Polyphasic characterization of Nocardioides aquaegermanicae sp. nov., a novel water-derived actinobacterium
Shuangqing Zhou, Sarah Kirstein, Lara Keunecke, Alexandra Lehmann, Lars-Olaf Schulz, Marlen Jando, Gabriele Pötter, Meina Neumann-Schaal, Yvonne Mast, Imen Nouioui

TL;DR
Scientists discovered a new species of water-derived bacteria, Nocardioides aquaegermanicae, based on genetic and chemical analysis.
Contribution
A novel species of actinobacterium is described with distinct genomic and chemotaxonomic features.
Findings
The strain DSM 117947T has less than 70% DNA-DNA hybridization and 95-96% ANI with its closest relative.
The polar lipid profile and fatty acid composition distinguish the strain from related species.
Genomic G+C content of 73% and menaquinone MK-8(H4) are key features of the new species.
Abstract
Strain DSM 117947T was isolated from a Micromonospora matsumotoense co-culture originating from a water sample collected in Germany. The strain was subjected to a polyphasic taxonomic analysis. It exhibited 99.3% 16S rRNA gene sequence similarity with Nocardioides aurantiacus DSM 12652T. Digital DNA-DNA hybridization and average nucleotide identity values between the strain and its close phylogenetic neighbour were below the threshold of 70% and 95−96% for prokaryotic species demarcation, respectively. The strain had a polar lipid profile composed of diphosphatidylglycerol (DPG), phosphatidylethanolamine (PE), phosphatidylmethylethanolamine (PME), phosphatidylinositol (PI), glycophospholipid (GPL), and phospholipids (PLs). The predominant menaquinone (>20%) was MK-8(H4). The major fatty acids (>5%) were C16.0, C16:1 ω7c, and C18:1 ω9c. The genomic G + C content of the strain is 73%. The…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
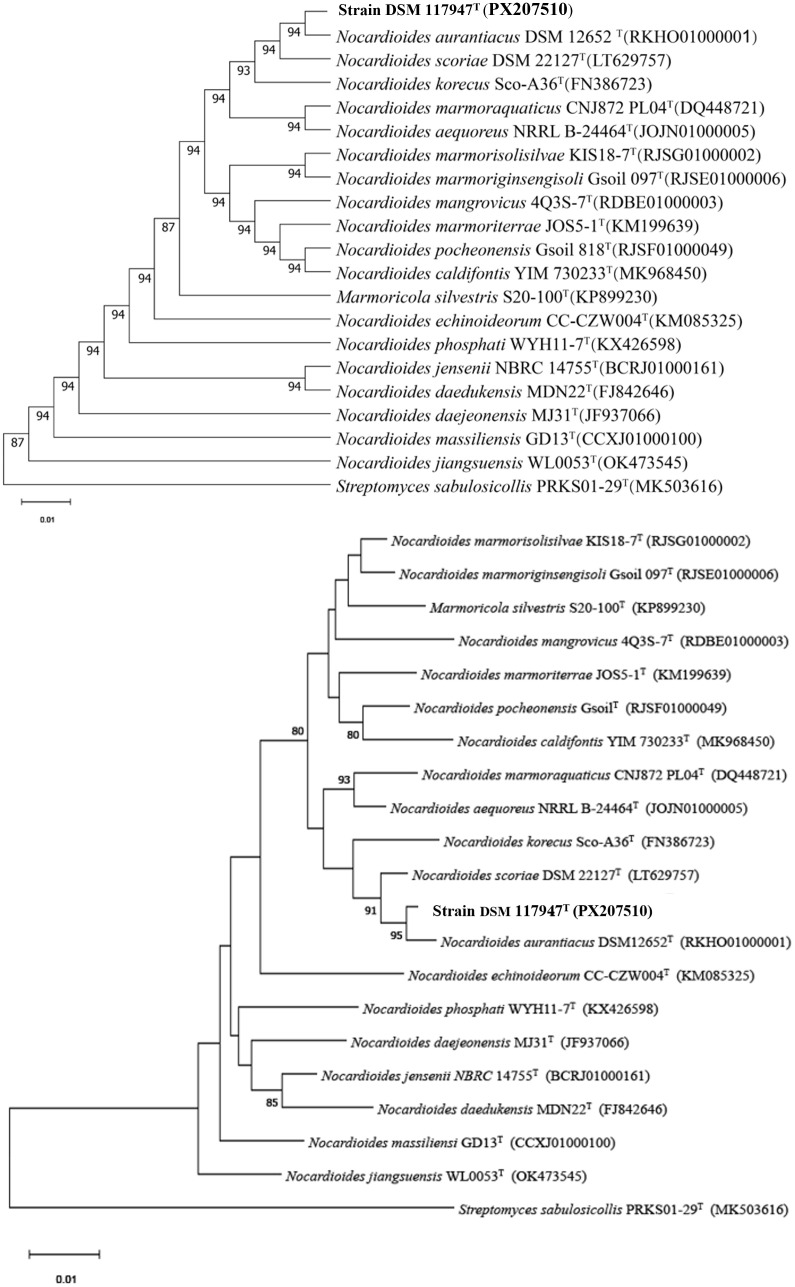 Fig 1
Fig 1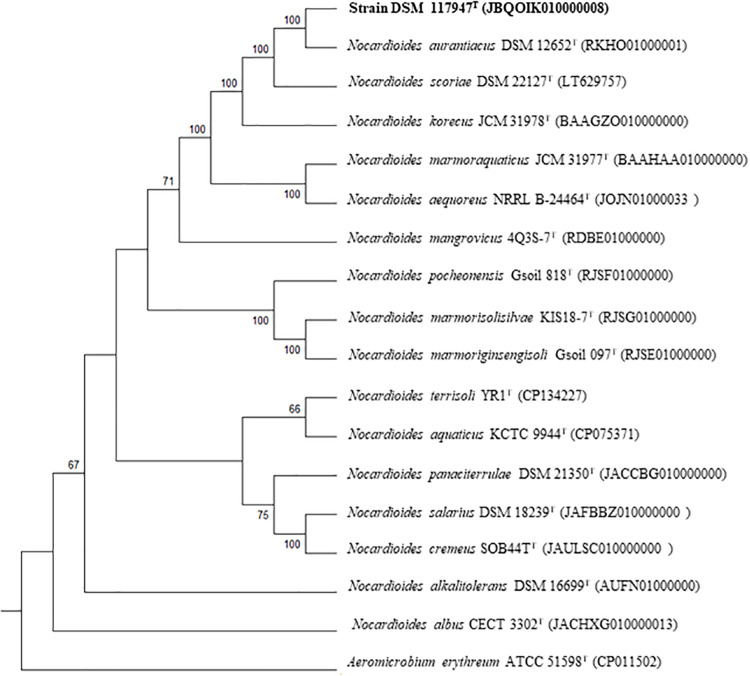 Fig 2
Fig 2Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGenomics and Phylogenetic Studies · Actinomycetales infections and treatment · Microbial Natural Products and Biosynthesis
Introduction
The genus Nocardioides of the family Nocardioidaceae within the order Propionibacteriales comprises 171 validly named species (https://lpsn.dsmz.de/ [accessed in July 2025]). Members of this genus have been isolated from different environments, including plastic waste [1], Arctic marine sediments [2], stony coral [3], waterfalls [4], the feces of Tibetan antelope [5,6], kaolinite clay [7], sewage sludge [8], the gastrointestinal tract of lake prawn [9], intestines of fish species [10], and cave stalactite surfaces [11]. Nocardioides strains are Gram-positive, aerobic, non-acid-fast bacteria, exhibiting coccoid, short rod-shaped, rod-shaped, or filamentous cell morphologies. Several Nocardioides species do not produce aerial mycelium. The cell wall peptidoglycan contains LL-diaminopimelic acid as the diagnostic diamino acid. The predominant menaquinone is MK-8 (H_4_), and the major cellular fatty acid is 14-methyl pentadecanoic acid (iso-C_16:0_). The average genome size is approximately5.7 Mb, with a DNA G + C contentranging from 67.5% to74.8 mol% [12].
Nocardioides strains are known for their ability to utilize various carbon and nitrogen sources, and degrade various organic compounds and environmental pollutants, such as hydrocarbons, haloalkanes, aromatic compounds, nitrogen-containing heterocycles, and polyester pollutants. For example, Nocardioides sp. KP7, Nocardioides simplex FJ2-1A, Nocardioides sp. JQ2195, Nocardioides carbamazepine, Nocardioides alcanivorans NGK65^T^, Nocardioides limicola DJM-14^T^, and Nocardioides sp. JWJ-L0 have been reported to degrade phthalates [13], 2,4-dinitroanisole [14], dibenzofuran [15], ibuprofen [16], hexadecane [17], alkanes [18], and polycyclic aromatic hydrocarbons [19], respectively. In the course of activating and clarifying the taxonomic status of old actinobacterial strains deposited at the DSMZ before the 1990s, several strains belonging to different orders of the phylum Actinomycetota were found to be candidates for novel species. In this context, Nocardioides sp. DSM 117947^T^ was obtained from a purification of a culture of Micromonospora matsumotoense isolated from water collected in Germany and deposited at the DSMZ open culture collection. Given the bioremediation potential of Nocardioides, strain DSM 117947^T^ was subjected to a comprehensive polyphasic taxonomic analysis, including comparative genomic studies. The results demonstrated that the strain represents a novel species of the genus Nocardioides, for which the name Nocardioides aquaegermanicae sp. nov. is proposed.
Materials and methods
Origin and maintenance
The strain DSM 117947^T^ (WG_orange^T^ = KCTC 59414^T^) was isolated from a co-culture of Micromonospora matsumotoense, which was isolated from water collected in Germany before 1993 and deposited at the DSMZ culture collection. The strain was purified after serial dilution of the original culture. One hundred microliters of bacterial suspension (dilution 10^−6^) dissolved in NaCl solution (0.9% w/v) was spread on GYM medium [glucose (4 g), yeast extract (4 g), malt extract (10.0), CaCO_3_ (2.0), agar 20.0g, H_2_O (1l), pH 7.2] and incubated at 28 °C for 10 days. A single colony was streaked onto GYM medium and incubated at 28 °C for 10 days. The purity of strain DSM 117947^T^ was confirmed using a light microscope (Nikon Eclipse, 100x objective). For comparative analysis, the closest phylogenetic neighbor of the strain, Nocardioides aurantiacus DSM 12652^T^, was included in the study. Information on the origin, history, and growth conditions of strain DSM 12652^T^ is available in the DSMZ online catalogue (https://www.dsmz.de/collection/catalogue). Phenotypic characterisation and molecular identification were carried out using wet biomass from a 10-day-old culture grown in ISP2 liquid medium [glucose (4 g), yeast extract (4 g), malt extract (10.0), agar 20.0g, H_2_O (1l), pH 7.2] at 28 °C with shaking at 150 rpm. Chemotaxonomic studies were performed using freeze-dried cells, except for fatty acid and menaquinone analyses, for which specific sample preparation protocols were applied.
Molecular identification and phylogenetic studies
To determine the taxonomic status of strain DSM 117947^T^ at the species rank, genomic DNA was extracted as described previously [20] and the 16S rRNA gene was amplified by PCR using primers 27F (5′–AGAGTTTGATC(AC)TGGCTCAG–3′) and 1492R (5′–ACGG(CT)TACCTTGTTACGACTT–3′) [21]. Amplicons were sequenced using the Applied Biosystems (ABI) 96-capillary-system, as described by Risdian et al. [22]. The nearly complete 16S rRNA gene sequence (1525 bp) of strain DSM 11794^T^ was compared with those of validly named Nocardioides species, using the EzBioCloud server [23]. The pairwise 16S rRNA gene sequence similarity between strain DSM 117947^T^ and its closest phylogenetic neighbour was determined using EzBioCloud server. Maximum-likelihood (ML) [24–25] and Neighbour Joining (NJ) [26] phylogenetic trees were constructed via MEGA software (v 11.0) with 1000 replicates [27].
Comparative genomic studies
Biomass harvested from strain DSM 117947^T^ culture, grown under the conditions described above, was used for DNA extraction and whole-genome sequencing by MicrobesNG (Birmingham, UK; https://microbesng.com/). The Illumina NovaSeq 6000 (Illumina, San Diego, USA) platform with a 250 bp paired-end protocol was used for sequencing. Library construction, assembly, and quality control were carried out by the service. Genome quality (completeness and contamination) was assessed via CheckM (v1.2.4) [28]. Genome of the strain was automatically annotated by the NCBI GenBank via PGAP (Prokaryotic Genome Annotation Pipeline) (https://www.ncbi.nlm.nih.gov/datasets/genome/GCF_052591715.1/) [29]. The genome sequence was deposited in the DDBJ/ENA/GenBank database under the accession number JBQOIK000000000.
Genome-based phylogeny and digital DNA–DNA hybridization (dDDH) values between strain DSM 117947ᵀ and its closest phylogenomic relatives were carried out using the Type Strain Genome Server (TYGS) (https://tygs.dsmz.de/) [30–32]. Average nucleotide identity (ANI) scores between the strain and its neighbours were calculated using the ANI calculator available in the EzBioCloud server (https://www.ezbiocloud.net/tools/ani) [33].
Chemotaxonomic properties of strain DSM 117947T
Freeze-dried cells of strain DSM 117947^T^ and its close relative N. aurantiacus DSM 12652^T^ were analysed, using thin layer chromatography, for whole-cell sugars [34,35], polar lipids [36], and diaminopimelic acid isomers [37]. Isoprenoid quinones of strain DSM 117947^T^ were examined by high-performance liquid chromatography (HPLC) coupled with a diode array detector and high-resolution mass spectrometry (MS) [38]. Cellular fatty acid profiles of the strains were determined using gas chromatography–mass spectrometry (GC-MS) on an Agilent GC-MS 7000D instrument [39]. The position of double bonds in fatty acids was determined by derivatization of fatty acid methyl esters with dimethyl disulfide [40].
Growth properties, biochemical, and enzymatic features of strain DSM 117947T
The growth of strain DSM 117947^T^ was assessed on a variety of media, including DSMZ 1746 (ISP1), DSMZ 987 (ISP2), DSMZ 84 (ISP3), DSMZ 252 (ISP4), DSMZ 993 (ISP5), DSMZ 1269 (ISP6), DSMZ 1619 (ISP7), DSMZ 535 (TSA, Tryptic Soy Agar), DSMZ 65 (Glucose-Yeast-Malt extract, GYM), DSMZ 83 (Czapek’s agar), and DSMZ 554 (N-Z-Amine agar). The composition of these media is available in the cultivation media database (https://mediadive.dsmz.de/). Growth was tested across a range of different temperatures (4, 10, 16, 20, 25, 28, 35, 37, 42, and 45°C), pH (5, 5.5, 6.0,6.5, 7.0,7.5, 8.0,8.5, and 9.0), and salinity (0%, 2.5%, 5%, 7.5%, and 10% NaCl w/v). The following buffer systems were used: 0.1 M citric acid/ 0.1 M sodium citrate for pH < 6; 0.1 M KH_2_PO_4_/0.1 M NaOH for pH 8.0–8.5; and 0.1 M NaHCO_3_/0.1 M Na_2_CO_3_ for pH 9.0. The morphology of the strain including the colour of the substrate mycelium was determined using RAL colour charts [41]. The cell structures from 7- and 14-day-old cultures, prepared on GYM medium at 28 °C, were observed using a lightmicroscope (Nikon Eclipse, 100x objective).
The biochemical and enzymatic profiles of strain DSM 117947ᵀ and its closest phylogenomic neighbour, N. aurantiacus DSM 12652^T^ were examined using API 20NE, API 50CH, and API ZYM test strips according to the manufacturer’s instructions (bioMérieux, France). All tests were inoculated with a homogeneous bacterial suspension equivalent to a density of 5 on the McFarland scale, prepared from cultures grown in ISP2 liquid medium for 10 days at 28°C with shaking at 150 rpm.
Potential applications of strain DSM 117947T
The genome sequence of strain DSM 117947^T^ and its close phylogenomic neighbour, N. aurantiacus DSM 12652^T^, were screened for secondary metabolite biosynthetic gene clusters (BGCs) using the antiSMASH web tool version 8.0 with default settings (https://antismash.secondarymetabolites.org) [[42]]. Antimicrobial activity of crude extracts from strains DSM 117947^T^ and DSM 12652^T^ was tested against Gram-negative bacteria (Escherichia coli ΔtolC JW5503–1 and Proteus vulgaris DSM 2140), Gram-positive bacteria (multi-resistant Staphylococcus aureus DSM 18827 and Enterococcus faecium DSM 20477^T^), and yeast (Candida albicans DSM 1386). Crude extracts were prepared from 10-day old cultures of the strains grown in 50 ml ISP2, NL19, NL800, and R5 media, respectively, at 28°C with shaking at 180 rpm, as described previously [43]. The potential of strain DSM 117947^T^ for xenobiotic biodegradation and neutralising biotic and abiotic stresses was evaluated using the PGPT-Pred tool, available in PLaBAse server (https://plabase.cs.uni-tuebingen.de/pb/plabase.php). The aligner used for this study is Blast and blastp+hmmer [44–46].
Results and discussion
Taxonomic novelty and distinctive genomic features of strain DSM 117947T
The authenticity of strain DSM 117947^T^ was confirmed by comparing the 16S rRNA gene sequence obtained by PCR with that extracted from the whole genome sequence. The 16S rRNA gene sequence similarity values between strain DSM 117947^T^ and validly named Nocardioides species ranged from 94.7% to 99.3%, with N. aurantiacus (99.3%) identified as the closest relative (S1 Table). These results were consistent with the close phylogenetic relatedness between DSM 117947^T^ and N. aurantiacus in the ML and MP phylogenetic trees, in which these strains formed a well-supported sub-cluster next to Nocardioides scoriae (Fig 1).
ML and NJ phylogenetic trees based on the 16S rRNA gene sequences showing the phylogenetic position of strain DSM 117947T within the radiation of the genus Nocardioides.The numbers above the branches are support values when greater than 50% from ML and NJ bootstrapping. Bar indicates 1 nucleotide substitutions per 1000 nucleotides.
The genomes of strain DSM 117947^T^ and its close neighbour N. aurantiacus DSM 12652^T^ had a size of 4.1 Mbp and 4.0 Mbp, with G + C contents of 73.0% and 72.5%, containing 49 and 54 RNAs, and 4,077 and 3,905 coding sequences, respectively (Table 1). The genomic features of strain DSM 117947^T^ were consistent with those reported for the genus Nocardioides, which typically exhibit G + C contents ranging from 67.5 to 74.8% [12]. The whole genome-based tree further confirmed the close phylogenetic relationship between strain DSM 117947ᵀ and N. aurantiacus (Fig 2). The dDDH and ANI values between the genome sequences of DSM 117947^T^ and N. aurantiacus DSM 12652^T^ were 35.2% and 88.6%, respectively; values well below the accepted species delineation thresholds of 70% for dDDH and 95–96% for ANI [49,50] (S2 Table).
Table 1: Biochemical, enzymatic and genomic features that distinguish strain DSM 117947T from its close phylogenetic neighbour Nocardioides aurantiacus DSM 12652T.
Tree inferred with FastME from GBDP distances calculated from genome sequences.The branch lengths are scaled in terms of GBDP distance formula d5. Only Bootstrap values above 60% are displayed above the branches.
Growth properties, phenotypic, and chemotaxonomic features of strain DSM 117947T
The phenotypic and morphological characteristics of strain DSM 117947^T^ were consistent with those of the genus Nocardioides [23]. The strain grew well on ISP1, ISP2, ISP6, TSA, GYM, Czapek’s agar, and N-Z-Amine agar media, forming a yellow substrate mycelium after 10 days of incubation at 28°C (RAL 1028). The morphology of the strain in GYM medium plate is available in the DSMZ online catalogue (https://www.dsmz.de/collection/catalogue/details/culture/DSM-117947). Strain DSM 117947^T^ showed good growth on GYM medium at 16, 20, 25, 28, and 37°C. The strain grew at pH values ranging from 5.5 to 9.0 and tolerated up to 2.5% NaCl in GYM medium. Optimal growth on GYM medium was observed at 28, 35, and 37°C at pH 7.5–8.5, and in the absence of added salt. The strain developed yellow colonies under optimal growth condition (RAL 1028). Coccoid-shaped cells were observed in 15 days-old culture after incubation at 28°C (S1 Fig).
Strain DSM 117947^T^ was able to metabolise L-arabinose, cellobiose, D-fructose, galactose, D-glucose, glycerol, maltose, D-mannose, melibiose, D-raffinose, trehalose, and D-xylose, as sole carbon sources (Table 1). Strain DSM 117947^T^ was distinguished from its close phylogenetic neighbour, strain DSM 12652^T^, by its ability to hydrolyse gelatin, oxidise tyrosine, and produce alkaline phosphatase, α-galactosidase, and lipase (Table 1). In addition, strain DSM 117947^T^ could be distinguished from its phylogenetic relatives, Nocardioides scoriae and Nocardioides korecus, and the type species, Nocardioides albus, by several biochemical and enzymatic features as shown in Table 1.
Whole-cell hydrolysates of strain DSM 117947^T^ were rich in LL-diaminopimelic acid (LL-DAP) and galactose, glucose, and ribose as whole cell sugars while N. aurantiacus DSM 12652ᵀ lacked galactose [51]. The polar lipid profile of strain DSM 117947^T^ consisted of diphosphatidylglycerol (DPG), phosphatidylethanolamine (PE), phosphatidylmethylethanolamine (PME), phosphatidylinositol (PI), glycolipid (GL), and phospholipids (PLs) (S2 Fig), whereas its close relatives N. aurantiacus, N. korecus and N. scoriae lacked PE and contained phosphatidylglycerol (PG) including the type species N. albus [12,47,48,51] (Table 1). The menaquinone profile of the strain consisted of MK-8 (H_4_) (89.1%) and MK-7 (H_4_) (12.6%); coherent with its close relatives (Table 1) [12,47,48,51]. The major fatty acids (>5%) of strains DSM 117947ᵀ and DSM 12652^T^,were C_16.0_, C_16:1_ ω7c, and C_18:1_ ω9c (Table 1). C_16.0_ and C_18:1_ ω9c were also present in N. korecus and N. scoriae, while the type species exhibited a different profile, reflecting its distant phylogenetic relationship to the studied strain (Table 1).
Potential applications of strain DSM 117947T
Genome mining for secondary metabolite BGCs revealed that strain DSM 117947^T^ and its close relative N. aurantiacus DSM 12652^T^ each possessed only a single terpene BGC, showing certain similarity to an isorenieratene and carotenoid BGC, respectively (S3 Table). These findings were consistent with the absence of antimicrobial activity of the crude extracts prepared from various production media when tested against different microbial strains, including Gram-positive and Gram-negative bacteria, as well as yeast.
In silico genome analysis of strains DSM 117947^T^ and DSM 12652^T^ revealed the presence of genes associated with the degradation of various aromatic compounds, including benzoate, biphenyl, gentisate, p-hydroxybenzoate, quinate, naphthalene, protocatechuate, toluene, salicylate, xylene and (S4 Table). In addition, strain DSM 117947^T^ harboured in its genome several genes whose products are involved in xenobiotics biodegradation, including oil (dichloroethane, vinylchloride, dichloropropene, dichloromethane), and steroid (adrostenedione, epiandrosterone, cholesterol) and styrene (acrylamide, phenylacetate) degradation. Genes associated with heavy metal detoxification (arsenic, bismuth, cadmium, chromate, cobalt, copper, iron, lead, manganese, nickel, selenium, tellurium, and zinc) were also present in the studied genomes. These compounds, which possess diverse toxicological properties, can originate from natural sources or anthropogenic activities, and their accumulation in the environment disrupt ecosystem functions and pose a threat to human health [52,53].
Furthermore, the genomic sequences of strains DSM 117947^T^ and DSM 12652^T^ were rich in genes controlling environmental stresses (heat shock, cold shock, heat inducible, high and low temperature), reflecting their ability to adapt to different ecological habitats and conditions. The strains seemed to possess the genetic machinery necessary to neutralise acidic (soxalate and spermidine), herbicidal (organophosphate and toxoflavin), osmotic (cardiolipin, trigonelline, glycine-betaine metabolism), salinity (magnesium, sodium, proline, riboflavin, pyridoxine, and folate metabolisms), and oxidative (peroxidised compounds, arylpolyene, carotenoids, glutathione, lipoic acid, terpenoids, spermidine and ubiquinone) stresses. Moreover, genes whose products are involved in the resistance to volatile compounds were also detected.
Several microorganisms, including actinobacteria such as Mycolicibacterium vinylchloridicum, have been found to mineralize such pollutants and their derivatives [54].These findings reflect the bioremediation potential of the studied strains.
These findings were consistent with the previously reported bioremediation potential of several species of the genus Nocardioides [13–19].
This study provides insight into the genetic machinery involved in the catabolism of polycyclic aromatic hydrocarbons. However, further experimental validation is needed to assess the effectiveness with which the proposed novel type strain DSM 117947^T^ degrades environmentally toxic substances.
Conclusions
Phenotypic, phylogenetic, and genomic data distinguished strain DSM 117947^T^ from all validly named Nocardioides species, supporting its classification as a representative of a novel species, for which the name Nocardioides aquaegermanicae sp. nov., is proposed.
Description of Nocardioides aquaegermanicae sp. nov
aquaegermanicae (a.quae.ger.ma’ni.cae. L. fem. n. aqua, water; L. fem. adj. germanica, German; N.L. gen. fem. n. aquaegermanicae, of the German water, referring to the freshwater source in Germany from which the type strain was isolated).
Gram-stain-positive, aerobic, catalase positive, oxidase negative with coccoid-shaped cells. The strain exhibits good growth on Czapek’s agar, GYM, ISP1, ISP6, N-Z-Amine agar, and TSA media, producing an orange substrate mycelium. Optimal growth occurs at 28°C, 35°C, and 37°C, within a pH range of 7.5–8.5, and in the absence of salt. The strain utilises L-arabinose, cellobiose, D-fructose, galactose, D-glucose, glycerol, maltose, D-mannose, melibiose, D-raffinose, trehalose, and D-xylose as sole carbon sources. The polar lipids are diphosphatidylglycerol (DPG), phosphatidylethanolamine (PE), phosphatidylmethylethanolamine (PME), phosphatidylinositol (PI), glycolipid (GL), and phospholipids (PLs). The predominant menaquinone (>20%) is MK-8 (H4). The major fatty acids (>5%) are C_16.0_, C_18:1_ ω9c, and C_16:1_ ω7c. The genome size of the type strain DSM 117947^T^ is 4.1 Mbp with a G + C content of 73%.
The type strain DSM 117947^T^ (WG_orange^T^ = KCTC 59414^T^) was isolated from a Micromonospora matsumotoense co-culture originating from a water sample collected in Germany. The 16S rRNA gene and whole genome sequences have been deposited at NCBI GenBank under accession numbers PX207510 and JBQOIK000000000, respectively.
This Whole Genome Shotgun project has been deposited at DDBJ/ENA/GenBank under the accession JBQOIK000000000. The version described in this paper is version JBQOIK000000000.
Repositories
The GenBank accession number for the 16S rRNA gene and genome sequences of strain DSM 117947^T^ are PX207510 and JBQOIK000000000, respectively.
Supporting information
S1 FigMorphological features (left) and cell structures (right) of strain 117947^T^ grown on GYM medium at 28 °C for 15 days.The picture on the right was taken using a light microscope (Nikon Eclipse, 100x objective).(DOCX)
S2 FigTwo-dimensional TLC plate of polar lipids extracted from strain DSM 117947^T^ stained with molybdatophosphoric acid (a), molybdenum blue (b), ninhydrin (c), anisaldehyde (d), Dragendorff (e) reagents.Key: DPG, diphosphatidylglycerol; PE, phosphatidylethanolamine; PI, phosphatidylinositol; PLs, phospholipids; GL glycolipid: PME, phosphatidylmethylethanolamine. Solvent1: chloroform: methanol: distilled water (65:25:4 v/v/v/); solvent 2: chloroform: glacial acetic acid: methanol: distilled water (80:12:15:4 v/v/v).(DOCX)
S1 Table16S rRNA gene sequence similarity between strain DSM 117947^T^ and its close phylogenetic neighbours.(DOCX)
S2 TabledDDH and ANI between the genome sequence of strain DSM 117947^T^ and its close phylogenomic relatives.(DOCX)
S3 TableBGCs associated with terpene biosynthesis of strain DSM 117947^T^ and N. aurantiacus DSM 12652^T^ assessed using AntiSMASH.(DOCX)
S4 TableStress envinronmental genes of Nocardioides sp. DSM 117947^T^.(XLSX)
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Mitzscherling J, Mac Lean J, Lipus D, Bartholomäus A, Mangelsdorf K, Lipski A, et al. Nocardioides alcanivorans sp. nov., a novel hexadecane-degrading species isolated from plastic waste. Int J Syst Evol Microbiol. 2022;72(4). doi: 10.1099/ijsem.0.005319 35482521 · doi ↗ · pubmed ↗
- 2Cong B, Zhang H, Li S, Liu S, Lin J, Deng A, et al. Taxonomic Identification of the Arctic Strain Nocardioides Arcticus Sp. Nov. and Global Transcriptomic Analysis in Response to Hydrogen Peroxide Stress. Int J Mol Sci. 2023;24(18):13943. doi: 10.3390/ijms 241813943 37762246 PMC 10531085 · doi ↗ · pubmed ↗
- 3Shi S-B, Cui L-Q, Zeng Q, Long L-J, Tian X-P. Nocardioides coralli sp. nov., an actinobacterium isolated from stony coral in the South China Sea. Int J Syst Evol Microbiol. 2022;72(5). doi: 10.1099/ijsem.0.005342 35511243 · doi ↗ · pubmed ↗
- 4Kim I, Chhetri G, Kim J, Kang M, So Y, Lee B, et al. Nocardioides donggukensis sp. nov. and Hyunsoonleella aquatilis sp. nov., isolated from Jeongbang Waterfall on Jeju Island. Int J Syst Evol Microbiol. 2021;71(12). doi: 10.1099/ijsem.0.005176 34928203 · doi ↗ · pubmed ↗
- 5Cheng Y, Jiao Y, Zhang S, Yang J, Lu S, Jin D, et al. Nocardioides dongkuii sp. nov. and Nocardioides lijunqiniae sp. nov., isolated from faeces of Tibetan antelope (Pantholops hodgsonii) and leaves of dandelion (Taraxacum officinale), respectively, on the Qinghai-Tibet Plateau. Int J Syst Evol Microbiol. 2021;71(7). doi: 10.1099/ijsem.0.004920 34313583 · doi ↗ · pubmed ↗
- 6Zhang G, Liu Y, Cheng Y, Yang J, Jin D, Lu S, et al. Identification of Nocardioides marmotae sp. nov. and Nocardioides faecalis sp. nov., two new members of the genus Nocardioides. Int J Syst Evol Microbiol. 2023;73(5). doi: 10.1099/ijsem.0.005875 37232277 · doi ↗ · pubmed ↗
- 7Yoon DJ, Cho ES, Hwang CY, Nam YD, Park SL, et al. Nocardioides luti sp. nov., belonging to the family Nocardioidaceae isolated from kaolinite, exhibiting the biosynthesis potential of alkylresorcinol. Antonie van Leeuwenhoek. 2021;114:983–95.33864547 10.1007/s 10482-021-01570-2 · doi ↗ · pubmed ↗
- 8Zhang X, Wu Z, Yan C, Chen C, Zheng G, Ye Y, et al. Nocardioides malaquae sp. nov., a novel actinobacterium isolated from sewage sludge of a fisheries processing factory. Int J Syst Evol Microbiol. 2021;71(7). doi: 10.1099/ijsem.0.004852 34241591 · doi ↗ · pubmed ↗