Discovery of Widespread Migrasome Formation During Amoeboid Migration in Dictyostelium discoideum
Bridget K Plude, Cynthia K Damer

TL;DR
The study shows that migrasomes form during cell migration in Dictyostelium and may help with communication and development.
Contribution
The discovery of migrasome formation during amoeboid migration in Dictyostelium discoideum is novel.
Findings
GFP-CpnE labels retraction fibers and migrasomes during chemotaxis and migration.
Migrasomes contain RNA and are stained by WGA and SYTO RNASelect.
Abstract
Migrasomes are organelles that form along retraction fibers of migrating cells and mediate intercellular communication. We performed localization studies on the calcium-dependent phospholipid-binding protein, copine E (CpnE), in Dictyostelium discoideum . We observed GFP-CpnE labeled retraction fibers and migrasomes in multiple contexts: chemotaxis toward folate, random migration, and early development. Migrasome membranes were stained with WGA and some migrasomes were stained with SYTO RNASelect, indicating the presence of RNA. Our findings suggest that CpnE may have a role in migrasome formation and indicate that migrasomes may play important roles in Dictyostelium development, cell communication, and/or homeostasis in a variety of environments.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
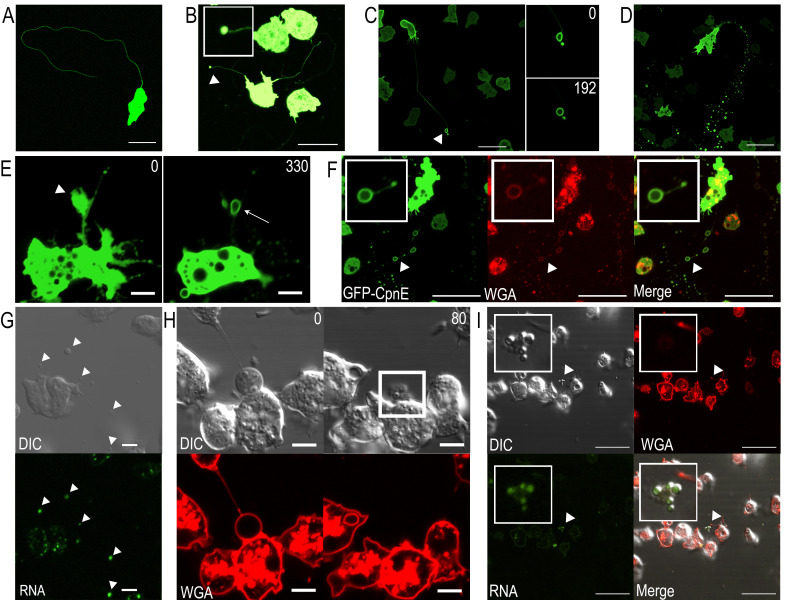 Figure 1
Figure 1|
Cell strain /plasmid name |
Description |
Origin |
Available from |
|
NC4A2 |
Axenic
|
Knecht & Sheldon 1995 |
Dicty Stock Center |
|
pTX-GFP/CpnE |
Extrachromosomal vector (pTX-GFP) with
|
Levi et al. 2000 (pTX-GFP); Ilacqua et al. 2018 (pTX-GFP/CpnE) |
This lab |
- —National Institutes of Health (United States)https://ror.org/01cwqze88
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCellular Mechanics and Interactions · Lipid Membrane Structure and Behavior · Cellular transport and secretion
Description
Migrasomes are vesicular organelles that form along retraction fibers of migrating cells (Ma et al. 2015). They contain multiple smaller vesicles carrying diverse cargo such as damaged mitochondria, autophagosomes, signaling molecules, and mRNA (Jiang et al. 2019; Jiao et al. 2021; Zhu et al. 2021; Lee et al. 2024). Migrasomes are released from retraction fibers in a process known as migracytosis and contribute to intercellular communication, morphogen transfer during development, mitochondrial quality control, and immune cell guidance in animals (Lim et al. 2015; Ma et al. 2015; Jiang et al. 2019; Jiao et al. 2021). Migrasomes are also made by cancer cells, where they promote tumor growth and metastatic spread (Zhang et al. 2024; Huang et al. 2025; Wang et al. 2025). A recent study confirmed their presence in the amoeba, Dictyostelium discoideum , forming when cells migrate on highly adhesive surfaces (Yu et al. 2025). Here, we expand on migrasome research in Dictyostelium by identifying additional migration processes involving these organelles.
Dictyostelium lives in the soil as single amoebae feeding on bacteria when food is available but develops into multicellular structures when starved. In both states, the cells use chemotaxis: to follow folate released by bacteria during feeding and to follow cAMP signals released by other cells during development (Kessin 2001). We are using Dictyostelium to study a conserved family of calcium-dependent phospholipid-binding proteins called copines. This study focuses on Copine E (CpnE), one of six copine proteins identified in Dictyostelium . In a previous study, we showed that GFP-tagged CpnE is a soluble cytosolic protein that binds strongly to phosphatidylserine and phosphatidylinositol-4-phosphate in the presence of calcium. In starved cells, GFP-CpnE exhibited a rapid and robust translocation from the cytosol to the plasma membrane in response to cAMP stimulation. This response was distinct from the other copine family members examined, which exhibited slower kinetics, incomplete translocation, and less sustained membrane association after cAMP stimulation (Ilacqua et al. 2018), suggesting CpnE may have a unique role in cAMP chemotaxis. While studying CpnE localization during chemotaxis, we discovered structures consistent with migrasomes.
Cells expressing GFP-tagged CpnE (GFP-CpnE) were used in an under-agar folate chemotaxis assay and visualized with confocal microscopy. As cells migrated, they produced long, thin GFP-CpnE labeled trails ( Figure 1A ). At the end of trails, there were often small GFP-CpnE labeled vesicular structures that remained stationary while the cell continued migrating ( Figure 1B, arrowhead, inset). At the time of our initial observations, migrasomes had not been reported in Dictyostelium . However, because they showed striking similarities to retraction fibers and migrasomes reported in animal cells, we hypothesized that these structures were indeed migrasomes.
Next, we asked whether these structures formed when cells were randomly moving in media on glass, without physical compression. Again, we observed what we believed were retraction fibers and migrasomes; however, the migrasomes appeared larger. Due to the larger size, we were able to visualize that CpnE was associated with the membrane of the migrasomes ( Figure 1C, arrowhead). Using time-lapse imaging, we were able to capture the formation of a migrasome as it became more circular over time ( Figure 1C, right images).
We then asked if cells made migrasomes during aggregation, the first stage of development. Aggregating cells produced migrasomes that were often found in clusters or in a chain marking a migration path ( Figure 1D ). Time-lapse imaging captured a large bulbous structure extending off a retraction fiber ( Figure 1E, arrowhead) that transformed into a ringlike structure ( Figure 1E, white arrow) over time, indicating CpnE recruitment from the cytosol to the membrane. Next, we used wheat germ agglutinin (WGA) to label migrasomes during aggregation. WGA labels glycoproteins and glycolipids found on the extracellular surface of membranes and has been routinely used to identify migrasomes and distinguish them from non-membranous structures and cellular debris (Chen et al. 2019). The GFP-CpnE signal and WGA stain colocalized on migrasome membranes ( Figure 1F, insets).
To determine if Dictyostelium migrasomes contain mRNA, we used cells that were not expressing GFP-CpnE and stained them with SYTO RNASelect and imaged cells with differential interference contrast (DIC) and confocal microscopy. We found examples of both large and small migrasomes that were labeled with SYTO RNASelect ( Figure 1G, arrowheads). We also used time-lapse imaging to capture a large migrasome that ruptured ( Figure 1H ). Once ruptured, the migrasome membrane was no longer stained with WGA ( Figure 1H, white box). By refocusing on the expelled contents, we found that the inner vesicles were stained with the SYTO RNASelect ( Figure 1J ), indicating the presence of RNA.
Our novel findings demonstrate that migrasome formation occurs under three different migratory conditions in Dictyostelium : chemotaxing under agarose towards folate, migrating randomly in media on glass, and aggregating under starvation conditions on glass. Retraction fibers and migrasomes were observed in all cases. However, retraction fiber length and migrasome number and size varied. The longest retraction fibers and smallest migrasomes were observed in cells migrating under agar. We hypothesize agar compression may disrupt migrasome formation, which is why the migrasomes were smaller in the under-agar experiments. When allowed to freely form in random migration and aggregation, the retraction fibers were more difficult to image because they were able to float away from the glass, while the migrasomes were able to grow and stabilize over time. Notably, cells that were not expressing GFP-CpnE appeared to make the largest migrasomes ( Figure 1H ). We did not observe migrasomes associated with every cell during imaging experiments, and because migrasomes are released from retraction fibers, it was often difficult to link individual migrasomes to their cell of origin. Future studies will examine the factors governing migrasome formation and why only some cells are observed with associated migrasomes within a given field of view.
Our observations indicate that migrasome formation in Dictyostelium occurs during early aggregation. In this context, migrasomes may act as concentrated signal packages, carrying mRNA or signaling molecules to enhance intercellular coordination. Alternatively, they may serve a homeostatic role, allowing cells to dispose of unnecessary or damaged components during the developmental transition. The enrichment of CpnE on these structures suggests CpnE could have a role in migrasome formation, perhaps by selecting cargo or enhancing vesicular stabilization. Calcium has been shown to be essential for migrasome formation and Synaptotagmin-1, a protein with two C2 domains like copines, is critical for the formation of migrasomes (Han and Yu 2024). Functional studies using cpnE mutant cells will be required to determine whether CpnE plays a critical role in migrasome biogenesis and/or function. Our results point to a previously uncharacterized vesicular pathway in Dictyostelium , potentially important for development, communication, and/or homeostasis.
Methods
Cell Culture
The Dictyostelium discoideum strain used was NC4A2, an axenic strain made from the NC4 strain (Knecht and Shelden 1995) and was obtained from the Dicty Stock Center (Fey et al. 2013). Cells were grown in VL6 medium (Formedium, VL60102) supplemented with penicillin-streptomycin at 60 U/mL at 18°C on plastic Petri dishes. The cDNA for
- cpnE* was subcloned into the
- SacI* site of the pTX-GFP plasmid (Levi et al. 2000; Ilacqua et al. 2018). The plasmid was electroporated into NC4A2 cells; cells were pulsed twice with a 5 second interval at 0.85 kV. Cells were placed on ice for 5 minutes and then plated in VL6. Transformants (NC4A2/GFP-CpnE cells) were selected using G418 (geneticin) at 60 µg/mL after 24 hours. Expression of GFP-CpnE was verified with a Western blot using an α-GFP antibody (Ilacqua et al. 2018; Santa Cruz sc-9996).
Under Agar Chemotaxis Assay
Glass-bottom plates (35 mm) were incubated with BSA (10% in water) for 8 minutes, rinsed, and allowed to dry. Agarose (1%) was added to SM media (1% glucose, 1% proteose peptone, 0.1% yeast extract, 0.05% MgSO 4 , 0.19% KH 2 PO 4 , 0.06% K 2 HPO 4 , pH 6), melted, and then 2 mL was added to each plate. After solidification, a rectangular trough in the center of the plate was cut out of the agarose and two holes were made 5 mm from the trough with a plastic straw. NC4A2/GFP-CpnE cells were harvested from plates and centrifuged at 1500 rpm for 5 minutes. Cells were resuspended in SM media at 5x10 ^6^ cells/mL and 30 µL was added to each hole in the agarose plate. A 100 mM folic acid stock was prepared and 100 µL of the stock was added to 900 µL of warmed SM agarose. The mixture was added to fill the rectangular trough. A coverslip was placed over the center of the plate to cover holes. Migrating cells were imaged 2-8 hours later.
Random Movement Assay
NC4A2/GFP-CpnE cells were harvested from plates, counted using a hemocytometer, and centrifuged at 1500 rpm for 5 minutes. The cells were resuspended in VL6 at 2x10 ^6^ cells/mL. In 35 mm glass bottom dishes, 1 mL of cells was mixed with 1 mL of VL6. Cells were incubated at room temperature for 5-6 hours. Immediately before imaging, VL6 was replaced with developmental buffer ((DB) 5 mM Na 2 HPO 4 , 5 mM KH 2 PO _4, _ pH 6.5).
Streaming Assay
NC4A2 and NC4A2/GFP-CpnE cells were harvested from plates, counted using a hemocytometer, and centrifuged at 1500 rpm for 5 minutes. Cells were resuspended in DB at 5x10 ^6^ cells/ml. In 35 mm glass bottom dishes, 1 mL of cells was mixed with 1 mL of DB. Cells were incubated at room temperature for 6 hours. Prior to imaging, DB on plates with NC4A2/GFP-CpnE was replaced with DB containing wheat-germ agglutinin (WGA) Alexa Fluor 555 (Invitrogen W32464) at 25 µg/mL. On plates with NC4A2 cells, DB was replaced with DB containing WGA at 25 µg/mL and/or SYTO RNASelect (Invitrogen S32703) at 1µM.
Confocal Imaging and Analysis
All images were taken with a Nikon A1R laser scanning confocal microscope. GFP-CpnE, WGA-Alexa Fluor 555, and SYTO RNASelect were detected using 488, 561, and 488 nm lasers, respectively. Cells were imaged using the Plan Apo 60X oil-immersion objective with a 1.2 NA. Laser power was consistent between imaging at 8.00. Images were collected at 1024 x 1024 pixels. Image averaging was set to 4X, 8X, or 16X to reduce background noise. HV and offset values were utilized to optimize individual images.
Reagents
Table 1. Cell Strains and Plasmids
**: **
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Chen L Ma L Yu L 2019226 WGA is a probe for migrasomes.Cell Discov 52056-5968131310.1038/s 41421-018-0078-230820350 PMC 6389894 · doi ↗ · pubmed ↗
- 2Fey P Dodson RJ Basu S Chisholm RL 2013 One stop shop for everything Dictyostelium: dicty Base and the Dicty Stock Center in 2012.Methods Mol Biol 9831064-3745599210.1007/978-1-62703-302-2_423494302 PMC 3762881 · doi ↗ · pubmed ↗
- 3Han Y Yu L 2024422 Calcium ions promote migrasome formation via Synaptotagmin-1.J Cell Biol 22380021-952510.1083/jcb.20240206038647453 PMC 11035859 · doi ↗ · pubmed ↗
- 4Huang Z Wang M Chen Y Tang H Tang K Zhao M Yang W Zhou Z Tian J Xiang W Li S Luo Q Liu L Zhao Y Li T Zhou J Chen L 2025120 Glioblastoma-derived migrasomes promote migration and invasion by releasing PAK 4 and LAMA 4.Commun Biol 81919110.1038/s 42003-025-07526-w 39833606 PMC 11747271 · doi ↗ · pubmed ↗
- 5Ilacqua AN Price JE Graham BN Buccilli MJ Mc Kellar DR Damer CK 2018716 Cyclic AMP signaling in Dictyostelium promotes the translocation of the copine family of calcium-binding proteins to the plasma membrane.BMC Cell Biol 191131310.1186/s 12860-018-0160-530012091 PMC 6048903 · doi ↗ · pubmed ↗
- 6Jiang D Jiang Z Lu D Wang X Liang H Zhang J Meng Y Li Y Wu D Huang Y Chen Y Deng H Wu Q Xiong J Meng A Yu L 201981 Migrasomes provide regional cues for organ morphogenesis during zebrafish gastrulation.Nat Cell Biol 2181465-739296697710.1038/s 41556-019-0358-631371827 · doi ↗ · pubmed ↗
- 7Jiao H Jiang D Hu X Du W Ji L Yang Y Li X Sho T Wang X Li Y Wu YT Wei YH Hu X Yu L 2021527 Mitocytosis, a migrasome-mediated mitochondrial quality-control process.Cell 184110092-867428962910.e 1310.1016/j.cell.2021.04.02734048705 · doi ↗ · pubmed ↗
- 8Kessin, R. 2001. Dictyostelium : Evolution, Cell Biology, and the Development of Multicellularity. Cambridge University Press.