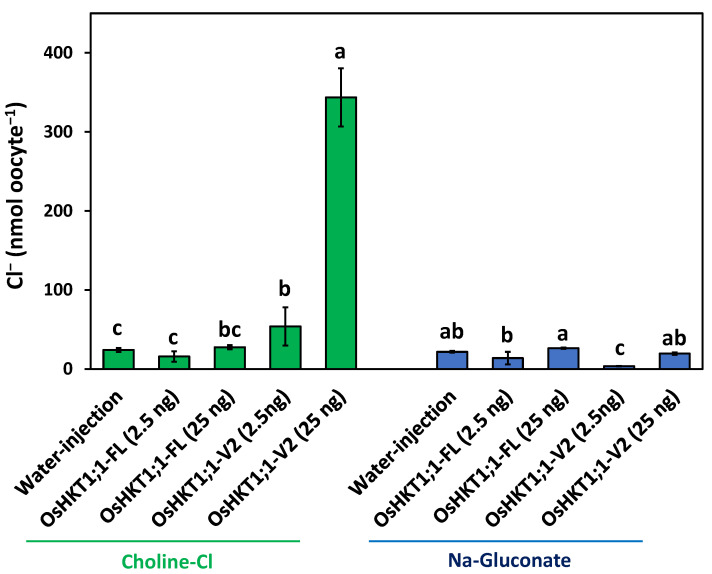 Figure 6
Figure 6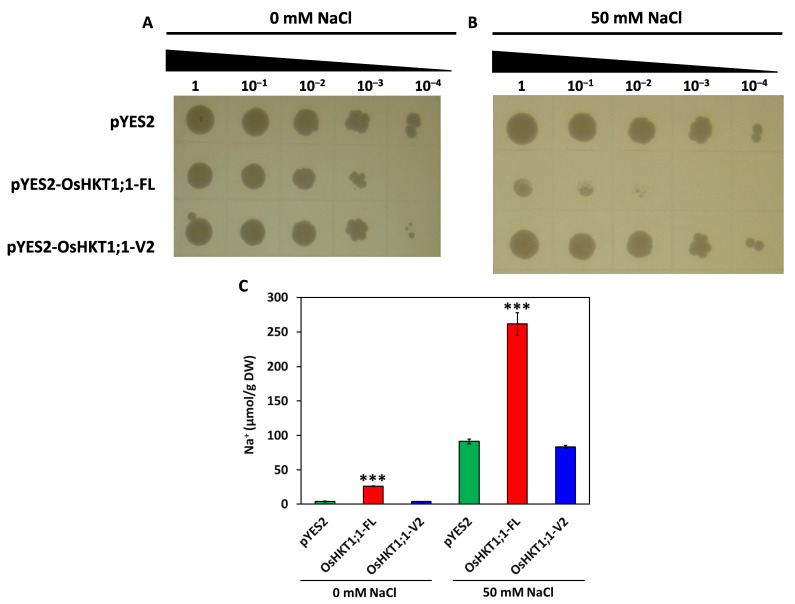 Figure 7
Figure 7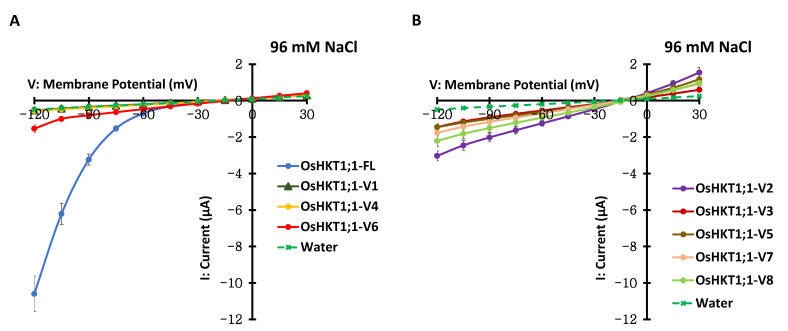 Figure 8
Figure 8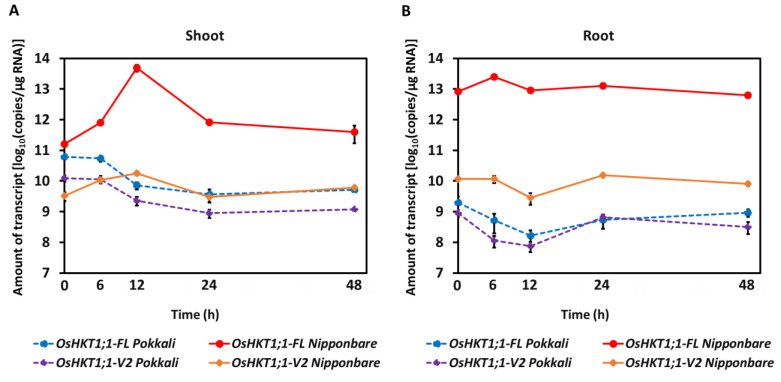 Figure 9
Figure 9- —JSPS KAKENHI
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Stress Responses and Tolerance · Plant nutrient uptake and metabolism · Aluminum toxicity and tolerance in plants and animals
1. Introduction
High concentrations of soluble or exchangeable salts (in most cases, the cation is Na^+^) inhibit plant growth under salinity stress, which is a globally alarming issue that significantly lowers the yield and development of major crops. Rice (Oryza sativa) is the most susceptible to salt stress among cereal crops [1,2]. When toxic Na ions are adequately excluded, redistributed, and compartmentalized in the plant body, the plants are likely to exhibit tolerance to salinity stress.
High-affinity K^+^ transporters (HKTs) are known to be involved in the transport of Na^+^ and K^+^ in plants, and some of them are crucial for maintaining Na^+^/K^+^ balance under salinity stress [3,4]. Therefore, HKTs have been widely examined since the first report of the TaHKT2;1 gene in bread wheat (Triticum aestivum) [5]. HKTs are classified into two subfamilies, and transporters belonging to subfamily 1 (HKT1s) have generally been shown to be selective for Na^+^ [6,7,8,9]. The ability of plants to withstand salt has been demonstrated for both HKT1s and HKT2s.
In monocotyledon rice, the japonica cultivar was reported to maintain seven functional HKT genes, among which four genes (OsHKT1;1, OsHKT1;3, OsHKT1;4, and OsHKT1;5) were grouped in subfamily 1 [10,11]. Heterologous expression studies of OsHKT1;1 from rice using Xenopus laevis (X. laevis) oocytes showed Na^+^-selective transport in an inward-rectifying manner [8,12]. HKT1;1 transporters identified in various plant species exhibit robust Na^+^ selectivity when expressed in Xenopus laevis oocytes: AtHKT1;1 from Arabidopsis [7], HvHKT1;1 from barley [13], CmHKT1;1 from pumpkin [14], SvHKT1;1 from halophytic turf grass (Sporobolus virginicus) [15], VviHKT1;1 from grapevine [16], and VcHKT1;1 from blueberry [17]. Some HKT1 transporters interact with other ion homeostasis systems, such as the SOS pathway and K^+^ transporters [18,19], forming a coordinated regulatory network that maintains ionic balance and cellular stability under salinity stress [20,21]. OsHKT1;1 has been shown to be involved in limiting Na^+^ accumulation in leaves and contributing to salt tolerance based on the analysis of an insertional mutation in the OsHKT1;1 gene [22]. Recent studies have further demonstrated that natural allelic variations in OsHKT1;1 contribute significantly to genotypic differences in Na^+^ distribution within rice. Genome-wide association analysis using a global diversity panel identified OsHKT1;1 as the gene underlying a major QTL (RNC4) that regulates root Na^+^ content, with indica accessions generally carrying alleles that promote higher root Na^+^ accumulation than japonica accessions [23]. Functional assays revealed that three indica-predominant non-synonymous substitutions increased the inward Na^+^ transport activity of OsHKT1;1, and introduction of the indica allele into a japonica background reproduced the high-root-Na^+^ phenotype. These results support earlier findings that OsHKT1;1 limits Na^+^ accumulation in shoots and improves salt tolerance [22]. Furthermore, a combination of screening of salt-tolerant rice mutant lines and the MutMap method identified a key transcription factor gene that encodes a B-type response regulator OsRR22, suggesting the involvement of OsHKT1;1 in rice salt tolerance [24].
In our previous study, an interesting phenomenon was found in the OsHKT1;1 gene in a salt-tolerant landrace, Pokkali, in which potential alternative splicing variants encoding transporters that show distinct ion selectivity were identified in addition to the full-length OsHKT1;1 transcript [8]. Alternative splicing (AS) is a post-transcriptional process that fine-tunes gene function. In rice, AS is broadly reprogrammed by salinity and other abiotic stresses, generating isoforms that adjust protein abundance and activity as conditions change [25]. This regulatory flexibility is evident in genes such as OsDREB2B, OsHSFA2d, and OsMYB7, where stress-dependent switching to functional splice variants enhances stress tolerance [25,26,27,28]. Similar patterns in other species reinforce the central role of AS in stress adaptation. In foxtail millet, for example, salt stress induces two SiCYP19 splice variants, and SiCYP19-b more effectively improves tolerance by increasing proline and reducing ROS [29]. In Populus euphratica, AS of PeuHKT1;3 generates the isoform PeuHKT1;3a, which alters pore-loop structure, enhances K^+^ transport, and improves K^+^/Na^+^ homeostasis [30]. In Arabidopsis, At-SRAS1 produces two splice variants with opposite functions: At-SRAS1.1 (full-length, active E3 ligase) enhances salt tolerance, while At-SRAS1.2 (truncated) reduces it [31]. In the halophyte Thellungiella salsuginea, salt stress induces multiple splice variants of TsHKT1;2, among which the functional isoforms (TsHKT1;2a/2b) interact with other HKT1 proteins to reduce Na^+^ uptake and enhance salt tolerance [32]. In rice, OsHKT1;4 also shows tissue-dependent splice variation that affects Na^+^ retrieval and supports ionic balance under salinity [33]. Despite these findings, AS regulation of HKTs in rice remains largely uncharacterized, and only a few other studies have reported splicing variants of HKTs [9,34].
Although alternative splicing of OsHKT1;1 has been reported, the functional analyses of these variants, particularly whether they exhibit different transport characteristics from the full-length protein, remain unclear. Therefore, in the present study, we focused on one of the eight variants of the OsHKT1;1 gene from Pokkali, OsHKT1;1-V2, to characterize its expression profile and transport properties. We present the results of heterologous expression analyses, which exhibit Cl^−^ transport features with far less selectivity for Na^+^ of OsHKT1;1-V2 than those of full-length OsHKT1;1. Furthermore, we showed evidence that identical potential splicing variants exist in a salt-sensitive japonica cultivar, Nipponbare, and that OsHKT1;1-V2 from Nipponbare also exhibits similar Cl^−^ transport properties, although the expression levels of the full-length and OsHKT1;1-V2 variant seemed to be differentially regulated between the two varieties. We provide new insights into the mechanisms of ion homeostasis in rice, mediated by the potential alternative splicing of an important salt tolerance gene, OsHKT1;1, and discuss questions to be elucidated for pursuing the physiological impacts.
2. Results
2.1. Identification of a Functional Frame in the OsHKT1;1-V2 Variant
In our previous study, two start codons were identified in the OsHKT1;1-V2 variant (Figure 1A) [8]. We artificially truncated and produced two fragments of OsHKT1;1-V2 named OsHKT1;1-V2(1) and OsHKT1;1-V2(2). OsHKT1;1-V2(1) is composed of 519 nucleotides, encoding 172 putative amino acid residues. In contrast, OsHKT1;1-V2(2) is composed of 957 nucleotides encoding 318 putative amino acid residues (Figure 1A, Supplementary Figure S1). X. laevis oocytes were injected with cRNAs of OsHKT1;1-V2, OsHKT1;1-V2(1), and OsHKT1;1-V2(2), and TEVC experiments were performed to identify which fragment induces bidirectional ionic currents like OsHKT1;1-V2 [8]. OsHKT1;1-V2(1) but not OsHKT1;1-V2(2) showed ionic currents in both NaCl and KCl bath solutions as OsHKT1;1-V2 (Figure 1B,C). These results suggest that the start codon of OsHKT1;1-V2 is the 1st methionine. Furthermore, histochemical analysis indicated that FLAG tag-fused OsHKT1;1-FL and OsHKT1;1-V2 proteins were localized in the plasma membrane (PM) of oocytes (Figure 1D,E). In contrast, control oocytes injected with water showed no robust green fluorescence in the PM (Figure 1F).
2.2. Subcellular Localization of GFP-Fused OsHKT1;1-FL and OsHKT1;1-V2 in Plant Cells
To study the subcellular localization in plant cells, green fluorescence protein (GFP) was fused with OsHKT1;1-FL and OsHKT1;1-V2 at the N-terminus. DNA constructs to express either GFP, GFP-OsHKT1;1-FL, or GFP-OsHKT1;1-V2 were introduced into Arabidopsis leaf protoplasts [35]. Cells expressing GFP showed strong fluorescence in the cytosol, which did not overlap with the plasma membrane marker FM4-64 (Figure 2A–D). GFP fluorescence from cells expressing GFP-OsHKT1;1-FL was observed at the cell periphery, overlapping with FM4-64 fluorescence (Figure 2F–I). A similar overlap of GFP and FM4-64 fluorescence was observed in cells expressing GFP-OsHKT1;1-V2 (Figure 2K–N). Plot-profile fluorescence intensity analysis of the merged fluorescence images supported the above-mentioned observation (Figure 2E,J,O). Together with the negative control experiments shown in Figure 2P–S, these results suggest the plasma membrane localization of OsHKT1;1-V2 as OsHKT1;1-FL.
2.3. Substrate Selectivity of OsHKT1;1-FL and OsHKT1;1-V2
Current–voltage relationships were obtained in the presence of 96 mM NaCl (96 mM Na^+^ and 96 mM Cl^−^), 9.6 mM NaCl + 84.6 mM Na gluconate (total 96 mM Na^+^ and 9.6 mM Cl^−^), and 9.6 mM NaCl + 84.6 mM Choline Cl (9.6 mM Na^+^ and total 96 mM Cl^−^) by conducting electrophysiological TEVC experiments using X. laevis oocytes. As described previously [12], a decrease in Na^+^ concentration from 96 mM to 9.6 mM largely reduced the inward currents in OsHKT1;1-FL and showed no tendency to produce robust outward currents (Supplementary Figure S2). In contrast, the same change in solution did not affect OsHKT1;1-V2-mediated currents (Figure 3A). Next, we changed the Cl^−^ concentration of the bath solution while keeping the Na^+^ concentration at 96 mM during TEVC measurements. The 10 times reduction in the Cl^−^ concentration led to an increase in OsHKT1;1-V2-mediated currents with a positive shift in the reversal potential of 13 mV (Figure 3A, purple and green lines). Note that similar large currents were never observed in water-injected oocytes under any bath condition (Figure 3B).
Next, oocytes expressing OsHKT1;1-V2 were analyzed in 96 mM NaCl (96 mM Na^+^ and 96 mM Cl^−^), 96 mM Na gluconate (total 96 mM Na^+^ and 0 mM Cl^−^), and 96 mM Choline Cl (0 mM Na^+^ and total 96 mM Cl^−^) solutions with voltage steps from +90 to −120 mV in 15 mV decrements to examine outward currents. OsHKT1;1-V2 expressed in oocytes exhibited large outward currents with no reversal potential shift when the solution was changed from 96 mM NaCl to 96 mM Choline Cl, by which Na^+^ concentration was decreased from 96 mM to 0 mM (Supplementary Figure S3A, purple and red lines). In contrast, reversal potential shifts occurred according to a change in Cl^−^ concentration from 96 mM to 0 mM in OsHKT1;1-V2 (Supplementary Figure S3A, purple and red lines vs. green line). Note that endogenous outward currents were also observed in water-injected oocytes, but they were never the same extent as OsHKT1;1-V2-expressing oocytes in every bath condition (Supplementary Figure S3B). These results suggest that the currents induced by OsHKT1;1-V2 largely depend on Cl^−^.
2.4. Substrate Concentration Dependency of OsHKT1;1-FL and OsHKT1;1-V2
Further TEVC measurements were performed using oocytes expressing OsHKT1;1-FL or OsHKT1;1-V2 in a series of bath solutions that contained Cl^−^ as the sole permeable monovalent anion (96, 48, 12, and 6 mM as choline chloride) or Na^+^ as the sole permeable monovalent cation (96, 48, 12, and 6 mM as Na gluconate). OsHKT1;1-FL-expressing oocytes showed currents comparable to those of water-injected control oocytes, with no reversal potential shift in a series of changes in Cl^−^ concentration (Figure 4A,B, Supplementary Figure S4B). However, OsHKT1;1-FL showed large and various currents with reversal potential shifts in a series of changes in the Na^+^ concentration (Figure 5A,B). These data suggest that OsHKT1;1-FL mediates the transport of Na^+^. In contrast, oocytes expressing OsHKT1;1-V2 exhibited robust inward and outward currents with reversal potential shifts in a series of changes in the Cl^−^ concentration (Figure 4C,D). In contrast, OsHKT1;1-V2-expressing oocytes showed larger currents than water-injected control oocytes in accordance with a series change in the Na^+^ concentration; however, the reversal potential did not alter as in oocytes expressing OsHKT1;1-FL (Figure 5B–D, Supplementary Figure S4A). These data suggest that, unlike OsHKT1;1-FL, OsHKT1;1-V2 is selective for Cl^−^ and mediates its transport.
2.5. Cl− Contents in Oocytes
Cl^−^ accumulation was investigated using oocytes injected with water, OsHKT1;1-FL cRNA, or OsHKT1;1-V2 cRNA, which were incubated in 96 mM choline chloride-containing medium (Figure 6). The total Cl^−^ content showed that OsHKT1;1-V2-expressing oocytes (2.5 ng cRNA) accumulated significantly more Cl^−^ than OsHKT1;1-FL-expressing (2.5 ng cRNA) and water-injected oocytes (Figure 6). Furthermore, a 10-fold increase in the injection volume of the OsHKT1;1-V2 cRNA (25 ng) resulted in the accumulation of approximately 6 times more Cl^−^ (Figure 6). To verify that the observed Cl^−^ increase was specifically due to Cl^−^ influx and not caused by nonspecific anion accumulation, the same experiment was performed in Na-gluconate medium without Cl^−^. Under these Cl^−^-free conditions, all oocytes, including those expressing OsHKT1;1-V2, showed low Cl^−^ levels (Figure 6). These results support that OsHKT1;1-V2 mediates Cl^−^ transport in the PM of oocytes, as suggested by the TEVC experiments shown in Figure 3 and Figure 4.
2.6. Functional Characterization of OsHKT1;1 Transporters in Yeast
OsHKT1;1-FL and OsHKT1;1-V2 were introduced into the Na^+^-sensitive mutant yeast strain G19 to further examine the Na^+^ transport capacity of OsHKT1;1-FL and OsHKT1;1-V2. No remarkable difference was found in the growth among OsHKT1;1-FL, OsHKT1;1-V2, and empty vector-harboring G19 cells under the control condition (Figure 7, left panel). However, the presence of 50 mM NaCl caused severe growth defects in OsHKT1;1-FL-expressing cells. In contrast, cells expressing OsHKT1;1-V2 grew similarly to the vector-harboring control cells (Figure 7, right panel). These data indicate that OsHKT1;1-FL, but not OsHKT1;1-V2, possibly accelerated Na^+^ uptake in yeast cells, leading to toxicity and growth reduction in the cells under NaCl-added conditions. Thus Na^+^ contents of G19 transformants were determined under no or 50 mM NaCl conditions (Figure 7C). After 18 and 21 h incubation, for control and NaCl-added conditions, respectively, in the presence of galactose, G19 cells expressing OsHKT1;1-FL were found to accumulate significantly higher levels of Na^+^ than the others in both conditions (Figure 7C). Whereas Na^+^ contents of vector control and OsHKT1;1-V2 lines were at similar levels (Figure 7C). These results were consistent with the growth phenotype (50 mM NaCl; Figure 7B).
2.7. Transport Properties of OsHKT1;1-FL and Variants from Nipponbare
In a previous study [8] and in Figure 1, Figure 2, Figure 3, Figure 4, Figure 5, Figure 6 and Figure 7 in the present study, possible OsHKT1;1 splicing variants, in addition to the full-length cDNA from salt-tolerant indica rice, Pokkali, were investigated. Therefore, we attempted to determine whether these OsHKT1;1 splicing variants could also be found in the salt-sensitive japonica rice, Nipponbare, and whether they functioned in a manner similar to those from Pokkali. OsHKT1;1 cDNAs were isolated from Nipponbare, and their transport properties were examined by TEVC experiments. Eight possible OsHKT1;1 variants were detected in addition to the full-length cDNA from Nipponbare as Pokkali [8]. Comparative analysis revealed amino acid substitutions in OsHKT1;1 between the two genotypes (Supplementary Figure S5A–I). However, oocytes expressing Nipponbare (Ni) OsHKT1;1-FL and OsHKT1;1-V6 exhibited inward-rectifying currents in the presence of 96 mM NaCl, with much smaller currents of OsHKT1;1-V6 than OsHKT1;1-FL (Figure 8A). In contrast to Ni-OsHKT1;1-V1 and -V4, whose currents were similar to those from water-injected control oocytes (Figure 8A), Ni-OsHKT1;1-V2, -V3, -V5, -V7, and-V8 mediated ionic currents in a bidirectional manner, depending on the membrane voltage (Figure 8B). There was no remarkable difference in the pattern of elicited currents mediated by these variants between Pokkali and Nipponbare.
Next, oocytes expressing Ni-OsHKT1;1-FL and Ni-OsHKT1;1-V2 were examined in Cl^−^ solutions (96, 48, 12, and 6 mM as choline chloride) (Supplementary Figure S6). Current–voltage relationships obtained from Ni-OsHKT1;1-FL expressed in oocytes showed small or no currents and no reversal potential shift with changes in Cl^−^ concentrations in the bath solution (Supplementary Figure S6A,B). In contrast, oocytes injected with OsHKT1;1-V2 cRNA exhibited both inward and outward currents with apparent reversal potential shifts according to the Cl^−^ concentration (Supplementary Figure S6C,D). These data suggest that Ni-OsHKT1;1-V2, but not Ni-OsHKT1;1-FL, mediates Cl^−^ transport, as observed in those transporters from Pokkali.
2.8. Expression Analyses of OsHKT1;1-FL and OsHKT1;1-V2 in Shoots and Roots
We next investigated the transcript levels of OsHKT1;1-FL and OsHKT1;1-V2 in Pokkali and Nipponbare by conducting qPCR-based absolute quantification. Without stress, Nipponbare plants accumulated approximately five times more OsHKT1;1-FL transcripts than Pokkali in shoots (Figure 9A). However, in the case of OsHKT1;1-V2, the transcript level in shoots was approximately six times higher in Pokkali than in Nipponbare (Figure 9A at time 0; note that the scale for transcript amounts is log-10-based). Interestingly, salt stress (100 mM NaCl) temporally increased both OsHKT1;1-FL and OsHKT1;1-V2 transcripts in Nipponbare, peaking at 12 h, whereas those of Pokkali decreased (Figure 9A). In roots, the OsHKT1;1-FL transcript level was much lower in Pokkali than in Nipponbare, and so was the level of the OsHKT1;1-V2 transcript under control conditions (Figure 9B). The OsHKT1;1-FL transcript level was temporally increased, peaking at 6 h after salt stress in Nipponbare, but this observation was not applied to Pokkali (Figure 9B). The levels of the other transcripts were maintained relatively constant during the periods investigated (Figure 9B). These results indicate a differential regulatory mechanism of OsHKT1;1 gene expression between rice genotypes.
3. Discussion
HKT membrane proteins are known to be relevant to Na^+^ homeostasis in plants, including under salinity stress. Na^+^ exclusion protects photosynthetic tissues from salt-induced damage [6], and some of the Na^+^-selective class I HKT (HKT1) transporters play a key role in excluding Na^+^ from leaves, enhancing salinity tolerance [4,22,36,37,38,39]. In rice, OsHKT1;1 and OsHKT1;3 exhibit strong and weak inward rectification for Na^+^ transport, respectively [8,9,12]. Many other studies on the homologue of HKT1;1 from barley, pumpkin, grapevine, blueberry, and wild turf Sporobolus virginicus have also revealed Na^+^-selective transport as OsHKT1;1 [13,14,15,16,17]. Campbell et al. [23] showed that natural allelic variants of OsHKT1;1 underlie the divergence in root Na^+^ content between japonica and indica rice cultivars. They reported three non-synonymous mutations associated with higher OsHKT1;1 activity and Na^+^ transport efficiency, explaining the genetic basis for the diversity in salt tolerance in rice. This study also provides strong genetic evidence that OsHKT1;1 plays a central role in Na^+^ homeostasis and is an important target for improving salinity tolerance through molecular breeding. Additionally, OsHKT1;1 interacts with the Salt Overly Sensitive (SOS) pathway and other transporters, implying a complex regulatory network for ion balance and salt stress response [11,18,20,40].
In our previous study, eight OsHKT1;1 variants (V1-V8) that could be the product of alternative splicing were identified in addition to the full-length OsHKT1;1-FL cDNA in the salt-tolerant rice cultivar Pokkali [8]. In this study, we focused on the variant OsHKT1;1-V2 as the transcript level was significantly higher than that of OsHKT1;1-FL in shoots of Pokkali, and OsHKT1;1-V2 cRNA-injected oocytes exhibited larger inward and outward currents than other variants in both NaCl and KCl bath solutions [8]. Although two start codons are present in OsHKT1;1-V2 mRNA, the present study demonstrated that the first start codon of OsHKT1;1-V2 is functional (Figure 1A–C). The plasma membrane localization of FLAG-tagged OsHKT1;1-V2 in oocytes (Figure 1E) indicated that the ionic currents elicited by OsHKT1;1-V2 (Figure 1, Figure 3, Figure 4 and Figure 5) were not artifacts or leaks. In the present study, the co-localization of GFP-fused OsHKT1;1-FL or OsHKT1;1-V2 with the dye FM4-64 in the plasma membrane was further demonstrated in Arabidopsis protoplasts (Figure 2), supporting the above-mentioned notion.
OsHKT1;1-FL was confirmed to mediate Na^+^ transport, as reported previously [8] (Figure 7, Supplementary Figure S2). Interestingly, however, OsHKT1;1-V2 displayed distinct selectivity for Na^+^ and Cl^−^: the OsHKT1;1-V2 protein expressed in oocytes was responsive to the changes in the Cl^−^ concentration but not the Na^+^ concentration, suggesting a preference for Cl^−^ transport over Na^+^ transport (Figure 3, Supplementary Figure S3). OsHKT1;1-FL showed a concentration-dependent reversal potential shift only in Na^+^ bath solutions, whereas OsHKT1;1-V2 showed no concentration-dependent reversal potential shifts in Na^+^ solutions (Figure 5, Supplementary Figure S7). In contrast to the lack of reversal potential shifts in OsHKT1;1-V2-expressing oocytes upon Na^+^ concentration changes, the observed apparent reversal potential shifts upon Cl^−^ concentration changes again suggested that OsHKT1;1-V2 is more selective for Cl^−^ (Figure 4 and Figure 5). This electrophysiological suggestion was further supported by the measurements of Cl^−^ contents of oocytes expressing OsHKT1;1-V2, which accumulated significantly more Cl^−^ than the oocytes expressing OsHKT1;1-FL after incubation in the examination solution (Figure 6). In fact, oocytes expressing OsHKT1;1-V2 (2.5 ng cRNA) in the medium exhibited an average reversal potential of −17 mV (Supplementary Figure S3A), while the resting potential of the oocytes was approximately −9 mV, which was +8 mV higher than the reversal potential, indicating that Cl^−^ influx can be triggered under the tested conditions. The obtained results were consistent with those from the TEVC analysis (Figure 3, Figure 4 and Figure 5, Supplementary Figure S3). On the other hand, functional assays using the yeast G19 strain supported the notion that OsHKT1;1-FL, but not OsHKT1;1-V2, mediates Na^+^ uptake (Figure 7). In addition to the growth phenotype, quantification of Na^+^ levels in yeast cells provided evidence that OsHKT1;1-FL but not OsHKT1;1-V2 led to overaccumulation of Na^+^ even under a control condition where the existence of 1 mM or less Na^+^ is expected (Figure 7C). This result confirms that the growth inhibition observed in OsHKT1;1-FL-expressing G19 strain is attributable to enhanced Na^+^ uptake. More importantly, the lack of remarkable Na^+^ accumulation in OsHKT1;1-V2-expressing cells is fully consistent with the electrophysiological analyses in Xenopus oocytes (Figure 3 and Figure 5).
During salt stress, maintenance of Cl^−^ homeostasis is crucial because excess cytosolic Cl^−^ can be toxic [41,42]. Cl^−^ detoxification involves xylem loading, vacuolar sequestration, and efflux from roots [41,43]. Chloride channels (CLCs) are membrane proteins that mediate Cl^−^ and NO_3_^−^ transport across plant membranes, contributing to pH regulation and stress tolerance [44,45]. Each CLC monomer contains 16–18 transmembrane α-helices that form a self-contained anion pore and a cytosolic CBS domain that regulates gating through conformational and nucleotide-dependent mechanisms [45,46]. The conserved GxGxPE and GKxGPxxH motifs form part of the selectivity filter, where serine or proline residues determine Cl^−^ versus NO_3_^−^ specificity [47]. Overexpression of GmCLC1, CsCLC-c, and ZmCLC-d has been reported to enhance salt tolerance by promoting vacuolar Cl^−^ sequestration and reducing cytosolic accumulation [48,49,50]. The plasma membrane transporters NPF2.4 and SLAH1 are proposed to function in regulating Cl^−^ loading into the xylem [51,52], whereas tonoplast-localized CLCs mediate Cl^−^/H^+^ antiport to sequester Cl^−^ into vacuoles [41,42]. Additionally, AtCCC1 functions as a Na^+^:K^+^:Cl^−^ cotransporter essential for maintaining ion balance and osmotic potential [53]. Loss-of-function mutants of AtCCC1 and OsCCC1 exhibit stunted growth and excessive Cl^−^ accumulation in shoots, indicating that CCC1 participates in Cl^−^ retrieval and homeostasis [53,54]. Moreover, some ALMT members (AtALMT9 and AtALMT12) facilitate Cl^−^ transport [55,56].
In the present study, we demonstrated that the OsHKT1;1-V2 splice variant encodes a protein of 172 amino acids with a single predicted transmembrane helix (Supplementary Figure S1A). Despite lacking the canonical pore-forming domains of HKTs, TEVC assays revealed robust inward and outward currents, suggesting that OsHKT1;1-V2 may form a self-associated complex that is permeable to Cl^−^. Sequence inspection of OsHKT1;1-V2 revealed a central hydrophobic stretch (residues 96–118; “LWVLILLMLMGGEVFTSMLGLYF”) (Supplementary Figure S1A), likely forming the ion-conducting α-helix, surrounded by charged and polar residues that could stabilize Cl^−^ within the pore. Notably, OsHKT1;1-V2 contains an upstream GKxGPxx pattern (residues 61–67) that resembles part of the conserved GKxGPxxH selectivity motif present in plant CLC proteins (Supplementary Figure S1A). Although this pattern lacks the canonical histidine present in CLC selectivity filters, it may still contribute to local structural flexibility or pore shaping, providing a residual signature of the anion-interacting architecture. Interestingly, the overall minimal architecture of OsHKT1;1-V2 parallels the basic structural layout of inwardly rectifying K^+^ (Kir) channels, which are composed of two transmembrane helices (TM1 and TM2) connected by a pore-forming loop and assembled as a tetramer to form a functional channel [57]. Kir channels exhibit inward rectification through pore blockage by intracellular Mg^2+^ and polyamines, allowing greater ion influx than efflux [57]. By analogy, OsHKT1;1-V2 might represent a minimal anion-conducting module, in which its single transmembrane helix or multimeric assembly provides a rudimentary Cl^−^ pathway.
Although Xenopus oocytes have endogenous anion channels, the following facts minimize the possibility that these channels contributed to the OsHKT1;1-V2-associated currents. First, in contrast to the Cl^−^-dependent reversal potential changes seen in OsHKT1;1-V2-expressing oocytes (Figure 4C,D, Supplementary Figure S6C,D), water-injected controls showed only small inward and outward currents that did not change with external Cl^−^ concentration (Supplementary Figure S4B). Second, Cl^−^ accumulation assays demonstrated that water-injected oocytes consistently showed minimal Cl^−^ accumulation under all tested conditions, and OsHKT1;1-V2-expressing oocytes showed an increase in Cl^−^ accumulation (Figure 6). Note, however, that we cannot exclude the possibility that the OsHKT1;1-V2 variant interacts with other membrane proteins to form a minimal Cl^−^-selective channel in oocytes. Thus, further mechanistic research will be needed. Approaches such as expressing dominant-negative OsHKT1;1-V2 mutants, testing OsHKT1;1-V2 multimerization biochemically, and reconstituting purified OsHKT1;1-V2 into liposomes or artificial membranes would clarify whether OsHKT1;1-V2 exhibits an inherently channel-forming feature by itself.
Although some amino acid substitutions were detected in the variants of OsHKT1;1 between Pokkali and Nipponbare (Supplementary Figure S5), the lineup of OsHKT1;1 transcripts was identical between the two cultivars [8]. In addition, OsHKT1;1-FL and other OsHKT1;1 variants derived from Nipponbare exhibited transport profiles similar to those of OsHKT1;1 transporters from Pokkali (Figure 8) [8]. Moreover, the similarity in the substrate selectivity of the OsHKT1;1-V2 variants from both rice varieties suggested that the function of OsHKT1;1-V2 in ion homeostasis might be conserved across different genetic backgrounds (Figure 4 and Figure 5, Supplementary Figure S6).
It has been reported that the transcript levels of OsHKT1;1 in shoots increased significantly, showing a 3- to 5-fold rise after 12 h of salt treatment, whereas no upregulation was observed in roots [22]. In contrast, HvHKT1;1 was upregulated in roots under salinity stress, with relatively lower expression levels detected in leaves and sheaths [41]. Similarly, the transcript levels of HKT1;1 have been analyzed in other plant species, in which the expression level was found to be increased in response to salt stress in both shoots and roots, or only in roots [58,59,60]. However, previous reports have only quantified full-length mRNA and have not considered variants. Absolute quantification of OsHKT1;1 transcripts in the present and previous studies demonstrated that the expression levels varied among variants and between the cultivars. Nipponbare exhibited higher and more stable expression of OsHKT1;1-FL under salt stress (Figure 9). As OsHKT1;1 has been proposed to be a salt-tolerant determinant in rice [22,23,24], it is intriguing that salt-tolerant Pokkali did not increase OsHKT1;1 transcripts and maintained a lower level of expression during salt stress compared with those in salt-sensitive Nipponbare (Figure 9). These results indicate that OsHKT1;1 may function to maintain basal ion homeostasis in Pokkali under normal condition and have a limited role in Pokkali under salt stress, although OsHKT1;1 in a salt-tolerant indica rice cultivar Zhenshan 2 was found to play a more essential role in the mechanism of salt tolerance [23]. In Pokkali, other specific transporters/channels, including OsHKT1;5 [2], may have significant functions under salt stress.
Overall, these results highlight the unexpected characteristics of the OsHKT1;1 splicing variant OsHKT1;1-V2 in rice, which was suggested to encode a possible Cl^−^ channel. However, the expression of OsHKT1;1-V2 was lower than that of OsHKT1;1-FL in both Pokkali and Nipponbare. However, important questions remain to be elucidated: (i) Does OsHKT1;1-V2 function as a Cl^−^ channel in rice? If so, (ii) does OsHKT1;1-V2 contribute to Cl^−^ homeostasis and salt tolerance? (iii) Does the differential regulation in the expression of OsHKT1;1-FL and OsHKT1;1-V2 between Nipponbare and Pokkali reflect the mechanism of salt tolerance in rice? In addition, further studies are needed to explore the regulatory mechanisms behind the splicing of OsHKT1;1 and to investigate whether some of the variants other than OsHKT1;1-V2 could also contribute to ion homeostasis in rice plants, including upon salt stress.
4. Materials and Methods
4.1. Plant Material and Growth Conditions
Seeds of Nipponbare (Oryza sativa L. ssp. japonica), a salt-sensitive rice cultivar, were sterilized, germinated, and grown hydroponically as described previously [8]. Total RNA was extracted from 14-day-old plants.
4.2. Extraction of RNA, and cDNA Synthesis
Total RNA was extracted, and its quality and integrity were assessed before synthesizing cDNA, following a previously described protocol [8]. The OsHKT1;1 cDNAs were then amplified using OsHKT1;1 cloning primers and subsequently inserted into a pCR4 topo vector (Invitrogen, Carlsbad, CA, USA).
4.3. Preparation of cRNA for X. laevis Oocytes Heterologous Expression
Full-length OsHKT1:1 (OsHKT1;1-FL) and a variant (OsHKT1;1-V2) cRNAs from Pokkali (Oryza sativa L. ssp. indica) have already been prepared [8]. The cDNAs of OsHKT1;1 from Nipponbare (Oryza sativa L. ssp. japonica) were cloned into a pXβG vector, and capped RNA (cRNA) synthesis was performed according to the procedure outlined by Imran et al. [8]. Oocytes were collected and injected with cRNA at concentrations of 50 ng/50 nL or 2.5 ng/50 nL, or with 50 nL of nuclease-free water as a negative control. The oocytes were then incubated at 18 °C, unless otherwise mentioned, in a modified Barth’s solution (MBS) until electrophysiological measurements were conducted as described [61].
4.4. Electrophysiology
Oocyte currents were measured using the two-electrode voltage clamp (TEVC) technique, conducted 1 d after cRNA injection, unless otherwise mentioned. TEVC recordings and subsequent data analysis were performed using an Axoclamp 900A amplifier (Molecular Devices, San Jose, CA, USA), Axon Instruments Digidata 1440A (Molecular Devices, San Jose, CA, USA), and pCLAMP 10 software (Molecular Devices, San Jose, CA, USA). The bath solutions were prepared with a baseline concentration of 1.8 mM MgCl_2_, 1.8 mM CaCl_2_, and 10 mM HEPES at pH 7.5 with Tris, and chloride salts of the monovalent cations were used. Gluconic acid magnesium (II) salt or D-mannitol was added as needed to maintain the osmolality of the external solutions between 200 and 220 mOsm in all measurements. To obtain current–voltage relationships, voltage steps (2 s) were applied from +30 to −120 mV in 15 mV decrements unless otherwise mentioned.
4.5. Determination of Cl− Content in X. laevis Oocytes
Oocytes were injected with 50 nL of water or cRNA of OsHKT1;1-FL (2.5 ng) and OsHKT1;1-V2 (2.5 ng and 25 ng), then incubated in 1× MBS medium for 24 h (OsHKT1;1-FL) or 4 h (OsHKT1;1-V2) at 18 °C. For the measurement of Cl^−^ accumulation, the oocytes were transferred to 1 mL of Na-gluconate or choline-Cl solutions. The Na-gluconate solution contained 96 mM Na gluconate, 1.8 mM Ca gluconate, 1.8 mM Mg gluconate, 1.8 mM mannitol, and 10 mM HEPES (pH 7.5 with Tris). The choline-Cl solution contained 96 mM choline Cl, 1.8 mM Ca gluconate, 1.8 mM Mg gluconate, 1.8 mM mannitol, and 10 mM HEPES (pH 7.5 with Tris). After a 6 h incubation at room temperature, the oocytes were washed three times with isotonic Ca-gluconate solution and then placed in a 1.5 mL tube to homogenize in Milli-Q water. Cell debris was removed by centrifugation. The supernatant was collected, diluted, and used for ion analysis using an anion chromatography system.
4.6. Subcellular Localization Analysis Using Arabidopsis Protoplasts
Subcellular localization analysis of OsHKT1;1-FL and OsHKT1;1-V2 proteins was conducted in Arabidopsis leaf protoplasts [35], which were transfected with the modified pTH2 vector [62] containing OsHKT1;1-FL or OsHKT1;1-V2 fused with GFP at the N-terminus under the control of the CaMV 35S promoter. Transfection was performed using the polyethylene glycol (PEG 4000) method, as described by Yoo et al. [63] and Sasaki et al. [64]. Protoplasts suspended in the re-suspension solution (0.5 M mannitol, 20 mM KCl, and 4 mM MES, pH 5.7) were incubated in the dark at 24 °C for 16 h [65]. The plasma membrane was visualized using FM4-64 (16 μM, 5 min). Images of the protoplasts were captured using an Olympus FluoView 1000 Confocal Microscope (Olympus Corporation, Hachioji, Tokyo, Japan). GFP was detected using 473 nm excitation with a 490–525 nm bandpass filter, whereas FM4-64 and chlorophyll were observed with 543 nm excitation and a 560–620 nm bandpass filter.
4.7. Subcellular Localization in Oocytes by Immunocytochemistry
Subcellular localization in oocytes was analyzed using OsHKT1;1-FL and OsHKT1;1-V2 with a FLAG tag (DYKDDDDK) at their N-termini. cRNAs were synthesized using an oocyte expression vector, pXβG vector, including OsHKT1;1-FL or OsHKT1;1-V2 with the sequence of FLAG tag, according to the procedure outlined by Imran et al. [8]. Immunohistochemical analysis was performed as described by Shibasaka et al. [66]. A volume of 50 ng/50 nl for OsHKT1;1-FL and 2.5 ng/50 nl for OsHKT1;1-V2 cRNAs or RNase-free water as a negative control was injected into the oocyte and incubated at 18 °C for 24 h (OsHKT1;1-FL) or 4 h (OsHKT1;1-V2). Subsequently, the samples were fixed for 3 h in 4% (w/v) formaldehyde solution (pH 7.4) and embedded in 5% agarose. Sections of 100 μm thickness were prepared using a micro slicer (Doshin EM, Osaka, Japan) and subjected to blocking in a buffer containing 50 mM Tris (pH 8.0), 150 mM NaCl, 0.1% Tween 20, and 3% BSA for 1 h at 25 °C. Immunolabeling was performed using primary antibodies, including rat anti-FLAG (raised against the synthetic peptide CDYKDDDDK, Invitrogen, Carlsbad, CA, USA), applied overnight at 4 °C. After three washes, the samples were incubated for another hour with a secondary antibody (Alexa Fluor 488-conjugated goat anti-rabbit IgG, Invitrogen, Carlsbad, CA, USA). After two washes with TBS-T (50 mM Tris, pH 8.0, 150 mM NaCl, 0.1% Tween 20) and a final wash with TBS (50 mM Tris, pH 8.0, 150 mM NaCl), fluorescent signals were observed using a fluorescence microscope (BZ-X700, Keyence, Osaka, Japan).
4.8. Functional Characterization of OsHKT1;1-FL and OsHKT1;1-V2 in Yeast
The CDSs of OsHKT1;1-FL and OsHKT1;1-V2 were cloned into the pYES2 vector and transformed into the well-known Na^+^-sensitive yeast strain G19, which lacks Na^+^ efflux transporters ENA1-4 [67,68]. Positive transformants were selected on Ura-selective medium plates [0.67% (w/v) yeast nitrogen base without amino acids, 0.077% (w/v) dropout mix-Ura, 2% (w/v) glucose, and 2% (w/v) agar]. For the yeast growth test, all transformed yeasts were cultured overnight at 30 °C in SC-Ura medium until the OD_600_ reached 1.0, and 10-fold serially diluted cultures were incubated on Ura-induced medium [0.67% (w/v) yeast nitrogen base without amino acids, 0.077% (w/v) dropout mix-Ura, 2% (w/v) galactose, 1% (w/v) raffinose, and 2% (w/v) agar] plates containing 50 mM NaCl. The plates were incubated at 30 °C for 4 days, and yeast cell growth was recorded.
4.9. Determination of Ion Contents in Yeast Cells
To quantify intracellular Na^+^, each transgenic G19 line was grown in liquid synthetic complete (SC) medium supplemented with or without 50 mM NaCl and galactose at 30 °C. Prior to the main culture, each transformant was cultured in a glass test tube filled with 5 mL SC-Ura glucose medium for overnight at 30 °C. 0.1 mL of each full growth-culture solution was inoculated in a 300 mL Erlenmeyer flask filled with 100 mL SC-Ura galactose medium. After incubation for 18 h (control) and 21 h (NaCl) at 30 °C at 130 rpm, each line was collected by swing-bucket centrifugation using 50 mL centrifuge tubes and then washed by 100 mL of sterilized ultrapure water. After 3 time washes, 10 mL of the washed culture solution was devided in a 15 mL centrifuge tube, and then samples were collected by swing-bucket centrifugation. All harvested samples were dried at 60 °C for 2 days. The dried samples were weighed and digested by 1 mL of ultrapure nitric acid for 24 h. Digests were subsequently heated at 95 °C for 15 min, which was repeated two times. Sodium (Na^+^) concentrations were quantified by inductively coupled plasma–mass spectrometry (ICP-MS).
4.10. Expression Analysis
Total RNA extraction, first-strand cDNA synthesis, and absolute quantification of gene expression using specific cDNAs standards were performed as previously described [8]. Total RNA was treated with DNase to remove contaminating genomic DNA (gDNA).
4.11. Statistical Analyses
Statistical analyses were performed using IBM SPSS Statistics (version 25). Significant differences were identified using a t-test (*** p < 0.001) for Figure 7 and a one-way ANOVA, and Tukey’s HSD test (p < 0.05) for Figure 6.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Munns R. Tester M. Mechanisms of salinity tolerance Annu. Rev. Plant Biol.20085965168110.1146/annurev.arplant.59.032607.09291118444910 · doi ↗ · pubmed ↗
- 2Singh R.K. Redoña E. Refuerzo L. Varietal Improvement for Abiotic Stress Tolerance in Crop Plants: Special Reference to Salinity in Rice Abiotic Stress Adaptation in Plants Pareek A. Sopory S. Bohnert H. Springer Dordrecht, Germany 200938741510.1007/978-90-481-3112-9_18 · doi ↗
- 3Rus A. Yokoi S. Sharkhuu A. Reddy M. Lee B.H. Matsumoto T.K. Koiwa H. Zhu J.K. Bressan R.A. Hasegawa P.M. At HKT 1 is a salt tolerance determinant that controls Na+ entry into plant roots Proc. Natl. Acad. Sci. USA 200198141501415510.1073/pnas.24150179811698666 PMC 61183 · doi ↗ · pubmed ↗
- 4Mäser P. Eckelman B. Vaidyanathan R. Horie T. Fairbairn D.J. Kubo M. Yamagami M. Yamaguchi K. Nishimura M. Uozumi N. Altered shoot/root Na+ distribution and bifurcating salt sensitivity in Arabidopsis by genetic disruption of the Na+ transporter At HKT 1FEBS Lett.200253115716110.1016/S 0014-5793(02)03488-912417304 · doi ↗ · pubmed ↗
- 5Rubio F. Gassmann W. Schroeder J.I. Sodium-driven potassium uptake by the plant potassium transporter HKT 1 and mutations conferring salt tolerance Science 19952701660166310.1126/science.270.5242.16607502075 · doi ↗ · pubmed ↗
- 6Hauser F. Horie T. A conserved primary salt tolerance mechanism mediated by HKT transporters: A mechanism for sodium exclusion and maintenance of high K+/Na+ ratio in leaves during salinity stress Plant Cell Environ.20103355256510.1111/j.1365-3040.2009.02056.x 19895406 · doi ↗ · pubmed ↗
- 7Xue S. Yao X. Luo W. Jha D. Tester M. Horie T. Schroeder J.I. At HKT 1;1 Mediates Nernstian Sodium Channel Transport Properties in Arabidopsis Root Stellar Cells P Lo S ONE 20116 e 2472510.1371/journal.pone.002472521931830 PMC 3170383 · doi ↗ · pubmed ↗
- 8Imran S. Horie T. Katsuhara M. Expression and ion transport activity of rice Os HKT 1;1 variants Plants 202091610.3390/plants 9010016 PMC 702020431877741 · doi ↗ · pubmed ↗