Swine Enteric Coronaviruses: An Updated Overview of Epidemiology, Diagnosis, Prevention, and Control
Yassein M. Ibrahim, Can Liu, Yuandi Yu, Liu Yang, Qianlin Chen, Wenjie Ma, Gebremeskel Mamu Werid, Shaomei Li, Jie Luo, Shengbin Gao, Suhui Zhang, Lizhi Fu, Yue Wang

TL;DR
This paper reviews swine enteric coronaviruses, their impact on pig health and economics, and recent advances in diagnosis and vaccine development.
Contribution
The paper provides an updated overview of epidemiology, diagnostics, and vaccines for swine coronaviruses, emphasizing recent innovations and control strategies.
Findings
Swine enteric coronaviruses cause severe intestinal disease and high neonatal mortality in pigs.
New diagnostic tools and next-generation vaccines like mRNA-based and virus-like particle platforms are being developed.
Viral evolution and recombination limit vaccine efficacy, especially for PEDV genogroup II strains.
Abstract
Swine enteric coronaviruses are major causes of severe intestinal disease in pigs, resulting in high piglet mortality and significant global economic losses. These viruses spread rapidly between farms and evolve quickly, undermining vaccine effectiveness. This review outlines their epidemiology, emerging diagnostic tools, and available vaccines. Because some swine coronaviruses can infect other animal species and potentially humans, they also present public health concerns. Strengthening farm biosecurity, improving vaccine strategies, and enhancing genomic surveillance are essential to control these infections and safeguard both swine production and human health. Swine enteric coronaviruses (SECoVs), including transmissible gastroenteritis virus (TGEV), porcine epidemic diarrhea virus (PEDV), porcine deltacoronavirus (PDCoV), and swine acute diarrhea syndrome coronavirus (SADS-CoV),…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
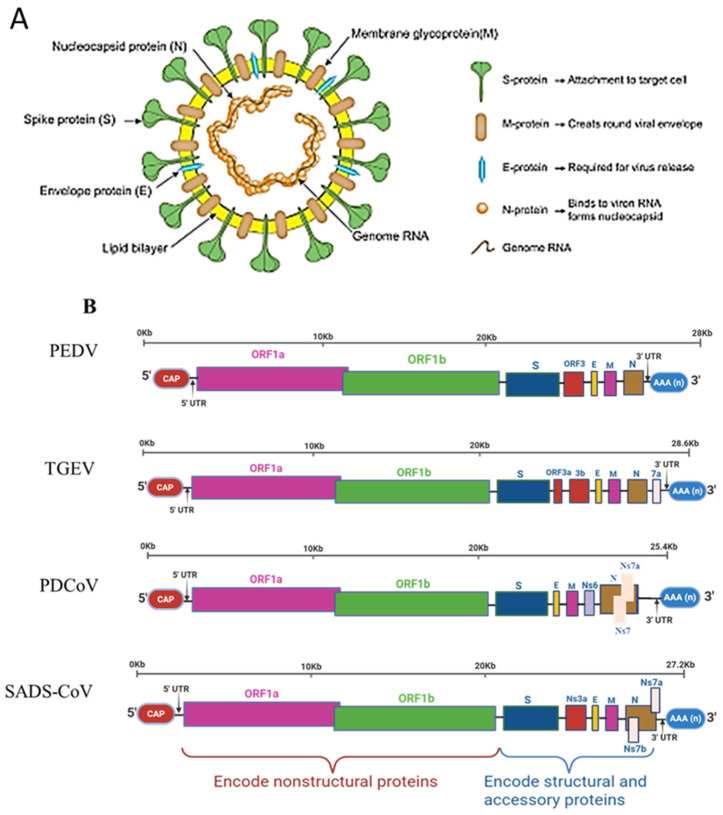 Figure 1
Figure 1| Genus | Virus | First Detection/Major Outbreaks | Key Prevalence Trends | Morbidity | Mortality of Piglets | Geographic Distribution | Notable Recent Findings | Reference |
|---|---|---|---|---|---|---|---|---|
| α-CoV | TGEV | 1946, USA; widespread mid-20th century | Now rare in Europe/USA; sporadic in Asia (i.e., China, Korea) | Less than 3% | Up to 100% in neonates | Declining globally, persists sporadically in Asia | Decline linked to PRCV cross-protection; a low-pathogenic mutant providing partial immunity | [ |
| α-CoV | PEDV | 1971, UK; 2010 China resurgence (>1 M piglets’ death); 2013 U.S. epidemic (>8 M piglets lost) | Persistent high prevalence in Asia (6–90%); lower but recurrent in Europe/North America | 80–100% | 50–100% | Asia, Europe, Americas; sporadic in Africa | G2 strains dominate globally; vaccine mismatch persists; recombinant strains reported | [ |
| δ-CoV | PDCoV | 2012, Hong Kong; 2014, USA outbreaks | Prevalence generally lower (1–30%), localized peaks >60% (e.g., Thailand 2015) | 20–30% | 30–40% | Asia, Americas; not reported in Europe/Africa | Serology suggests broader exposure; China meta-analysis ~14% positivity; detection in children in Haiti 2021. | [ |
| α-CoV | SADS-CoV | 2017, Guangdong, China (~24,500 piglet deaths) | Low PCR detection (<5%); serology shows widespread exposure (60–80% in China) | About 10% | 90–100% in piglets <5 days | China; related viruses in Vietnam bats/pigs; 2024 | Strong bat origin evidence; re-emergence in 2019 & 2021; zoonotic potential | [ |
| Virus | Country | Period | No. of Samples | Sample Type | Positive Samples | Type of Test | Prevalence Rate | References |
|---|---|---|---|---|---|---|---|---|
| TGEV | Canada | 1990 | 305 | Serum | 22 | ELISA | 7.2% | [ |
| Belgium | 1990 | 160 | Serum | 12 | ELISA | 7.5% | [ | |
| USA | 2008–2016 | 29,397 | Fecal | 667 | qPCR | 2.3% | [ | |
| Hungary | 2015–2016 | 908 | Serum | 140 | IFA | 15.4% | [ | |
| Italy | 2016–2017 | 444 | Serum | 3 | ELISA | 0.67% | [ | |
| 2022 | 438 | Serum | 24 | ELISA | 5.5% | [ | ||
| Poland | 2021–2024 | 828 | Serum | 18 | ELISA | 2.2% | [ | |
| Spain | 2017–2018 | 215 | Fecal | 6 | PCR | 2.8% | [ | |
| Germany | 2000–2005 | 1194 | Serum | 19 | ELISA | 1.59% | [ | |
| Czech Republic | 1999–2005 | 134 | Serum | 1 | ELISA | 0.75% | [ | |
| Croatia | 2005–2009 | 556 | Serum | 2 | ELISA | 0.4% | [ | |
| Argentina | 2014–2017 | 87 | Serum | 3 | ELISA | 3.4% | [ | |
| Cuba | 2008 | 90 | Intestine | 9 | PCR | 10% | [ | |
| Japan | 2007–2010 | 2703 | Serum | 389 | ELISA | 14.4% | [ | |
| South Korea | 2010 | 1295 | Serum | 64 | ELISA | 4.9% | [ | |
| 2023 | 350 | Serum | 15 | ELISA | 4.3% | [ | ||
| China | 2012–2014 | 314 | Fecal | 6 | PCR | 1.9% | [ | |
| 2015 | 44 | Intestine | 8 | nPCR | 18.2% | [ | ||
| 2015–2016 | 114 | Fecal | 4 | qPCR | 3.5% | [ | ||
| 2012–2016 | 390 | Fecal | 6 | PCR | 1.5% | [ | ||
| 2015–2016 | 27 | Intestine | 5 | PCR | 18.5% | [ | ||
| 2016–2018 | 181 | Fecal | 1 | mPCR | 0.55% | [ | ||
| 2015–2018 | 543 | Fecal | 46 | PCR | 8.5% | [ | ||
| 2017–2018 | 672 | Fecal | 35 | qPCR | 5.2% | [ | ||
| 2012–2018 | 2987 | Fecal | 21 | PCR | 0.7% | [ | ||
| 2019–2021 | 176 | Fecal + Intestine | 12 | PCR | 6.8% | [ | ||
| 2020 | 300 | Serum | 162 | SiELISA | 54% | [ | ||
| 2021–2023 | 1791 | Fecal | 15 | qPCR | 0.84% | [ | ||
| 2022–2023 | 5483 | Fecal + Intestine | 11 | qPCR | 0.2% | [ | ||
| PEDV | Italy | 2007–2014 | 51 | Fecal | 38 | PCR | 74% | [ |
| 2016–2017 | 444 | Serum | 17 | ELISA | 3.8% | [ | ||
| 2022 | 438 | Serum | 65 | ELISA | 14.8% | [ | ||
| 2007–2014 | 51 | Fecal | 47 | ELISA | 92% | [ | ||
| Vietnam | 2011–2016 | 108 | intestine | 87 | PCR | 80.6% | [ | |
| Spain | 2017–2018 | 215 | Fecal | 5 | PCR | 2.3% | [ | |
| 2017–2019 | 106 | Fecal | 41 | PCR | 38.7% | [ | ||
| China | 2011–2012 | 288 | Fecal + Intestine | 267 | PCR | 92.7% | [ | |
| 2011–2012 | 577 | Intestine + Milk | 417 | PCR | 72.3% | [ | ||
| 2011–2021 | 149,869 | Fecal | 70,384 | PCR | 46.9% | [ | ||
| 2012–2014 | 314 | Fecal | 76 | PCR | 24.2% | [ | ||
| 2012–2015 | 356 | Fecal + Intestine | 231 | nPCR | 64.9% | [ | ||
| 2014–2015 | 129 | Intestine | 119 | PCR | 92.3% | [ | ||
| 2014–2018 | 645 | Fecal+ Intestine | 231 | PCR | 35.8% | [ | ||
| 2015–2016 | 252 | Fecal | 165 | PCR | 65.5% | [ | ||
| 2012–2016 | 390 | Fecal | 88 | PCR | 22.6% | [ | ||
| 2015–2016 | 114 | Fecal | 55 | qPCR | 48.2% | [ | ||
| 2015–2016 | 27 | Intestine | 25 | PCR | 92.6% | [ | ||
| 2015–2017 | 398 | Fecal | 78 | mPCR | 19.6% | [ | ||
| 2015–2018 | 543 | Intestine | 363 | PCR | 66.9% | [ | ||
| 2015–2019 | 575 | Fecal + Intestine | 297 | PCR | 51.7% | [ | ||
| 2016–2017 | 116 | Fecal + Intestine | 61 | PCR | 52.6% | [ | ||
| 2016–2017 | 170 | Fecal + Intestine | 133 | PCR | 78.2% | [ | ||
| 2016–2018 | 719 | Fecal | 267 | qPCR | 36.7% | [ | ||
| 2016–2018 | 181 | Fecal | 56 | mPCR | 30.9% | [ | ||
| 2012–2018 | 2987 | Intestine | 1712 | PCR | 57.3% | [ | ||
| 2017–2018 | 672 | Fecal | 128 | qPCR | 19.1% | [ | ||
| 2017–2019 | 634 | Fecal + Intestine | 204 | qPCR | 32.2% | [ | ||
| 2019–2020 | 413 | Fecal + Intestine | 257 | PCR | 62.2% | [ | ||
| 2019–2021 | 176 | Fecal + Intestine | 66 | PCR | 37.5% | [ | ||
| 2020 | 300 | Serum | 18 | SiELISA | 6% | [ | ||
| 2017–2021 | 882 | Fecal | 564 | qPCR | 63.9% | [ | ||
| 2017–2022 | 673 | Fecal + Intestine | 363 | PCR | 53.9% | [ | ||
| 2020–2022 | 548 | Fecal | 258 | PCR | 47.1% | [ | ||
| 2021–2022 | 112 | Intestine | 58 | qPCR | 51.8% | [ | ||
| 2021–2023 | 1791 | Fecal | 890 | qPCR | 49.7% | [ | ||
| 2022–2023 | 481 | Fecal | 341 | PCR | 71% | [ | ||
| 2022–2023 | 5483 | Fecal + Intestine | 485 | qPCR | 8.8% | [ | ||
| India | 2022–2023 | 339 | Serum | 18 | ELISA | 5.3% | [ | |
| Japan | 2013–2014 | 1269 | Fecal | 248 | PCR | 19.5% | [ | |
| 2013–2014 | 204 | Fecal | 148 | PCR | 72.5% | [ | ||
| South Korea | 2013–2022 | 1131 | Fecal + Intestine | 140 | PCR | 12.4% | [ | |
| Mexico | 2016–2018 | 68 | Fecal | 53 | qPCR | 77.9% | [ | |
| 2019–2021 | 1054 | Serum | 650 | ELISA | 61.7% | [ | ||
| 2021 | 74 | Swab | 50 | PCR | 67.6% | [ | ||
| Vietnam | 2019 | 600 | Serum | 533 | ELISA | 88.8% | [ | |
| Thailand, Vietnam, Philippines and Lao PDR | 2015 | 97 | Fecal | 82 | PCR | 84.5% | [ | |
| Philippines | 2014–2015 | 34 | Fecal | 10 | PCR | 29.4% | [ | |
| Croatia | 2016 | 397 | Serum | 62 | ELISA | 15.6% | [ | |
| PDCoV | South Korea | 2014–2016 | 683 | Fecal | 130 | nPCR | 19.0% | [ |
| USA | 2014 | 293 | Fecal | 89 | qPCR | 30.4% | [ | |
| 2014 | 435 | Fecal | 109 | qPCR | 25% | [ | ||
| Japan | 2013–2014 | 477 | Fecal | 72 | qPCR | 15.1% | [ | |
| Vietnam | 2011–2016 | 108 | Intestine | 11 | PCR | 10.2% | [ | |
| Mexico | 2014–2017 | 885 | Swab | 85 | mPCR | 9.6% | [ | |
| Thailand | 2015 | 30 | Different | 26 | PCR | 86.7% | [ | |
| Thailand, Vietnam, Philippines and Lao PDR | 2015 | 97 | Fecal | 12 | PCR | 12.4% | [ | |
| China | 2014–2015 | 192 | Fecal | 73 | LAMP | 38% | [ | |
| 2015–2016 | 252 | Fecal | 55 | PCR | 21.8% | [ | ||
| 2015–2016 | 871 | Serum | 96 | ELISA | 11% | [ | ||
| 2012–2015 | 356 | Fecal + Intestine | 120 | nPCR | 33.7% | [ | ||
| 2012–2016 | 390 | Fecal | 5 | PCR | 1.3% | [ | ||
| 2015–2017 | 430 | Fecal | 101 | PCR | 23.5% | [ | ||
| 2015–2017 | 398 | Fecal | 144 | mPCR | 36.2% | [ | ||
| 2015–2018 | 543 | Fecal | 26 | PCR | 4.8% | [ | ||
| 2016–2017 | 170 | Fecal + Intestine | 15 | PCR | 8.8% | [ | ||
| 2012–2018 | 2987 | Fecal | 813 | PCR | 27.2% | [ | ||
| 2016–2018 | 719 | Fecal | 94 | qPCR | 13.1% | [ | ||
| 2016–2018 | 181 | Fecal | 32 | mPCR | 17.7% | [ | ||
| 2017–2018 | 672 | Fecal | 26 | qPCR | 3.87% | [ | ||
| 2017–2019 | 177 | Fecal + Intestine | 123 | PCR | 69.5% | [ | ||
| 2017–2019 | 634 | Fecal + Intestine | 84 | qPCR | 13.3% | [ | ||
| 2020 | 300 | Serum | 196 | SiELISA | 65.3% | [ | ||
| 2020–2023 | 4897 | Fecal | 362 | mqPCR | 7.4% | [ | ||
| 2021–2022 | 112 | Intestine | 3 | qPCR | 2.7% | [ | ||
| 2021–2023 | 1791 | Fecal | 307 | qPCR | 14.1% | [ | ||
| 2021–2023 | 481 | Fecal | 10 | PCR | 2.1% | [ | ||
| 2022–2023 | 5483 | Fecal + Intestine | 67 | qPCR | 1.2% | [ | ||
| SADS-CoV | China | 2012–2018 | 2987 | Fecal | 7 | PCR | 0.23% | [ |
| 2016–2017 | 170 | Fecal + Intestine | 74 | PCR | 43.5% | [ | ||
| 2016–2018 | 181 | Fecal | 21 | mPCR | 11.6% | [ | ||
| 2020 | 300 | Serum | 245 | SiELISA | 81.7% | [ | ||
| 2021–2023 | 1791 | Fecal | 40 | qPCR | 2.2% | [ | ||
| 2022–2023 | 12,978 | Serum | 7783 | ELISA | 59.9% | [ | ||
| 2024 | 72 | Fecal + Intestine | 11 | qPCR | 15.3% | [ | ||
| Vietnam | 2018–2023 | 69 | Fecal | 5 | PCR | 7.2% | [ |
| Period | Country | Pathogens | Samples No. | Coinfection Rate | Host | Reference |
|---|---|---|---|---|---|---|
| 2021–2023 | China | PEDV/PDCoV | 1791 | 41(2.29) | Swine | [ |
| PEDV/TGEV | 5 (0.28) | |||||
| PEDV/SADS-CoV | 4 (0.22) | |||||
| 2018–2021 | China | PEDV/PDCoV | 4468 | 1.50% | Swine | [ |
| PEDV/TGEV | 1.17% | |||||
| 2016–2018 | China | PDCoV/PEDV | 719 | 34 (4.73%) | Swine | [ |
| 2016–2018 | China | PEDV/PDCoV | 181 | 17 (9.4%) | Swine | [ |
| PEDV/SADS-CoV | 13 (7.2%) | |||||
| 2015–2018 | China | PDCoV/PEDV | 430 | 61(14.2%) | [ | |
| PDCoV/TGEV | 6 (5.9%) | |||||
| 2015–2018 | China | PDCoV/PEDV | (4.96%) | Swine | [ | |
| 2013–2018 | China | PDCoV/PEDV | 1547 | 17 (1.1%) | Swine | [ |
| 2012–2018 | China | PDCoV/PEDV | 2987 | 380 (12.72%) | Swine | [ |
| PEDV/TGEV | 9 (0.3%) | |||||
| PEDV/SADS-CoV | 3 (0.1%) | |||||
| PDCoV/TGEV | 3 (0.1%) | |||||
| 2015–2016 | China | PDCoV/PEDV | 252 | 2 (3.6%) | Swine | [ |
| 2015–2016 | China | PEDV/TGEV | 114 | 3 (2.6%) | Swine | [ |
| 2015–2016 | China | PEDV/TGEV | 27 | 3 (11.1%) | Swine | [ |
| 2016–2017 | China | PDCoV/PEDV | 3 (1.59%) | Tibetan pigs | [ | |
| 2016–2017 | China | PEDV/SADS-CoV | 170 | 30 (17.7%) | Swine | [ |
| 2017–2019 | China | PEDV/PDCoV | 634 | 47 (7.4%) | Swine | [ |
| 2016–2017 | China | PDCoV/PEDV | 172 | 16 (9.3%) | Swine | [ |
| 2017 | China | SAD-CoV/PEDV | 170 | 30 (17.65%) | Swine | [ |
| 2014–2017 | Mexico | PDCoV/PEDV | 885 | 46 (5.2%) | Swine | [ |
| 2014–2016 | South Korea | PDCoV/PEDV | 683 | 43 (6.3%) | Swine | [ |
| 2012–2016 | China | PDCoV/PEDV | 420 | 25 (5.95%) | Swine | [ |
| 2012–2016 | China | PDCoV/PEDV | 390 | 5 (1.28) | Swine | [ |
| 2011–2016 | Vietnam | PDCoV/PEDV | 108 | 11 (10.19%) | Swine | [ |
| 2014 | USA | PDCoV/PEDV | 435 | 19 (4.37%) | Swine | [ |
| 2014 | USA | PDCoV/PEDV | 293 | 29 (9.9%) | Swine | [ |
| 2015 | Thailand, Vietnam, Philippines and Lao PDR | PDCoV/PEDV | 97 | 29 (29.9%) | Swine | [ |
| 2012–2015 | China | PDCoV/PEDV | 356 | 70 (19.66%) | Swine | [ |
| Method | Examples | Advantages | Limitations | Applications | References |
|---|---|---|---|---|---|
| Traditional | Virus isolation, IF, IHC, electron microscopy | Visual confirmation; gold standard for lesions | Slow, labor-intensive, not field-suitable | Confirmation of pathology, research | [ |
| Conventional PCR | RT-PCR, nested RT-PCR | Specific, reliable | Time-consuming; contamination risk | Routine lab detection | [ |
| qPCR/multiplex RT-PCR | RT-qPCR, multiplex RT-PCR | High sensitivity; detects co-infections; quantification | Requires expensive equipment | Surveillance, clinical diagnosis | [ |
| Advanced PCR | ddPCR, DPO-PCR, NanoPCR | Ultra-sensitive; quantifies low-load samples | High cost; technical complexity | Precision diagnostics; mixed infections | [ |
| Isothermal amplification | RAA, RPA, LAMP | Rapid, low-equipment; field-friendly | Limited validation; risk of false positives | Point-of-care screening | [ |
| CRISPR-based detection | Cas12a, Cas13a | Rapid, highly sensitive; multiplexing | Cost; requires optimization | Future field diagnostics; rapid outbreak detection | [ |
| Serology | ELISA, phage-ELISA, recombinant ELISA | Herd monitoring; vaccine evaluation | Not suitable for early piglet infection | Serosurveillance, vaccine studies | [ |
| Rapid antigen/antibody tests | Lateral-flow strips | Simple, immediate | Lower sensitivity | Farm-level rapid screening | [ |
| Sequencing | NGS, amplicon sequencing | Detects variants; surveils evolution | Expensive; limited routine use | Genomic surveillance, emerging strain detection | [ |
| Virus Target | Vaccine Type | Expression System/Platform | Antigen | Adjuvant | Animals Model | Vaccine Efficacy | Limitations | Route of Administration | Reference |
|---|---|---|---|---|---|---|---|---|---|
| TGEV | Subunit |
| S protein epitopes (in silico predicted) | – | In silico | Predicted non-toxic, highly immunogenic, stable, and cost-effective epitopes | Experimental validation required (in vitro/in vivo) | – | [ |
| TGEV | Subunit | S protein epitopes (in silico) | – | In silico | Predicted immunogenic epitopes | Requires experimental verification | – | [ | |
| PEDV | Live attenuated | Serial in vitro passages | ORF3-deleted GIIb strain | – | Piglets | Immunogenicity and protective efficacy in pigs | Effects of mutations on pathogenicity require further investigation | IM | [ |
| PEDV | Live attenuated (chimeric construction) | Reverse genetic | YN144 (G2) | - | piglets | Broad-spectrum: Neutralizes both G1 and G2 strains. | Preclinical | PO | [ |
| PEDV | Live attenuated backbone | Reverse genetic modification of polymerase/ | PC22A (G2b) | - | piglets | Prevents recombination between vaccine and wild- | Preclinical | PO | [ |
| PEDV | Live attenuated CRISPR/Cas9 | Replacement of TRS-S with TRS- | AJ1102/G2 | - | piglets | Enhanced viral gene expression and antigen | Preclinical | PO | [ |
| PEDV | Inactivated | Formaldehyde inactivation | PEDV ShXXY2-2023 | ISA 201 | Piglets | Induced neutralizing antibodies and protective immunity; virus-neutralizing activity in vitro | Additional animal studies required | IM | [ |
| PEDV | Subunit (plant-based) | Plant expression system | CO-26K-equivalent epitope (COE) | Water-in-oil | Piglets | High PEDV-specific IgG, IgA, neutralizing antibodies, and IFN-γ responses | Optimization of dose and adjuvant; shedding studies required | IM | [ |
| PEDV | Subunit | Insect cells and silkworm | S protein trimer | Montanide IMS 1313 | Mice | Immunogenicity in mice and virus-neutralizing activity of mouse sera in vitro | No pig challenge model | IM | [ |
| PEDV | Subunit | HEK 293F cells | S protein trimer, S1 subunit, COE | M103 or M401 | Mice, piglets | Immunogenicity in mice and pigs, | IM | [ | |
| PEDV | Subunit | HEK-293F cells | S protein trimer (GIIa and GIIb) | M103 | piglets | Immunogenicity and protective efficacy in pigs and virus-neutralizing activity of swine sera in vitro | Preclinical | IM | [ |
| PEDV | Subunit | Trimer-Tag technology subcloned into the pPink-HC expression vector and expressed in yeast | S1 subunit trimer, COE trimer, RBD trimer | IAS 201 | Mice, sows | Immunogenicity in mice and pigs, | Preclinical | IM | [ |
| PEDV | Subunit | CSFV vector expressing S1 | S1 protein | – | Mice | High neutralizing antibody titers, IgG1/IgG2a, and IL-4 induction | No pig challenge model | IM | [ |
| PEDV | Subunit | CHO cells | Spike ectodomain | Montanide, IAS 201 VG | piglets | Immunogenicity and protective efficacy in pigs and virus-neutralizing activity of swine sera in vitro | Preclinical | IM | [ |
| PEDV | Bacterial vector |
| S1 glycoprotein | – | Mice | Strong humoral and cellular immune responses after oral administration | Protective efficacy in pigs not evaluated | PO | [ |
| PEDV | Bacterial vector |
| S1 and S2 proteins | – | Mice, sows | Induced systemic and mucosal immune responses in mice and pregnant sows | Challenge studies lacking | PO | [ |
| PEDV | Bacterial vector | S1 protein | – | Mice, piglets | Immunogenic and protective efficacy in pigs | Limited field validation | PO | [ | |
| PEDV | Bacterial vector |
| PEDV S + Brucella OMP16 | Freund’s complete adjuvant | Mice | Elevated IgG, neutralizing antibodies, IL-4, IL-10, IFN-γ | No pig challenge | PO | [ |
| PEDV | VLP | Insect cells | S, M, E proteins | Freund’s adjuvant + CCL25/CCL28 | Pigs | Protective immunity and neutralizing antibodies in pigs | Preclinical | IM | [ |
| PEDV | VLP | HEK293T cells | S, M, E proteins | Alum | Mice | Immunogenic with virus-neutralizing antibodies | Protective efficacy not evaluated | IM | [ |
| PEDV/TGEV | Chimeric VLP (ADDomer) | High Five cells | TGEV S1 + PEDV S1/S2 epitopes | ISA 201VG | Piglets | Immunogenic with neutralizing antibodies | Preclinical | IM | [ |
| PEDV | Viral vector (Adenovirus 5) | HEK293 cells | S and S1 (GIIa/GIIb) | – | Mice | Immunogenic and neutralizing antibodies | No pig challenge | IM/PO | [ |
| PEDV | Viral vector (Adenovirus 5) | HEK293 cells | COE-GIIb | – | Mice | Immunogenic and neutralizing antibodies | No pig challenge | IM/IN | [ |
| PEDV | Viral vector (Adenovirus 5) | HEK293 cells | S-GIIb | – | Piglets | Immunogenic and protective efficacy in pigs | Preclinical | IM/IN | [ |
| PEDV | Viral vector (pseudorabies virus) | ST cells | S1 epitopes | – | Piglets | Protective immunity and neutralizing antibodies | Vector safety concerns | IM | [ |
| PEDV | Viral vector (BVDV) | – | S fragment (aa 499–602) | – | Mice | PEDV- and BVDV-specific IgG induction | No pig challenge | PO | [ |
| PEDV | Viral vector (CSFV) | – | S1 NCOE | – | Rabbits, piglets | Elevated PEDV antibodies and IFN-γ | Preclinical | IM/IN | [ |
| PEDV | Viral vector (PoRV) | Reverse genetics | ORF3 replaced with VP7 | – | Piglets | Dual PEDV and PoRV neutralizing antibodies | Preclinical | IM/PO | [ |
| PEDV | Viral vector (PRRSV) | – | S-G2 | – | Piglets | Dual immunity against PEDV and PRRSV | Safety and recombination concerns | IM | [ |
| PEDV | DNA vaccine | pPink-HC vector | S1-, COE-, and RBD-trimers | – | Mice, sows, piglets | Strong systemic and mucosal IgG/IgA responses | Not commercialized | IM | [ |
| PEDV | mRNA-LNP | In vitro transcription | S or SM | – | Mice, piglets | Protective immunity and neutralizing antibodies | Cold chain and cost constraints | SC/IM | [ |
| PDCoV | Inactivated | β-propiolactone | HNZK-02 strain | Al (OH)3 or ODN2395 | Mice | Strong humoral and cellular immunity | Low mucosal immunity vs. LAV | SC | [ |
| PDCoV | Inactivated vs. subunit | – | Whole virus vs. spike protein | – | Piglets, sows | Long-term antibodies and reduced shedding | Model-dependent | IM | [ |
| PDCoV | Live attenuated | NS6 deletion | USA/IL/2014/026 | – | Piglets | Reduced virulence, stable attenuation | Reversion risk | PO | [ |
| PDCoV | Live attenuated | Serial passage CZ2020 | CZ2020 | – | Piglets | Reduced virulence, improved gut microbiota | Variant-specific, preclinical | PO | [ |
| PDCoV | Live attenuated | Serial passage DHeB1 | DHeB1 | – | Piglets | Safe, reduced lesions and viral load | Field validation needed | IM | [ |
| PDCoV | VLP | HEK293F cells | S, M, E proteins | M103 | Mice, sows, piglets | Protective immunity and neutralizing antibodies | Adjuvant-dependent | IM/PO | [ |
| PDCoV | VLP | Insect cells | S, M, E proteins | Freund’s | Mice | Immunogenic | No pig challenge | IM | [ |
| PDCoV | Subunit | Insect cells | RBD | Al (OH)3, CpG2395, aqueous | Mice | Immunogenic | No pig challenge | IM | [ |
| PDCoV | Subunit |
| RBD dimer | Ferritin + ISA 201VG | Mice | Protective immunity and neutralizing antibodies | Preclinical | IM | [ |
| PDCoV | Subunit | Insect cells | S1 protein | Gel 01 | Mice, sows | Protective immunity | Preclinical | IM | [ |
| PDCoV | Subunit | CHO cells | S, N, M proteins | AddaVax | Mice, piglets | Immunogenic | Preclinical | IM | [ |
| PDCoV | Viral vector (PRV, CRISPR/Cas9) | – | S protein | – | Mice | Local and systemic immunity | Vector-related safety issues | IM | [ |
| PDCoV | Nanoparticle | S1-CTD | – | Mice, sows, piglets | Robust and prolonged protective immunity | Cost and scalability issues | IM | [ | |
| PDCoV | mRNA-LNP | In vitro transcription | S2P stabilized S | – | Piglets | Strong humoral and cellular immunity | Cold chain, limited trials | PO | [ |
| PDCoV | mRNA-LNP | In vitro transcription | S and S ectodomain | – | Mice, piglets | Protective immunity and neutralizing antibodies | Cold chain, limited trials | IM | [ |
| Compound | Target Virus | Testing Model | Antiviral Mechanism | Main Phases of Action | Reference |
|---|---|---|---|---|---|
| Rhodamine derivative LJ001 | TGEV | In vitro | Interferes with the replication stage of viral life cycle | Replication | [ |
| PDCoV | In vitro | Interferes with the replication stage of viral life cycle | Replication | [ | |
| Melatonin | TGEV | In vitro | Blocks viral entry | Replication | [ |
| PEDV | In vitro | Blocks viral entry | [ | ||
| PDCoV | In vitro | Blocks viral entry | [ | ||
| Indole | TGEV | In vitro | Blocks viral entry | Replication | [ |
| PEDV | In vitro | Blocks viral entry | [ | ||
| PDCoV | In vitro | Blocks viral entry | [ | ||
| Rifampicin | PEDV | In vitro | Inhibits viral replication | Replication | [ |
| SADS-CoV | In vitro | Inhibits viral replication | [ | ||
| Griffithsin | PEDV | In vitro | Inhibits viral attachment and disrupts cell-to-cell | Attachment | [ |
| PDCoV | In vitro | Inhibition | Attachment | [ | |
| Surfactin | TGEV | In vitro | Acts on viral lipids to inhibit fusion between viral and | Attachment, entry | [ |
| PEDV | In vitro and | Acts on viral lipids to inhibit fusion between viral and | [ | ||
| 25-hydroxycholesterol | TGEV | In vitro | Blocks viral entry | Attachment, entry | [ |
| PEDV | In vitro | Blocks viral entry | Attachment, entry | [ | |
| PDCoV | In vitro | Blocks viral entry | Attachment, entry | [ | |
| Ergosterol peroxide | PEDV | In vitro | Inactivates virions directly and inhibits viral | Entry, replication, | [ |
| PDCoV | In vitro and | Inhibits viral attachment, entry, and the early and | Attachment, entry, | [ | |
| Lithium chloride | TGEV | In vitro | Inhibited virus attachment | Entry, replication | [ |
| PEDV | In vitro | Inhibited virus attachment | Entry, replication | [ | |
| PDCoV | In vitro | Inhibited virus attachment | Entry, replication | [ | |
| Octyl gallate | PEDV | In vitro and | Interacts with PEDV 3CLpro and inhibits 3CLpro activity | Replication | [ |
- —National Center of Technology Innovation for Pigs
- —Chongqing Talent plan “contract system” project
- —Sichuan Provincial Regional Innovation Cooperation Project
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAnimal Virus Infections Studies · Viral gastroenteritis research and epidemiology · Animal Disease Management and Epidemiology
1. Introduction
Swine enteric coronaviruses (SECoVs), including porcine epidemic diarrhea virus (PEDV), transmissible gastroenteritis virus (TGEV), porcine deltacoronavirus (PDCoV), and swine acute diarrhea syndrome coronavirus (SADS-CoV), are highly contagious pathogens that threaten pig health and cause substantial economic losses in the global swine industry [1]. These viruses primarily infect intestinal epithelial cells, leading to villous atrophy and enterocyte loss. Clinically, infection manifests as severe enteritis characterized by anorexia, vomiting, watery diarrhea, dehydration, lethargy, metabolic acidosis, hyperkalemia, and high mortality in piglets [2,3]. Although clinically similar, SECoVs are antigenically distinct and confer no cross-protection [4,5]. Coronaviruses (CoVs) belong to the subfamily Orthocoronavirinae within the family Coronaviridae (order Nidovirales) [6], and are taxonomically divided into four genera: Alphacoronavirus (α-CoV), Betacoronavirus (β-CoV), Gammacoronavirus (γ-CoV), and Deltacoronavirus (δ-CoV). (https://ictv.global/report/chapter/coronaviridae/coronaviridae [ICTV, 2019], accessed on 22 November 2025). Gamma- and Deltacoronaviruses primarily infect birds, whereas alpha- and betacoronaviruses infect a wide range of mammals, including humans, livestock, rodents, bats, and other wild animals, causing diseases of the respiratory, digestive, hepatic, and nervous systems [7].
Coronavirus virions are spherical to pleomorphic, measuring 80–120 nm in diameter, and are characterized by trimeric spike (S) glycoproteins that form a distinctive crown-like structure under electron microscopy [6,7] (Figure 1A). The coronavirus genome consists of positive-sense, single-stranded RNA of 26–32 kb that encodes structural proteins (spike, envelope [E], membrane [M], and nucleocapsid [N]), non-structural proteins (nsp1–16), and multiple accessory proteins [6]. The accessory gene composition varies among SECoVs: TGEV encodes NS3a, NS3b, and NS7; PEDV encodes ORF3; PDCoV encodes NS6, NS7, and NS7a; and SADS-CoV encodes NS3, NS7a, and NS7b (Figure 1B). The S protein mediates viral entry, with the S1 subunit recognizing host cell receptors and determining tissue tropism, while S2 enables membrane fusion [6,8]. The S protein also harbors key epitopes essential for neutralization, vaccine design, diagnostics, and T-cell responses. The M, E, and N proteins support virion assembly, structural integrity, and host immune modulation, while non-structural proteins drive genome replication and evade antiviral defenses [8,9,10]. Accessory proteins, though dispensable for replication in vitro, contribute substantially to virulence and immune evasion in vivo [11].
SECoVs exhibit high mutation and recombination rates, particularly within the S gene, resulting in quasispecies diversity, which enables rapid adaptation, immune escape, and cross-species transmission [12,13]. Their genomic plasticity accelerates viral evolution and raises concerns regarding potential zoonotic spillover events [14]. Historical outbreaks demonstrate the devastating impact of SECoVs on global pig production: in 2010, China reported over one million piglet deaths [15]; in 2013, the United States experienced more than eight million fatalities with estimated economic losses of USD 900 million to 1.8 billion [16,17]; and in 2017, the first outbreak of SADS-CoV in China caused more than 20,000 piglet deaths [18].
Although several reviews on SECoVs have been published in recent years, most focus primarily on epidemiology, genomic variation, or phylogenetic classification, with limited integration of technological and translational advances. This review synthesizes recent progress (post-2020) in diagnostics, vaccine development, and next-generation sequencing-based genomic surveillance, bridging molecular evolution, epidemiology, and control strategies. It further offers a comparative synthesis of viral recombination and interspecies transmission, with particular emphasis on the emergence of recombinant and cross-species SECoV strains.
Ge. me organization and structure of SECoVs. (A) Schematic of porcine coronaviruses’ (PEDV, TGEV, PDCoV, and SADS-CoV) genome structure showing virions containing S, E, and M proteins in the lipid bilayer, enclosing the N protein–RNA complex (gray line) [19]. (B) Genome map showing structural proteins (S, E, M, N) and accessory genes (Ns3a, Ns6, Ns7, Ns7a, Ns7b), with a 5′ cap and 3′ poly(A) tail.
2. Epidemiology Occurrence and Transmission
2.1. Pathogenesis and Clinical Manifestations
TGEV has long circulated in pigs, whereas PEDV, PDCoV, and SADS-CoV are emerging or re-emerging coronaviruses that continue to threaten global pig health and production. SECoVs primarily replicate in the jejunum and ileum, where they induce interferon responses and apoptosis [20,21]. Infection leads to severe intestinal epithelial damage characterized by villous atrophy, mucosal thinning, and impaired nutrient absorption, resulting in diarrhea, dehydration, and weight loss [22,23].
Clinical signs are similar across SECoVs, appearing 1–3 days post-exposure, lasting up to 10 days, and often followed by extended viral shedding that enables recovered pigs to act as reservoirs [24]. Disease severity is strongly age-dependent [24]. Neonatal piglets are most susceptible: TGEV and PEDV can cause nearly 100% mortality, SADS-CoV is highly fatal (>90%) in piglets <5 days old, while PDCoV typically induces milder disease with 30–40% mortality in neonates [25]. Older pigs, especially sows, generally develop mild symptoms with low mortality. Clinical outcomes depend on host factors such as immature intestinal barriers and limited neonatal immunity, as well as viral factors including recombination and antigenic variation [25]. Co-infections, environmental stress, and high viral loads further aggravate disease severity [25,26]. Epidemiological patterns and mortality data are summarized in Table 1.
2.2. Historical Context and Global Distribution
TGEV was first identified in the United States in 1946 [40] and subsequently spread globally [41]. Until the 1980s, it remained a major cause of enteric disease despite vaccination. Currently, TGEV prevalence has markedly declined, largely due to the widespread circulation of porcine respiratory coronavirus (PRCV) [42], a naturally occurring TGEV variant carrying a 621–681 nt deletion in the spike gene that attenuates virulence and alters tropism from enteric to respiratory tissues. PRCV generally causes mild or subclinical respiratory infections but induces cross-neutralizing antibodies that confer partial protection against TGEV, reducing selective pressure for virulent enteric strains [43]. Despite lacking major antigenic sites, PRCV retains conserved epitopes that cause strong serological cross-reactivity with TGEV [28,44]. Consequently, PRCV-induced antibodies often yield false-positive results in standard TGEV ELISAs and neutralization assays, complicating serodiagnosis and obscuring true epidemiological patterns unless differential or molecular assays are used [43]. TGEV is classified into two genotypes: GI (including Purdue and Miller strains) and GII (comprising U.S. variants) [45,46], Figure 2A. Despite reduced prevalence, new virulent strains have emerged, including recombinant Purdue–Miller variants in China [47,48]. In Europe, a recombinant swine enteric coronavirus (SeCoV) derived from TGEV and PEDV was reported, possessing an S gene >90% identical to PEDV and the remaining genome up to 97% identical to virulent TGEV strains [49,50]. Additionally, virulent isolates such as JS2012, HQ2016, and SC2021 continue to appear sporadically in Asia and the Americas [48,51,52].
PEDV was first reported on British farms in 1971, and the CV777 strain identified in Belgium in 1978 became the classical PEDV reference [30]. During the 1970s–1980s, PEDV caused major outbreaks across Europe, leading to serious economic losses. From the late 1980s to 2010, the virus circulated mainly in Europe and Asia, causing sporadic or localized outbreaks [31]. A highly virulent variant emerged in China in 2010, spreading rapidly and causing catastrophic losses even in vaccinated herds, highlighting antigenic mismatch and vaccine failure [31]. In 2013, closely related strains introduced into the United States triggered an epidemic, reducing the national swine population by approximately 3%, with losses of USD 900 million to 1.8 billion [53]. By late 2016, genetically diverse PEDV lineages had become established across North America, Asia, and parts of Europe. Today, PEDV remains a persistent and re-emerging threat to swine health and production in most pig-producing regions worldwide [32,54].
Phylogenetic analysis defines two major genogroups: GI (classical) and GII (variant), comprising six subgroups (GIa, GIb, GIIa, GIIb, GIIc, GIId) [55,56,57,58], Figure 2B. GI includes early virulent European strains; GII contains variants that emerged around 2010 and now dominate globally, particularly in China and North America [56,59,60]. The recombination-driven GII lineage continues to evolve rapidly and remains poorly controlled by GI-based vaccines [33,60].
PDCoV was first detected in Hong Kong in 2012 [61]. Its pathogenic significance became evident in 2014 when it caused major outbreaks in Ohio that spread to >20 U.S. states [62,63]. Since then, PDCoV has been reported across North America, Asia, and South America [64,65], resulting in notable economic losses. PDCoV causes 30–40% mortality in neonatal piglets, increasing sharply during co-infections [29]. Phylogenetically, PDCoV comprises two genogroups: GI (China, America, Japan, Korea) and GII (Southeast Asia), including SEA-1 (Thailand), SEA-2 (Vietnam), and the recombinant SEA-2r strain [65], Figure 2C. Chinese isolates display the highest genetic diversity, positioning China as a principal evolutionary reservoir [65,66].
SADS-CoV was first identified in Guangdong, China, in 2017, causing outbreaks with approximately 90% case fatality rates in piglets <5 days old [18,37,67]. Whole-genome sequencing shows exceptional nucleotide identity (approximately 99.9%) among isolates from 2017 to 2023 [39,68], indicating minimal genetic diversification. Phylogenetic analyses show that all strains cluster with HKU2-like coronaviruses in bats, sharing 95–98% nucleotide identity [18,69], strongly supporting bat origin and likely a single spillover event followed by sustained pig-to-pig transmission [70]. The spike gene exhibits unexpected affinity to betacoronaviruses despite SADS-CoV being classified as an alphacoronavirus, a pattern also observed in some rodent α-CoVs [71], suggesting historical recombination or ancestral divergence between α- and β-CoVs [37,72,73], Figure 2D. Although currently confined to China, the detection of related viruses in Vietnamese bats and pig feces [68,74] indicates the potential for cross-border spread.
2.3. Transmission of SECoVs
Understanding SECoV transmission is essential for effective control. These viruses spread primarily via the fecal–oral route and indirectly through contaminated equipment, feed, personnel, and trailers [24,75,76,77]. PEDV and PDCoV can also transmit through aerosols [32,34], with PEDV RNA detected up to 10 km from infected farms [78,79]. PEDV RNA has been detected in the semen of infected boars with prolonged shedding in the sperm-rich fraction [80], indicating potential sexual transmission. Additionally, PEDV can be transmitted lactogenically; virus-bearing T cells migrate to the mammary gland and are secreted in colostrum and milk [81], providing a clinically significant route that sustains herd-level infections. Milk-borne viral shedding post-infection exposes entire litters at their most susceptible stage, bypassing biosecurity measures and potentially overwhelming maternal antibody protection [81]. Figure 3 summarizes the major transmission pathways of SECoV. The environmental resilience of SECoVs facilitates prolonged survival and sustained indirect transmission between farms.
2.4. Global Prevalence and Distribution
SECoV prevalence is associated with sampling season, region, year, pig stage, and clinical signs [82,83]. Geographic variation reflects differences in climate, production systems, biosecurity capacity, and surveillance intensity. Outbreaks are more frequent in winter and spring [1], when prolonged viral survival and increased indoor housing enhance transmission. Large commercial systems experience frequent detection via active surveillance, whereas smallholder regions report sporadic outbreaks due to limited diagnostic infrastructure [84]. International trade in pigs and feed, along with shared transport networks, facilitates cross-border transmission and the emergence of novel variants [85,86]. The rapid global spread of PDCoV after its 2014 emergence in the U.S. illustrates this transboundary risk [24]. Table 2 summarizes SECoV prevalence by country.
TGEV prevalence varies globally, but accurate estimation is hindered by extensive serological cross-reactivity with PRCV, which has circulated widely across Europe, North America, and Asia since the mid-1980s [43,48,51,52]. Studies lacking differential diagnostic methods often overestimate true TGEV exposure [28,87]. For example, the Canadian study reporting 7.2% seropositivity [88] did not differentiate PRCV, and concurrent circulation suggests some positives were PRCV-derived. Similarly, a 2020 Chinese survey reporting 54% seropositivity [89] likely reflected combined TGEV/PRCV antibody responses given PRCV’s endemic status [90]. In contrast, molecular surveillance using RT-PCR and S gene sequencing consistently reports very low TGEV detection (<1%), indicating that TGEV is now rare while PRCV remains widespread [91,92]. In the U.S., analysis of 29,397 samples (2008–2016) found 2.3% positivity overall, peaking at 6.8% in 2010 and declining to <0.1% by 2014 [42]. In Japan, TGEV caused eight outbreaks between 2001 and 2007, with 14.4% seroprevalence in 2010 but no detections after 2008 [93,94]. A 2023 South Korean study similarly found 4.3% seropositivity without active infection [95]. Overall, TGEV circulation appears sporadic. Seroprevalence data should be interpreted cautiously unless PRCV differentiation is confirmed, and future surveillance should combine molecular assays with validated differential serological tests.
PEDV spread widely through Europe and Asia in the 1990s [31,96], and re-emerged in China in 2010 [31], followed by the U.S. (2013) [97], Mexico (2013) [98], and Canada (2014) [99]. Subsequent outbreaks across South Korea, Thailand, Vietnam, and India [31,100], along with renewed activity in Europe [101,102,103,104,105,106,107], caused major losses. Piglet mortality reached approximately 30% in Hungary [108] and >70% in Germany [109]. In Croatia, mortality reached 20–30% in suckling piglets, with 82.8% seropositivity at the farm level [110]. In China, extensive surveillance (2011–2023) documented persistent PEDV circulation, with prevalence ranging from 6.0% to 92.7% depending on diagnostic method and sample type [32,33,55,92,111,112,113]. PEDV remains the dominant SECoV (>45% detection), requiring continuous surveillance and genotype-specific vaccines.
PDCoV prevalence from 2011 to 2023 varied substantially across Asia and the Americas. Moderate detection was reported in South Korea (19.0%) [114], Japan (15.1%) [64], Vietnam (10.2%) [115], and Mexico (9.6%) [116]. In the U.S., prevalence reached 25–30.4% in 2014 [117], with 244 outbreaks reported from 2015 to 2023 across 16 states as incidence increased from 0.44% to 4.28% [118]. A severe outbreak in Thailand in 2015 caused 27.6% mortality in sows and 64.3% in piglets, with 86.7% positivity [119]. In China, detection rates ranged from 1.2% to 69.5% depending on province and study design, while serological studies recorded 11–65% antibody positivity [91,92,120,121,122,123]. More recent nationwide surveys (2020–2023) consistently reported lower prevalence (1.2–14.1%) [92,123,124,125,126], suggesting declining circulation.
Molecular detection of SADS-CoV from 2012 to 2024 varied widely across studies. Large fecal surveys reported low molecular detection (0.23–2.2%) [91,92], whereas targeted sampling showed higher rates (15–43.5%) [127,128]. Serology revealed higher exposure (59.9–81.7%, 2020–2023) [89,129]. The detection of related viruses in Vietnamese bats and pigs (7.2%) [68,74] supports cross-border transmission.
SADS-CoV has been reported mainly in China and Vietnam, likely due to bat reservoir distribution, intensive pig production, and wildlife–livestock interfaces [18,67,69], with phylogenetic evidence indicating dissemination from China to Vietnam [68,130].
Under-surveillance, misdiagnosis, and biosecurity differences may explain the lack of reports elsewhere [131]. Therefore, expanded molecular and serological surveillance in regions with high bat diversity and intensive pig production is essential to clarify the global distribution and emergence risk of SADS-CoV.
2.5. Co-Infections Dynamics Underlying Conditions
Co-infection with enteric pathogens complicates SECoV control [148] by obscuring diagnosis and increasing morbidity and mortality in neonatal piglets [174,175]. Enterocytes in the jejunum and ileum are frequently co-infected by PEDV, PDCoV, TGEV, and SADS-CoV, creating ideal conditions for recombination and the emergence of novel variants [42,174,176]. SECoV co-infections are widely reported across Asia and the Americas, with common combinations including PEDV/PDCoV, PEDV/TGEV, and PEDV/SADS-CoV, as shown in Table 3.
PDCoV/PEDV co-infection reached 29.9% in Southeast Asia (2015) and 1.1–9.9% in South Korea, Mexico, and the USA [114,116,117,168]. In China (2012–2023), rates ranged from 0.1% to 19.66%, dominated by PEDV/SADS-CoV (17.7%) [127], and PDCoV/PEDV (19.66%) [120]. Moderate prevalence was observed for PEDV/TGEV (0.22–11.1%) [91,92,144,146,174] and PDCoV/TGEV (5.9%) [172]. Large-scale surveillance (2012–2018) reported PEDV/PDCoV co-infection in 12.72% of 2987 samples, while other combinations were <0.3% [91].
Co-infections exacerbate disease severity by impairing cytokine responses, damaging gut integrity, and enhancing susceptibility to secondary pathogens. For example, PEDV facilitates SADS-CoV infection [127]. Such interactions have likely contributed to the emergence of new SECoVs, including the recent chimeric TGEV-PEDV virus in Europe [61,62,101,177,178]. SECoVs co-circulate with porcine rotaviruses A and C, increasing disease severity and shedding [125,179] and highlighting the need for multiplex diagnostic assays.
2.6. Cross-Species Transmission and Zoonotic Risks
Coronaviruses exhibit high mutation rates (10^−3^ to 10^−5^ substitutions per site per replication cycle) and recombination frequencies despite the proofreading activity of nsp14 (ExoN) [182,183], generating quasispecies that enhance adaptability [12,13,184]. This enables rapid host adaptation, immune evasion, and cross-species transmission [185,186,187]. This genomic plasticity accelerates viral diversification and zoonotic potential. The animal origins of SARS-CoV, MERS-CoV, and SARS-CoV-2 [188], combined with widespread coronavirus circulation in birds and mammals and their rapid evolutionary dynamics [182,189,190], underscore the public health relevance of SECoVs, which share ancestral lineages with these zoonotic pathogens.
TGEV can infect wild and domestic carnivores (foxes, dogs, mink, cats) subclinically, enabling reservoir maintenance, although only canine-derived strains have been confirmed to infect pigs [49]. A novel canine coronavirus (CCoV-HuPn-2018) identified in Malaysian pneumonia patients, mainly children with animal exposure, showed high genetic similarity to canine coronaviruses and virulent TGEV [191]. Phylogenetic analysis clustered the virus with canine, feline, and porcine alphacoronaviruses, including TGEV and SeCoV [191], confirming close evolutionary relationships.
Evidence for PEDV cross-species transmission is scarce. The virus has been identified in wild pigs only in Korea and the United States, likely reflecting spillover from domestic herds [192,193]. However, PEDV can replicate in human, monkey, pig, and bat-derived cell cultures, supporting the hypothesis that it previously crossed species barriers, most plausibly from bats to pigs [194,195].
SADS-CoV exhibits broad host tropism, with efficient replication in cell lines derived from swine, poultry, cattle, felines, non-human primates, and humans [196,197,198,199,200,201,202,203,204,205], suggesting intrinsic viral determinants that facilitate cross-species infection. Accordingly, despite negative serological findings among occupationally exposed farm workers [18], subsequent experimental studies demonstrated significant zoonotic potential [204], as the virus replicates efficiently in primary human respiratory and intestinal cells via ACE2/DPP4/CD13-independent entry mechanisms [204].
PDCoV demonstrates the most robust cross-species transmission capacity among SECoVs. Experimental infections show productive replication and transmission in mice, calves, and poultry [39,201,206,207], while natural infections have been documented in wild birds, Asian leopard cats, and badgers [61]. In vitro studies reveal broad cellular tropism across multiple mammalian and avian lineages, including human intestinal epithelial cells [2,208]. Most significantly, three PDCoV strains were isolated from children presenting with acute febrile illness in rural Haiti [209], providing direct evidence of natural human infection in settings characterized by close livestock contact. These findings highlight the evolutionary adaptability of SECoVs and implicate swine as potential intermediate hosts for emerging zoonotic coronaviruses. Proactive One Health surveillance, integrating molecular monitoring in swine systems with seroepidemiological screening of exposed populations, is essential for early spillover detection and pandemic preparedness [210].
3. Diagnostic Approaches
SECoV diagnosis is challenging due to overlapping clinical signs, requiring reliable laboratory confirmation [25,211]. Traditional methods such as virus isolation, immunofluorescence (IF), immunohistochemistry (IHC), electron microscopy, and polymerase chain reaction (PCR) are informative but slow and unsuitable for field use [29]. Molecular diagnostics now serve as the primary tools for SECoV detection, offering higher sensitivity, faster turnaround, and improved capability to identify co-infections [212]. A detailed summary of available diagnostic platforms, including their advantages and limitations, is presented in Table 4.
PCR-based assays, including multiplex RT-PCR, RT-qPCR, nested RT-PCR, digital droplet PCR (ddPCR), and nanoparticle-assisted PCR (NanoPCR), offer high sensitivity and detect co-infections, though they require specialized equipment and extensive validation [41,121,144,147,179,213,214,215,216,217]. These platforms enable simultaneous detection of multiple pathogens, facilitating rapid differential diagnosis in complex clinical scenarios.
Isothermal methods such as recombinase-aided amplification (RAA) and recombinase polymerase amplification (RPA) (LAMP) provide rapid, portable detection and perform best when paired with microfluidics, lateral-flow strips, or CRISPR systems [218,219,220]. These approaches are particularly valuable for field deployment and resource-limited settings. CRISPR-Cas12a/Cas13a assays enable rapid, sensitive, and multiplex detection but remain costly and are not yet field-optimized [3,220,221,222,223,224]. These emerging platforms show considerable promise for point-of-care applications once scalability and cost challenges are addressed.
Serology, particularly ELISA, remains essential for herd surveillance and vaccine monitoring, although it is less effective for early piglet detection [211]. Advances include phage-ELISAs targeting M protein for TGEV differentiation, recombinant protein ELISAs for PEDV, M protein assays for PDCoV IgG, and multiplex immunoassays using recombinant S1 proteins that allow simultaneous detection of PEDV, TGEV, and PDCoV [171,225,226,227,228]. Rapid immunochromatographic strips (ICTs) offer significant advantages in point-of-care testing due to their brief testing duration, user-friendliness, and cost-effectiveness [229,230,231]. Histopathology combined with immunohistochemistry is valuable for confirming lesions and visualizing viral antigens in tissues, providing definitive diagnostic evidence in outbreak investigations.
Although isothermal amplification, sequencing, and nanotechnology offer promising advances, many diagnostics lack cross-strain validation. CRISPR-based methods need optimization, affordable portable tools are scarce, and next-generation sequencing is not yet widely applied in routine surveillance. Future priorities should focus on developing sensitive, cost-effective, field-ready diagnostics that enable earlier detection of emerging SECoV strains and facilitate real-time epidemiological monitoring.
4. Prevention and Control Measures
4.1. Current Vaccine Strategies
Current vaccination strategies for SECoVs primarily target pregnant gilts and sows to confer passive lactogenic immunity to neonatal piglets via colostrum and milk [237]. However, rapid mutation and frequent recombination within SECoV genomes have progressively reduced vaccine effectiveness [12,13,238], a challenge exacerbated by the emergence of PDCoV and SADS-CoV. Although the PEDV spike glycoprotein contains major neutralizing epitopes (COE, SS2, SS6, and 2C10) [239,240,241], extensive amino acid variation in contemporary G1b and G2 strains compromises antibody recognition [33]. Consequently, classical CV777-derived vaccines provide strong protection against homologous strains but limited efficacy against currently circulating G2 variants [242,243]. A comparative summary of key vaccine studies for SECoVs prevention, including methods, immune responses, efficacy, and limitations, is provided in Table 5.
Inactivated and Live-Attenuated Vaccines remain the primary platforms for TGEV and PEDV [32,243], while no licensed vaccines are available for PDCoV or SADS-CoV [123,244]. Live-attenuated vaccines induce robust immunity but pose safety concerns due to potential reversion via mutation or recombination [49]. Recent advances include ORF3-truncated GIIb PEDV strains, which elicit strong IgG and IgA responses in sows and provide partial cross-protection in piglets [245], as well as reverse-genetics-derived attenuated vaccines with deletions of interferon-antagonistic genes that improve genetic stability [246].
Inactivated vaccines are highly safe but weakly immunogenic, requiring booster doses and potent adjuvants. Parenteral administration induces limited mucosal and lactogenic immunity, restricting neonatal protection [49]. Adjuvant optimization has shown promise; for example, a ginseng-derived saponin mucosal adjuvant enhanced intestinal IgA responses and milk IgA levels when combined with inactivated PEDV vaccines [247]. Nevertheless, cross-protection against heterologous strains remains inconsistent.
In 2025, autogenous inactivated vaccines prepared from tissues infected with the PEDV G2c variant were rapidly deployed on farms, resulting in a 95% reduction in mortality and viral shedding. Genomic sequencing confirmed antigenic matching with circulating strains and showed no evidence of reversion risk [237,248].
Next-Generation Vaccine Platforms, including mRNA, virus-like particle (VLP), subunit, and vectored vaccines, aim to improve safety, mucosal immunity, and breadth of protection [96,243,244]. lipid nanoparticles-formulated mRNA vaccines enable rapid, flexible development and strong immunogenicity [249]. PEDV GIIb spike or S-M mRNA constructs elicited broad neutralizing antibodies (GI, GIIa, GIIb) in animal models and reduced disease severity in piglets [250], though viral shedding was not fully prevented.
VLP Vaccines: VLPs expressing PEDV E, M, and S induce strong mucosal and systemic immunity and reduce disease severity [251]. HEK293T- and baculovirus-derived VLPs elicit higher IgA and neutralizing antibodies than inactivated vaccines [252,253,254,255], supporting their potential as safe and effective SECoV platforms.
Subunit vaccines: Efforts focus on recombinant spike proteins and neutralizing domains. Bivalent S-trimer vaccines overcome limited GIIa/GIIb cross-protection, inducing strong lactogenic immunity and protection against both lineages [256]. Stabilized S-trimers and M103-adjuvanted S formulations elicit broad neutralizing responses and reduce disease and viral shedding in piglets [257,258]. Strain-matched GIIb vaccines further boost maternal titers, with pre-farrowing levels (~1:377–1:774) associated with >80% piglet protection [259].
Live bacterial/viral vectors: Oral recombinant Lactobacillus expressing PEDV S antigens induces strong mucosal and systemic immunity in sows, elevating milk SIgA and providing effective lactogenic protection [260,261]. In contrast, Adenoviral (Ad5) vectors encoding S or S1 elicit robust humoral responses after intramuscular delivery but have limited efficacy via the oral route [262].
For PDCoV, most vaccine studies are still in the preclinical stage. Highlighted among advances are mRNA plus nanoparticle-based, scalable, broad-spectrum protective coverage type future candidates [263]. Live attenuated PDCoV vaccine development has focused on genetic modification and serial passaging [52]. In 2020, an NS6-deficient mutant (rPDCoV-ΔNS6-GFP) was identified as a promising candidate [264], causing only mild clinical signs compared with the wild-type virus, indicating NS6 as a key virulence factor. This mutant was generated using an infectious cDNA clone of strain USA/IL/2014/026, enabling precise genetic engineering.
Inactivated vaccines provide partial lactogenic protection but fail to fully prevent disease [265]. Emerging platforms, including ferritin-based RBD nanoparticles, baculovirus-expressed subunit vaccines, and VLPs, induce robust immune responses and protection comparable to inactivated vaccines in experimental models [266,267,268].
Given the pronounced genetic and antigenic diversity of SECoVs, Polyepitopic vaccines incorporating conserved epitopes represent a promising strategy for achieving broader cross-protective immunity, supported by advances in in silico epitope design, though validation in swine remains essential [269].
4.2. Therapeutic and Antiviral Drugs
SECoV control remains limited by significant antigenic variation and gaps in ecological understanding [238]. The absence of licensed vaccines for several SECoVs and the modest efficacy of existing antivirals [3] underscore the urgent need for safe, effective therapeutics. Although hundreds of compounds have been screened, most exhibit single-mechanism activity with uncertain clinical relevance [3].
The main protease (Mpro) is highly conserved across PEDV, TGEV, PDCoV, and SADS-CoV [3], making it an attractive antiviral target. A summary of bioactive compounds with anti-SECoV activity and their proposed mechanisms of action is provided in Table 6. Several candidates that show promise include: Rhodamine derivative LJ001, which inhibits TGEV and PDCoV replication [296], while melatonin and related indole compounds (indole, tryptamine, L-tryptophan) impair viral entry and replication [297]; Rifampicin, which shows activity against PEDV and SADS-CoV [3]; Griffithsin, which blocks PEDV attachment and PDCoV entry through S-protein binding [298,299]; Surfactin, which inactivates TGEV and disrupts PEDV membranes [300,301]; and 25-hydroxycholesterol, which suppresses all major SECoVs and SARS-CoV-2 [157,302,303]. Ergosterol peroxide blocks PEDV and PDCoV entry, reduces apoptosis, and modulates cytokines through NF-κB and p38/MAPK pathways [304,305]. Lithium chloride (LiCl) suppresses PEDV and PDCoV replication and reduces apoptosis [306,307]. Octyl gallate (OG) is a 3CLpro inhibitor that protects piglets from PEDV in vivo [308]. In addition, RNA interference targeting PDCoV N or PEDV/SADS-CoV M genes effectively inhibits viral replication [309]. Drug repurposing of FDA-approved compounds provides a rapid, cost-effective strategy for identifying clinically relevant therapies. Host-targeted interventions are especially promising: for example, bile acids differentially modulate SECoV replication [310], enhancing PEDV [311], inhibit PDCoV [312], and promoting SADS-CoV via lipid raft-mediated endocytosis [313]. Such findings highlight critical virus-host differences that can guide rational antiviral design.
Natural products remain appealing antiviral candidates due to their low toxicity and reduced risk of resistance. Although systematic screening, structure–activity analyses, and compound optimization show promise, most findings are confined to in vitro studies. Validation in advanced models, such as organoids, 3D cultures, and piglet challenge systems, is still needed. Combination therapies targeting both viral and host pathways may further enhance therapeutic efficacy. Future research should integrate functional genomics, high-throughput screening, and computational drug design with rigorous preclinical evaluation to accelerate the development of safe, effective antivirals, an essential component of sustainable swine production.
4.3. Biosecurity Measures
Biosecurity refers to coordinated management and hygiene preventing pathogens from entering or spreading within livestock production [84,316]. No single measure is effective in isolation; protection relies on integrating multiple practices. Effective SECoV control requires understanding their diverse transmission routes. SECoVs spread primarily via the fecal-oral route, but PEDV and PDCoV also transmit through aerosols [32,34], and PEDV can spread lactogenically [81]. PEDV RNA detection in boar semen, with prolonged shedding in the sperm-rich fraction [80], suggests possible sexual transmission. Indirect transmission via contaminated feed, equipment, personnel, and transport trailers further amplifies risk [24,75,76,77]. Lactogenic and sexual routes are particularly problematic, as high viral loads in milk and semen can bypass standard biosecurity and undermine maternal antibody protection.
Critical practices:
Effective control of SECoVs relies on strict biosecurity measures that regulate the movement of animals, feed, vehicles, equipment, and personnel to limit pathogen introduction and spread. Facility sanitation should be strengthened through clearly defined Lines of Separation and Perimeter Buffer Areas, farm-specific clothing and boots, and handwashing or shower-in/shower-out protocols. Proper manure management, including fermentation or disinfection, and restrictions on staff and equipment movement between barns are critical to reducing indirect transmission. Feed and water supplies must be microbiologically safe, while transport vehicles and loading facilities, major sources of SECoV dissemination [24], require thorough cleaning and disinfection. Additional internal biosecurity measures, such as excluding wildlife and pets, routine disinfection and waste disposal, increased farm spacing, and reduced pig density, further limit viral circulation. Although controlled feedback exposure of pregnant sows to feces from infected piglets may enhance lactogenic immunity via the gut-mammary sIgA axis [317], this practice remains poorly standardized, produces inconsistent outcomes, and carries significant biosafety risks [32,191].
Implementation gaps remain substantial. Large commercial farms generally apply multi-layered biosecurity that reduces but does not eliminate-risk, whereas many smallholder systems lack fundamental measures such as effective waste management, air control, and movement restrictions, enabling viral persistence and transmission. Shared transportation further amplifies exposure risk [318]. Strengthening SECoV control will require updated farm-level data, risk-based biosecurity strategies tailored to production systems, and targeted producer training to address both structural and behavioral barriers.
5. Challenges and Future Perspectives
Despite extensive research on swine enteric coronaviruses, long-term control remains challenging due to genetic diversity, antigenic drift, and limited cross-protection among circulating strains. Conventional vaccines and surveillance systems often lag behind viral evolution, underscoring the need for real-time genomic monitoring and innovative immunization strategies. Integrating high-throughput sequencing, bioinformatics-driven epitope prediction, and advanced vaccine platforms (e.g., mRNA, viral vectors, and virus-like particles) represents a paradigm shift in swine coronavirus control.
Major challenges include spike-driven immune escape, antigenic divergence between PEDV genogroups (G1/G2), inadequate induction of mucosal IgA and lactogenic immunity, variable field performance of diagnostic tools, and persistent biosecurity gaps, particularly in smallholder production systems.
Research priorities include:
- Developing predictive models of viral evolution for real-time surveillance and early warning;
- Advancing broad-spectrum mucosal vaccines and delivery platforms to induce durable cross-protection and lactogenic immunity;
- Standardizing immunization evaluation frameworks with harmonized efficacy criteria;
- Implementing rapid, field-deployable diagnostics for early detection and strain differentiation;
- Elucidating host–pathogen interactions, including viral entry, immune evasion, and age-related susceptibility;
- Strengthening One Health-integrated surveillance to mitigate interspecies transmission risks.
Coordinated global surveillance at the human–animal interface is essential to track viral evolution, guide vaccine updates, and enhance preparedness for emerging coronavirus threats.
6. Conclusions
Integrated genomic surveillance, rapid field-deployable diagnostics, and next-generation vaccine platforms are essential for improving the control of swine enteric coronaviruses. Priorities include the development of broadly protective multivalent and mucosal vaccines, enhanced biosecurity and maternal immunization strategies, and real-time monitoring of infection dynamics and immune responses. Given the zoonotic potential of PDCoV and the broad cellular tropism of SADS-CoV, deeper mechanistic studies of pathogenicity and accelerated development of broad-spectrum antivirals and universal coronavirus vaccines are urgently needed. Proactive One Health surveillance frameworks that integrate animal and human health systems will be critical for early spillover detection and strengthened pandemic preparedness.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Thakor J.C. Dinesh M. Manikandan R. Bindu S. Sahoo M. Sahoo D. Dhawan M. Pandey M.K. Tiwari R. Emran T.B. Swine coronaviruses (S Co Vs) and their emerging threats to swine population, inter-species transmission, exploring the susceptibility of pigs for SARS-Co V-2 and zoonotic concerns Vet. Q.20224212514710.1080/01652176.2022.207975635584308 PMC 9225692 · doi ↗ · pubmed ↗
- 2Li Z. Chen Y. Li L. Xue M. Feng L. Different Infectivity of Swine Enteric Coronaviruses in Cells of Various Species Pathogens 20241317410.3390/pathogens 1302017438392912 PMC 10891669 · doi ↗ · pubmed ↗
- 3Liang J. Xu W. Pan X. Han S. Zhang L. Wen H. Ding M. Zhang W. Peng D. Advances research in porcine enteric coronavirus therapies and antiviral drugs Vet. Q.20244414910.1080/01652176.2024.242129939484691 PMC 11536681 · doi ↗ · pubmed ↗
- 4Jung K. Wang Q. Scheuer K.A. Lu Z. Zhang Y. Saif L.J. Pathology of US porcine epidemic diarrhea virus strain PC 21A in gnotobiotic pigs Emerg. Infect. Dis.20142066266510.3201/eid 2004.13168524795932 PMC 3966387 · doi ↗ · pubmed ↗
- 5Lin C.-M. Gao X. Oka T. Vlasova A.N. Esseili M.A. Wang Q. Saif L.J. Antigenic relationships among porcine epidemic diarrhea virus and transmissible gastroenteritis virus strains J. Virol.2015893332334210.1128/JVI.03196-1425589635 PMC 4337547 · doi ↗ · pubmed ↗
- 6Alluwaimi A.M. Alshubaith I.H. Al-Ali A.M. Abohelaika S. The coronaviruses of animals and birds: Their zoonosis, vaccines, and models for SARS-Co V and SARS-Co V 2Front. Vet. Sci.2020758228710.3389/fvets.2020.58228733195600 PMC 7543176 · doi ↗ · pubmed ↗
- 7Islam A. Ferdous J. Islam S. Sayeed M.A. Dutta Choudhury S. Saha O. Hassan M.M. Shirin T. Evolutionary dynamics and epidemiology of endemic and emerging coronaviruses in humans, domestic animals, and wildlife Viruses 202113190810.3390/v 1310190834696338 PMC 8537103 · doi ↗ · pubmed ↗
- 8Su S. Wong G. Shi W. Liu J. Lai A.C. Zhou J. Liu W. Bi Y. Gao G.F. Epidemiology, genetic recombination, and pathogenesis of coronaviruses Trends Microbiol.20162449050210.1016/j.tim.2016.03.00327012512 PMC 7125511 · doi ↗ · pubmed ↗