Mechanistic insights in the role of trehalose transporter in metabolic homeostasis in response to dietary trehalose
Bhagyashri Y Chaudhari, Vikram J Nichit, Vitthal T Barvkar, Rakesh S Joshi

TL;DR
This study explores how the sugar transporter HaST46 in insects helps regulate trehalose levels in response to diet.
Contribution
The study identifies HaST46 as a key transporter modulating trehalose homeostasis through diet-dependent regulation.
Findings
HaST46 is localized in the posterior midgut and its expression changes with dietary trehalose.
HaST46 preferentially transports trehalose over glucose in Sf9 cells.
Dietary trehalose reduces endogenous trehalose synthesis gene expression.
Abstract
Trehalose is the major sugar in insect hemolymph and plays a diverse role. Its level is regulated by the dynamics of biosynthesis and distribution by sugar transporters. The metabolic balance between trehalose synthesis and uptake remains poorly understood, despite its critical role in homeostasis. Here, we examined the role of the putative gut-specific sugar transporter, HaST46, in regulating trehalose levels in Helicoverpa armigera, a model Lepidopteran pest. Analysis of publicly available transcriptomics, proteomics data, and qRT-PCR study suggests that HaST46 showed localization in the posterior midgut and its expression alters in response to dietary contents. The liquid chromatography-mass spectrometry (LC-MS) analysis of HaST46 overexpressing Sf9 cells indicated that trehalose transport is preferred over glucose. HaST46 expression was found to be modulated tissue-specific manner…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
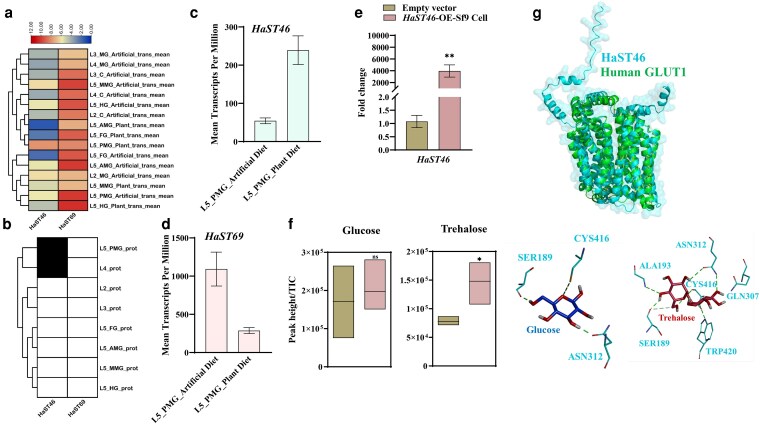 Fig. 1
Fig. 1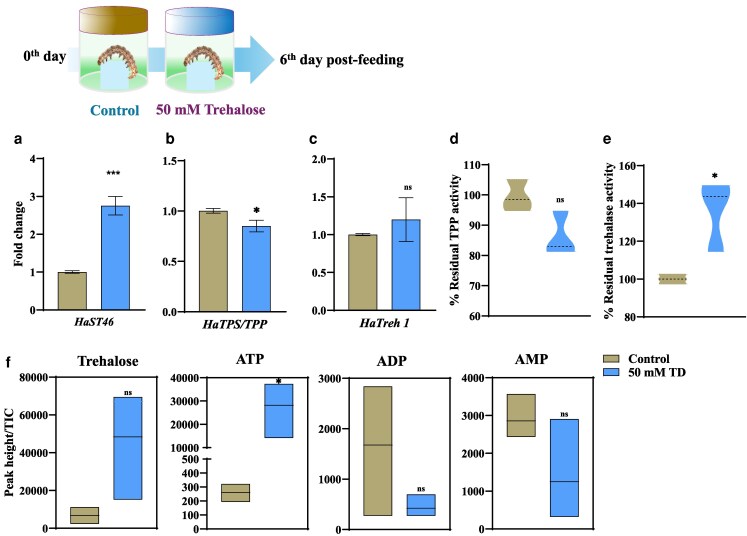 Fig. 2
Fig. 2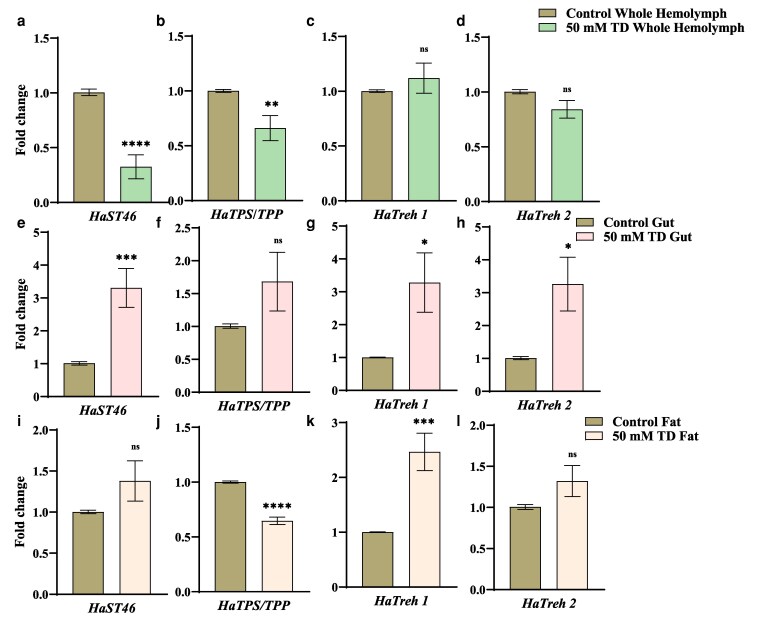 Fig. 3
Fig. 3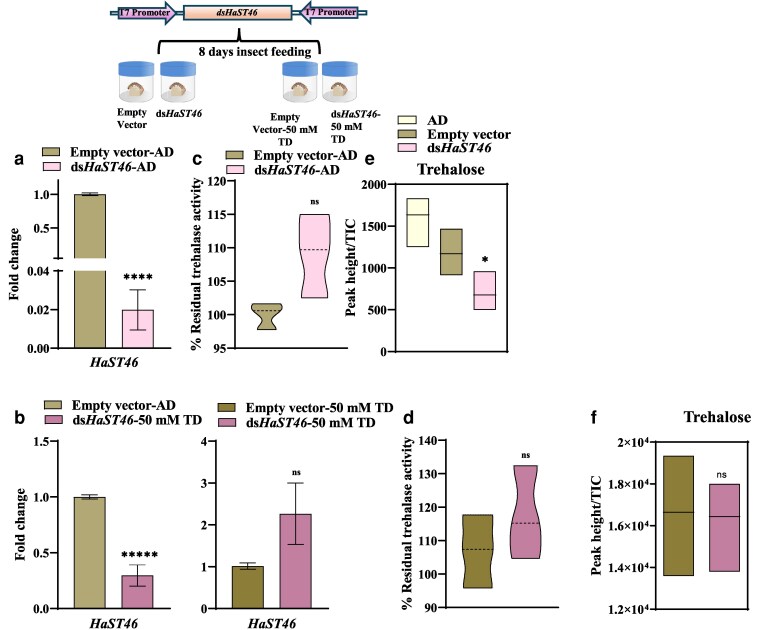 Fig. 4
Fig. 4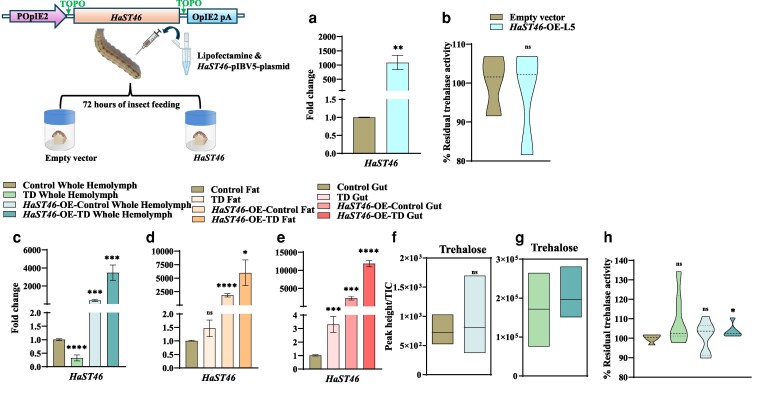 Fig. 5
Fig. 5- —CSIR-UGC-NET
- —ANRF10.13039/100000964
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsNeurobiology and Insect Physiology Research · Insect-Plant Interactions and Control · Insect Resistance and Genetics
Introduction
Insects exhibit a remarkable metabolic adaptation that contributes to their ecological success (Terblanche and Lehmann 2025). One of the key reasons for their extensive adaptability is their metabolic dynamics and variability in response to different stimuli. As carbohydrates are primary energy source for fueling flight, development, reproduction, diapause, and stress tolerance, modulation in their level plays critical role in insect survival (Thorat et al. 2012; Feofilova et al. 2014). Specifically under stress conditions, insects precisely regulate their energy acquisition, storage, and mobilization (Zhang et al. 2019). Among different sugars, trehalose acts as predominant hemolymph sugar, fulfilling both structural and metabolic functions (Murphy and Wyatt 1965; Becker et al. 1996; Shukla et al. 2015; Matsushita and Nishimura 2020). Trehalose metabolism and distribution are essential for insect growth and adaptability (Tellis et al. 2023b). Trehalose is primarily synthesized in the fat body through a conserved pathway involving trehalose-6-phosphate synthase (TPS) and trehalose-6-phosphate phosphatase (TPP) (Murphy and Wyatt 1965; Becker et al. 1996). Following synthesis, trehalose is transported into the hemolymph and then distributed to the target tissues as per the energy demand through sugar (STs) and trehalose transporters (TRETs) (Tellis et al. 2023a).
Insects can ingest different foods to obtain complementary nutrients during nutritional stress (Raubenheimer and Jones 2006; Koyama et al. 2020). They further optimize energy utilization through metabolic balancing, enabling adaptation to diverse dietary conditions (Chown and Gaston 1999; Smith et al. 2018). Trehalose synthesis requires a continuous supply of ATP and carbon skeletons from glycolysis, glycogenolysis, or gluconeogenesis (Miyamoto and Amrein 2017). Depending on resource availability, insect may switch in between nutrient absorption or biosynthesis or differential distribution (Izadi 2025). Hemolymph trehalose levels range from approximately 5 to 50 mM, which is maintained by its continuous biosynthesis, degradation, and distribution (Murphy and Wyatt 1965; Kikawada et al. 2007; Al Hinai et al. 2025). TRETs serve as key molecules in regulating metabolic trade-offs under controlled and stressed conditions (Leyria et al. 2020). The midgut is the primary site of sugar and nutrient absorption, detoxification, and enzymatic conversions (Huang et al. 2015). To meet the metabolic demand, insects rely on sugar absorption or distribution from the gut tissues by sugar transporters such as TRETs, sodium-glucose co-transporters (SGLTs), and facilitative glucose transporters (GLUTs) (Chaudhari et al. 2025).
Phytophagous insects such as Helicoverpa armigera can acquire trehalose directly from the diet (Shukla et al. 2015; Tellis et al. 2023b). In H. armigera, trehalose-specific sugar transporters (HaSTs) showed diversity and spatio-temporal expression patterns (Tellis et al. 2023a). In dipteran and hymenopteran insects, glucose absorption is primarily mediated by SGLTs and GLUTs (Wang et al. 2016). Sugar transporter activity is closely linked to the insulin and TOR signaling pathways, facilitating metabolic plasticity by coupling nutrient availability with growth regulation (Riera et al. 2016). This critical trait allows insects to cope with environmental stressors, shift between trophic modes, and optimize energy expenditure during metamorphosis or overwintering (Danks 2007). The interplay between trehalose metabolism and other physiological systems, including neuroendocrine signaling, symbiotic relationships, and hormonal regulation, facilitates nutrient sensing, energy homeostasis, and ecological adaptability (Tellis et al. 2023b).
Despite advances in understanding trehalose metabolism within insects, the STs’ role and dynamics in response to dietary trehalose remain poorly understood. H. armigera serve as valuable models for metabolic studies due to their high dietary adaptability, rapid development, and stress resilience (Riaz et al. 2021). Elucidating role STs in regulation of trehalose metabolism in response to diet offers insights into the metabolic homeostasis and its impact on physiological processes (Dawkar et al. 2016; Zhou et al. 2023; Guan et al. 2024; Tian et al. 2025). Here, we shed light on the functional role of the trehalose transporter, HaST46, in mediating the metabolic balance between trehalose's exogenous uptake and endogenous synthesis. Through differential gene expression analysis, quantitative real-time PCR (qRT-PCR), liquid chromatography–mass spectrometry (LC-MS)–based metabolomics, enzyme activity assays, and genetic manipulation, we investigated how dietary trehalose influences HaSTs dynamics and activity, mainly in the gut. This study advances the knowledge of sugar transport, metabolic homeostasis. and nutritional physiology.
Materials and methods
Differential gene expression and proteomic signature analysis
Publicly available H. armigera gut transcriptomic and proteomic datasets (Ioannidis et al. 2022) were used for expression profiling of HaSTs. In that study, transcriptome sequencing was performed on whole midguts from artificial diet-fed L2, L3, and L4 larvae along with their corresponding carcass samples. The midgut portion of these samples was also analyzed via proteomics. For L5 larvae, RNA-seq and complementary proteomics data were generated across 5 gut compartments such as foregut (FG), anterior midgut (AMG), middle midgut (MMG), posterior midgut (PMG), and hindgut (HG), and for both plant-fed and artificial diet-fed larvae with these available datasets, we evaluated gut-specific expression and proteomic occurrence of the chosen putative trehalose transporters (HaSTs) based on our previous study (Tellis et al. 2023a). For transcriptomics, raw fragments per kilobase of transcript per million mapped reads (FPKM) values were extracted, log₂(x + 1) transformed, and row-wise Z-score normalized before hierarchical clustering using Euclidean distance and complete linkage in the pheatmap package (R v4.x). For proteomics, the absence or presence of HaSTs in the proteomics set was denoted in binary form. Heatmaps were generated using TBtools 1.09 (Chen et al. 2020) to visualize stage- and tissue-specific expression patterns. For bar plots, mean FPKM values ± SEM were calculated across biological replicates.
HaST46 overexpression in Sf9 cells
For overexpression, HaST46 was cloned into the pIBV5 vector and sequence-confirmed. Sf9 cells (4 × 10^6^) were seeded in a T25 cell culture flask (Corning, NY, USA) with 1 mL serum-free SF900 II SFM media (Thermo Scientific, Waltham, MA, USA) and allowed to adhere by incubating for 30 min at 27 °C. For transfection, 1.5 µg of either the EGFP_pIBV5 control plasmid or the HaST46-pIBV5 plasmid was diluted in 100 µL SF-900 II SFM (without antibiotics). Separately, 8 µL Cellfectin II reagent (Gibco, Waltham, MA, USA) was diluted in 100 µL Sf-900 II SFM and incubated for 30 min. The DNA and lipid solutions were combined (total 200 µL) and incubated for an additional 30 min to allow complex formation. The transfection mix was added dropwise while gently rocking the flask for an even distribution. Flasks were incubated for 5 to 7 h at 27 °C. After the incubation, the media was removed, and fresh Sf-900 II SFM containing blasticidin (Gibco, Waltham, MA, USA) antibiotic was added and incubated for 48 h. Overexpression was confirmed by qRT-PCR. For the sugar uptake assay, 5 and 10 mM concentrations of glucose and trehalose were added to the media, and the cells were incubated for varying time points (0, 5, and 10 min). Following incubation, the media and cells were collected for metabolite extraction and analysis. Sugar uptake was determined from changes in extracellular and intracellular sugar concentrations by metabolite profiling.
Structural modeling and docking
In order to identify substrate-transport preference, SPOT analysis on the DeepMolecule server (https://www.deepmolecules.org/SPOT) was performed. Furthermore, the 3-dimensional structure of HaST46 was predicted using the Alphafold 3 server (https://alphafoldserver.com/) (Supplementary Information 1). Structural alignment of the best model was performed using PyMOL (The PyMOL Molecular Graphics System, Version 3.0 Schrödinger, LLC) using Human Glut1 (PDB ID: 4PYP) structure as reference. Binding site residues of Glut1 were used to predict the binding pocket of HaST46 and molecular docking of glucose and trehalose was performed using AutoDock Tool as described earlier (Barbole et al. 2024).
Insect rearing
Larval populations of H. armigera were obtained from the ICAR-National Bureau of Agricultural Insect Resources (NBAIR), Bengaluru, India, and maintained on a chickpea-based artificial diet (AD) for multiple generations (Supplementary Information 2). Insects were reared at 25 ± 1 °C, 70 ± 10% relative humidity, and a 16:8 h light: dark photoperiod. Adult mating boxes, containing equal numbers of males and females, were provided with 10% (w/v) sucrose solution and 1% (w/v) vitamin E, with a muslin cloth for egg collection (Dawkar et al. 2016).
Exogenous trehalose feeding and tissue collection for the bioassay
For exogenous trehalose feeding in the H. armigera bioassay, freshly moulted second instar larvae were fed AD supplemented with 10 mM, 50 mM, or 100 mM trehalose for 6 d. The components used were same to those described in the above table of artificial diet (AD), with the only modification being the addition of different trehalose concentration (10, 50 and 100 mM to the mixture for feeding assay. Then assess the impact on insect weight, nutritional indices efficiency of conversion of ingested food (ECI), efficiency of conversion of digested food (ECD), and approximate digestibility (AD) and expression of sugar transporter genes (HaSTs) (Supplementary Information 3). Three biological replicates (n = 10 larvae each) were used, and the samples were flash-frozen in liquid nitrogen and stored at −80 °C. For tissue collection, fifth instar larvae were starved for 1 to 2 h followed by washing with phosphate-buffered water and dissected under RNase-free conditions to collect hemolymph, gut tissues (FG, midgut, HG), and fat body. The samples were flash-frozen and stored at −80 °C.
In vitro targeted putative sugar transporter gene HaST46 silencing via dsRNA feeding
Silencing of the HaST46 gene was performed using microbial-based dsRNA production. To generate dsRNA targeting HaST46, a 400-bp fragment was PCR amplified using Phusion DNA Polymerase (Thermo Fisher Scientific, Waltham, MA, USA) and a Proflex PCR machine (Thermo Fisher Scientific, Waltham, MA, USA) with primers (Supplementary Information 4). PCR conditions included 98 °C for 30 s, followed by 35 cycles of 98 °C for 10 s, 60 °C for 30 s, and 72 °C for 30 s, with a final extension at 72 °C for 10 min. The purified PCR product was cloned into the pGEM-T Easy vector (Promega Corporation, Madison, WI, USA), excised, and ligated into the L4440 vector using NotI and EcoRI. The dsHaST46-L4440 plasmid was transformed into E. coli HT115 competent cells. A single colony was cultured in a 5 mL primary culture with tetracycline and ampicillin. A 200 mL 2 × YT media with antibiotics was inoculated for secondary culture and induced with 1 mM isopropyl β-D-1-thiogalactopyranoside (IPTG) at an OD of 0.4. After 4 to 5 h at 37 °C, the cells were harvested, resuspended in nuclease-free water (OD 2.0), and mixed into the larval feed. A bioassay with 15-s instar larvae per treatment was conducted for 6 d using an empty L4440 vector as a control. The feed was replaced, and the parameters were recorded every alternate day. On day 6, 7 larvae per treatment were starved for 2 to 3 h, flash-frozen, and stored at −80 °C. Nutritional indices (ECI, ECD, ADI) and body weight changes were calculated. RNA was extracted using TRIzol, treated with RQ1 RNase-free DNase, and visualized on a 1.2% agarose gel to confirm dsHaST46 production. qRT-PCR was performed to assess HaST46 expression in the control and treated insects, using actin as a reference.
Transient overexpression of HaST46
For overexpression, HaST46 was cloned into the pIB/V5 vector (Thermo Fisher Scientific, Waltham, MA, USA) and sequence-verified. Larvae were starved for 3 h before injection. Further process was carried out according to the procedures described in the Supplementary Method 3 in Supplementary Information 4. A mixture of 1500 ng of each plasmid and Lipofectamine (Invitrogen, Thermo Fisher Scientific Inc., Waltham, MA, USA) (1:1 v/v), incubated for 30 min, was injected into the hemocoel between the fifth and seventh abdominal segments. The EGFP pIB/V5 plasmid served as the control (Barbole et al. 2024). Gene expression was quantified by qRT-PCR at 48 and 72 h post-injection, with 6 biological replicates (3 larvae per replicate). For tissue-specific expression analysis, the tissues were dissected 48 h after injection. Following injection, larvae were maintained on AD supplemented with 50 mM trehalose or control AD for 24 h, then flash-frozen, and stored at −80 °C (Supplementary Information 2).
Quantitative real-time PCR (qRT-PCR) analysis
Total RNA was extracted from 80 to 100 mg of insect cells/tissues using the TRIzol reagent (Invitrogen, Waltham, MA, USA). To eliminate DNA contamination, RNA was treated with RQ1 RNase-free DNase. First-strand cDNA was synthesized from 5 µg RNA using oligo-dT primers and the High-Capacity cDNA Reverse Transcription Kit (Applied BioSystem, Foster, CA, USA), following the manufacturer's protocol. Gene expression was quantified using a 7500 Fast Real-Time PCR System (Applied Biosystems, Foster, CA, USA) with Takara TB Green Premix Ex Taq II. Primer list is provided in Supplementary Information 4. Expression dynamics by calculating fold change (2^−ΔΔCt), were visualized using GraphPad software. The expression of 5 HaSTs, trehalose-6-phosphate synthase (TPS), trehalose-6-phosphate phosphatase (TPP), and trehalase was assessed in trehalose-fed tissues, with the gut as the reference for fold change (2^−ΔΔCt) in fifth instar larvae. HaSTs expression was further evaluated in hemolymph, fat body, and silencing assays, using control expression as the reference.
Metabolite analysis using LC-Orbitrap-MS
To extract metabolites, 80 to 100 mg of finely crushed sample powder was mixed with 500 µL of 80% MS-grade methanol to ensure efficient metabolite extraction (J.T. Baker, Center Valley, PA, USA). Then this mixture vortexed for 15 to 20 min at room temperature to enhance homogenization, then proceed for 20 min sonication to disrupt cellular content followed by centrifugation at 18,000 × g for 10 to 20 min to get clear supernatant. The supernatant stored in −80°C deep freezer to stabilize metabolites and prevent further degradation. The next day, samples were again centrifuged at high speed for 10 to 20 min to remove of residual particles and supernatant filtered through a 0.2 µm syringe filter to remove micro-particles that may interfere with detection. The LC method started with 2% B for the first 0.3 min that is the conditions the column for metabolite separation and for increase compound resolution solvent strength increased to 30% in the next 2 min. The B% was increased from 30% to 45% till 5 min and further increased to 98% to 7 min at which it was held for the next 50 s. The column was equilibrated to the initial ratio of solvents (98% A: 2% B) in the last 2 min to restores initial conditions for the next run. The MS data from 2 or 3 independent biological replicates and 2 technical replicates each were acquired in 5 GHz extended dynamic range (Supplementary Information 2).
Enzymatic activity
The α,α-trehalase activity was determined by measuring the amount of glucose released from the hydrolysis of α,α-trehalose using the dinitrosalicylic acid (DNSA) reagent (Sigma-Aldrich, Merck KGaA, St. Louis, MO, USA). A premix containing buffer and equal volumes of crude sample extracts from the control and treatment groups were prepared. To this reaction, 150 μL of trehalose, i.e. 0.25%, was added. The reaction was allowed to hydrolyze the enzyme for 15 min at 37 °C. The reaction was then stopped with addition of 500 μL of DNSA reagent. The mixture was then placed in a boiling water bath for 5 min in order to enhance the detection of glucose through colorimetry. The quantitative measurement of glucose released during the trehalose hydrolysis was single at 540 nanometers. The absorbance of the measurement was retrieved at 540 nm. Under the defined conditions of the assay, 1 unit of trehalase activity was defined as the amount of enzyme liberated of 1μM of glucose per minute.
The enzyme activity of trehalose 6-phosphate phosphatase (TPP) was assessed by monitoring the release of inorganic phosphate (Pi) from the crude extract using the malachite-green reagent (Sigma-Aldrich, St. Louis, MO, USA). The assay was conducted following the modified protocol of (Klutts et al. 2003 ). The reaction was performed in a total volume of 100 µL with a final concentration of 1 mM trehalose 6-phosphate (Sigma-Aldrich, MA, USA), 2 mM MgCl₂ (Hi-Media, MS, India), 50 mM citrate-phosphate buffer (pH 4), and 35 μg of enzyme extract. The reaction mixture was then incubated at 57 °C for 35 min to allow for hydrolysis. The Klutts procedure was followed, wherein the reaction was first quenched by the addition of 2 volumes of malachite green 0.15% filtered, 1% ammonium molybdate (Hi-Media, MS, India), and 12.5% (v/v) concentrated HCl (Thomas Baker, MS, India). After spectrophotometric measurements of the absorbance at 630 nm, sediment was separated and color terbium titration was held. Color was developed for 5 to 7 min. It can be claimed that this composition effectively measures Pi release.
Statistical analysis
All experiments contain 3 or more biological replicates. Data were expressed as mean ± SE using GraphPad Prism v8.0 (GraphPad Software, San Diego, CA, USA). The Student’s unpaired t-test was used to analyze statistical significance between groups. Asterisks indicate significant changes compared to control (*P < 0.05; **P < 0.01; ***P < 0.001, ****P < 0.0001).
Results
HaST46 showed gut-specific expression at the transcript and protein levels
In our earlier published data we have described expression profiles of 14 HaSTs, out of which 5 HaSTs showed tissue dominant expression pattern (Tellis et al. 2023a ). HaST46 and HaST69 showed highest expression in gut and whole body, respectively (Fig. 1a). We analyzed the gut-specific expression and proteomic profiles of earlier studied 5 HaSTs, using publicly available H. armigera transcriptomic and proteomic datasets (Ioannidis et al. 2022). In the transcriptomics dataset, HaST46 and HaST69 showed high expression levels across larval stages (L2, L3, L4) and gut-region–specific expression in different gut regions, including FG, AMG, MMG, PMG, and HG in fifth instar (L5) larvae. HaST46 expression peaked in the gut during the L4 larval stage and in the PMG of L5 larvae (Fig. 1a). In corroboration with transcript expression of HaST46 and HaST69 in the gut tissues, HaST46 also showed proteomic level presence in the midgut (Fig. 1b). Given that feeding activity peaks during the fourth and fifth larval instars, elevated expression of the STs likely supports increased metabolic demand. Collectively, these data highlight that HaST46 could be putatively involved in sugar absorption in PMG during the active feeder larval stage. Transcriptomics comparisons L5 larvae fed an artificial diet vs those fed a plant (cotton) diet (PD) showed HaST46 expression was elevated in PD-fed larvae fed compared to AD, whereas HaST69 exhibited the opposite trend (Fig. 1, c and d). In combination of proteomic presence, transcript abundance in gut and diet response increase expression of HaST46 suggests its putative role in sugar absorption from feed in the gut. Furthermore, it was important to assess the sugar transport ability of HaST to establish their role in trehalose absorption from gut.
Spatial expression and functional characterization of HaST46. a) Effect of diet on putative trehalose transporters, i.e. HaST46 and HaST69 expression (transcript level; trans) pattern across 5 gut compartments: FG, AMG, MMG, PMG, and HG and carcass (C) in larvae fed on artificial diet (AD) and plant diet (PD). b) Protein abundance (protein level; prot) across larval stages (L2, L3, L4) and gut regions (FG, AMG, MMG, PMG, and HG). c and d) Expression (transcript) levels of HaST46 and HaST69 in the PMG under the 2 dietary regimes. This data analyzed using publicly available H. armigera transcriptomic and proteomic datasets (Ioannidis et al. 2022). Data represent mean ± SEM of biological replicates. Statistical significance was determined using an unpaired t-test (P < 0.05). e) Verification of HaST46 overexpression by qRT-PCR. f) Quantification of D-glucose and trehalose in cell supernatants and lysates using LC-MS/MS after supplementation of the culture medium with glucose (5 mM, 10 mM) or trehalose (5 mM, 10 mM) at 0, 5, and 10 min intervals. All data are presented as mean ± SEM. Statistical analysis was performed using an unpaired t-test (P < 0.05). g) Structural conservation of HaST46 with human GLUT1 transporter. Molecular interaction of HaST46 with glucose and trehalose.
HaST46 showed a preference toward trehalose transport
In order to understand sugar preference and transport ability we performed functional assays with 10 mM trehalose exposure to Sf9 cells overexpressing HaST46. LC-MS–based trehalose quantification of these cells revealed a significant increase in intracellular sugar accumulation in HaST46-expressing cells compared with the vector control (Fig. 1, e and f). HaST46-expressing cells accumulated ∼1.5-fold more trehalose relative to controls, while glucose uptake exhibited a moderate increase (∼1.2-fold), suggesting a trehalose preference of HaST46. In silico analysis using DeepMolecule (https://www.deepmolecules.org/SPOT) also suggested that HaST46 has more preference to trehalose (0.64) compared to glucose (0.58). Furthermore, the structural alignment of HaST46 with the human GLUT1 transporter (PDB: 4PYP) showed a high degree of conservation in transmembrane domain architecture with RMSD value of 3.67 Å (Fig. 1g). Molecular docking studies further revealed that the binding of trehalose (−6.2 kcal/mol) is lower compared to glucose (−4.7 kcal/mol) with HaST46, indicating trehalose structural preference over glucose for transportation. Molecular interaction studies showed CYS416 and ASN312 of HaST46 formed conserved hydrogen bonds with both glucose and trehalose. At the same time, TRP420 and ALA193 contributed additional interactions that might contribute to stabilize trehalose binding (Fig. 1g). These findings support that HaST46 may functions as a trehalose-preferred transporter, likely contributing to sugar uptake and homeostasis in H. armigera.
Trehalose-rich diet feeding alters HaST46 expression and trehalose metabolism genes
To assess the effect of trehalose feeding on trehalose metabolism and transport, we performed a feeding bioassay with a trehalose-containing diet, followed by the insect's molecular response to the altered diet composition. Exogenous trehalose feeding showed normal growth and development (Supplementary Information 5). qRT-PCR analysis showed that feeding of trehalose-containing diet, induced changes in the HaSTs (Supplementary Information 5 and 6). HaTPS/TPP, HaTreh1and HaST46 expression was altered upon trehalose feeding (Fig. 2a). Furthermore, HaST46 was upregulated, and HaTPS/TPP was downregulated, while only HaTreh1 exhibited a non-significant elevation in expression upon feeding a diet with 50 mM trehalose (Fig. 2, b and c). This alteration in the HaTPS/TPP expression level suggests a putative reduction in the energy-intensive endogenous trehalose biosynthesis and increased hydrolysis of exogenous trehalose for energy need. Notably, HaST69 was downregulated in the 50 mM trehalose diet, whereas in 10 and 100 mM trehalose diet, it showed increased expression. This variable expression of different HaSTs might be due to their differential substrate specificity and spatio-temporal expression profile in response to dietary content (Supplementary Information 5). Residual TPP activity was reduced, while trehalase activity was high in larvae fed the 50 mM TD (Fig. 2, d and e). Metabolomic quantification revealed that the insect fed on the TD showed relatively greater accumulation compared to the control, although the changes were not statistically significant compared to the control (Fig. 2f). TD-fed insects showed an increase in ATP levels, whereas ADP and AMP levels remained unchanged, indicating ATP conservation (Fig. 2f). This evidence suggests a preference for hydrolytic metabolism under elevated dietary trehalose, which is likely to convert absorbed trehalose into glucose for systemic energy needs.
Gene expression and enzymatic activity upon exogenous trehalose feeding in H. armigera. Schematic diagram (upper panel): experimental design showing 2 groups of larvae (control vs trehalose-treated) and sampling timeline a to c) qRT-PCR analysis of HaST46 and trehalose metabolism-related genes following dietary trehalose supplementation. Transcript abundance was normalized to reference genes and analyzed as a fold change (2^−ΔΔCt). d and e) In vivo activity of trehalase and trehalose-6-phosphate phosphatase (TPP) in larvae fed a 50 mM trehalose-enriched diet or an artificial diet. f) Quantification of trehalose, ATP, ADP, and AMP in whole larvae using LC-MS under the same dietary conditions. Data are presented as mean ± SEM, with statistical significance assessed by an unpaired t-test (P < 0.05).
HaST46 showed gut-specific upregulation in response to trehalose feeding
Tissue-specific gene expression was further analyzed in 50 mM TD-fed insects to elucidate the dynamics of trehalose transport. The relative expression of HaST46 and HaTPP was reduced in whole hemolymph (Fig. 3, a and b), while the expression of HaTreh 1 and HaTreh 2 was unaltered (Fig. 3, c and d). The gut showed a significant ∼3-fold increase in HaST46 expression under a 50 mM TD (Fig. 3e), indicating an increase in the need to transport and use excess trehalose across the gut tissue. HaTPS/TPP expression was found to be unchanged, while HaTreh 1 and HaTreh 2 expressions were significantly high (Fig. 3, f to h). HaST46 expression in the fat body (key tissue for trehalose synthesis) was found to be unchanged (Fig. 3i). Tissue-wise expression of other HaSTs also altered in response to trehalose feeding (Supplementary Information 5). Interestingly, the expression of HaTPS/TPP decreased significantly, indicating a reduction in endogenous trehalose biosynthesis (Fig. 3j). HaTreh1 showed significantly higher expression in the fat body, while HaTreh2 displayed a non-significant upregulation in the fat body of trehalose-fed insects (Fig. 3, k and l). These observations suggest that dietary trehalose alters trehalose transport and metabolism-related genes in tissue-specific manner.
Tissue-specific expression of HaST46 and trehalose metabolism genes following trehalose feeding. (a to l ) qRT-PCR analysis showing expression patterns in whole hemolymph, fat body, and gut tissues. Transcript levels were normalized to reference genes and expressed as fold change (2^−ΔΔCt) relative to control. Data represent mean ± SEM, with statistical differences determined by an unpaired t-test (P < 0.05).
Metabolomic profiling of trehalose diet-fed insects reveals trehalose metabolism trade-off
LC-MS analysis was performed for the whole hemolymph of insects fed on an artificial diet (AD) and 50 mM TD diet to quantify systemic metabolic changes. There was increased trehalose level in TD-fed larvae, although this was not significant, particularly in the whole hemolymph (Supplementary Information 5). These results confirm that trehalose supplementation alters gene expression and endogenous trehalose levels in the presence of excessive dietary trehalose in H. armigera.
HaST46 knockdown induces compensatory upregulation of the trehalose anabolism TPS/TPP gene
RNA interference (RNAi) was employed to silence gut-dominant HaST46 expression (Supplementary Information 5). We achieved around >50% reduction in the HaST46 transcript levels in *dsHaST46-*fed larvae compared with the empty vector (Fig. 4a) (Supplementary Information 5). Notably, larvae given the TD diet and treated with HaST46 dsRNA exhibited significantly elevated levels of HaST46 transcripts compared to dsRNA-treated larvae on the AD control diet, suggesting that trehalose consumption can stimulate HaST46 expression despite the presence of dsRNA-mediated knockdown (Fig. 4b, left panel) (Supplementary Information 5). Furthermore, trehalase activity was non-significantly elevated in TD-fed insects compared with AD-fed insects (Fig. 4, c and d). LC-MS analysis revealed a decrease in trehalose levels in the insect body upon silencing HaST46 (Fig. 4e). However, upon trehalose feeding to dsHaST46 insects, the trehalose level was almost the same as in the control (Fig. 4f). These findings highlight the potential role of HaST46 in trehalose transport. The impact of HaST46 silencing on nutritional efficiency was assessed by measuring nutritional indices such as ECI, ECD and, AD in both the control and silenced groups (Naseri et al. 2010). Nutritional indices were almost the same in both groups, such as the control and HaST46 silenced insects (Supplementary Information 5).
Silencing of HaST46 in H. armigera. Schematic diagram (top): outline of dsRNA feeding protocol, control (empty vector), and treatment (dsRNA feeding). a and b) qRT-PCR analysis of HaST46 expression in whole-body samples from control insects (empty L4440 vector) and dsRNA-fed insects, either fed on an artificial diet or an artificial diet supplemented with 50 mM trehalose. c and d) In vivo trehalase and TPP activities under the same conditions. e and f) Trehalose levels quantified in whole insects using LC-MS/MS. All values are mean ± SEM, with statistical analysis performed using an unpaired t-test (P < 0.05).
The expression of other trehalose transporter genes (HaST09, HaST29, HaST64, and HaST69) was examined in dsHaST46-fed larvae to explore potential compensatory mechanisms. Silencing HaST46 resulted in significant downregulation of these genes, suggesting overall impact due to reduced trehalose content upon silencing (Supplementary Information 5). HaST09 and HaST29 exhibited upregulation in exogenous trehalose, whereas HaST69 exhibited downregulated in dsHaST46 larvae fed on TD. However, HaST64 and HaTPS/TPP remain unchanged (Supplementary Information 5). However, the functional significance of these changes requires further investigation (Supplementary Information 5). These results highlight the crucial role of HaST46 in regulating trehalose transport and metabolism.
HaST46 overexpression showed gut tissue prevalence, potentially influencing trehalose uptake
To assess the effect of elevated HaST46 transcript levels on trehalose metabolism and transport across tissues, we did transient overexpression of HaST46 in the whole body. Whole-body transient overexpression of HaST46 was confirmed by qRT-PCR analysis, which showed a >1000-fold higher expression of HaST46 in treated larvae compared with the control (Fig. 5a). HaST46 overexpression in larvae did not significantly alter trehalase activity (Fig. 5b). Further, we observed the upregulation of HaST46 in all 3 tissues (whole hemolymph, gut, and fat) of the HaST46-OE insects upon trehalose feeding compared with the control group (Fig. 5, c to e). However, there was a significant increase in the HaST46 transcript level in a group of insects fed on a trehalose-rich diet in the gut, followed by whole hemolymph and fat. HaST46 overexpression causes increased transcript levels in various tissues, with comparatively higher enrichment in the gut (Fig. 5, c to e). Trehalose levels in whole hemolymph were unchanged in TD-fed HaST46_OE insects (Fig. 5, f and g). At the same time, trehalase activity was also found to be high in TD-fed HaST46_OE insects in whole hemolymph (Fig. 5h). These results suggest that HaST46 facilitates the uptake of dietary trehalose and plays a role in sugar homeostasis and energy balance in H. armigera.
Overexpression of HaST46 in fifth instar larvae of H. armigera. a) qRT-PCR expression of HaST46 in the whole body. b) In vivo trehalase activity in whole-body samples following HaST46 overexpression. c to e) Expression analysis of HaST46 in the whole hemolymph, fat body, and gut tissues under 4 feeding conditions: artificial diet (AD), trehalose-rich diet (50 mMTD), HaST46 overexpression with artificial diet, and HaST46 overexpression with trehalose-rich diet (50 mM). Transcript levels were normalized to reference genes and expressed as fold change (2^−ΔΔCt). f to g) LC-MS/MS-based quantification of trehalose in whole hemolymph after HaST46 overexpression. h) In vivo trehalase activity in whole hemolymph, fat body, and gut tissues samples following HaST46 overexpression. Data are shown as mean ± SEM, with significance assessed using an unpaired t-test (P < 0.05).
Discussion
Trehalose is key in energy homeostasis, functioning as a circulating sugar and substrate for rapid energy mobilization. Functional characterization of HaST46 as a trehalose-preferred transporter with potentially localized to the PMG, principal site for nutrient uptake, highlights its putative function in dietary sugar absorption (Fig. 1, a and b) (Terra et al. 2023). Furthermore, the nutrition-responsive expression patterns of HaST46 suggest that its expression likely depends on the diet composition and larval metabolic demand. The higher expression of HaST46 in artificial or plant diet reflects an adaptive shift in response to trehalose availability in diet (Fig. 1, c and d). This adaptive plasticity aligns with earlier reports of the gut-specific modulation of nutrient transporters in response to dietary compositions in insects (Azzouz et al. 2005), underscoring the ecological and physiological relevance of nutrient transporter regulation.
The substrate preference of HaST46 for trehalose over glucose represents its potential functional specialization (Fig. 1, e and f). Despite structural conservation with GLUT1, HaST46 exhibits a preference for trehalose over glucose, suggesting an evolutionary divergence toward trehalose transport (Fig. 1g). This specificity likely enhances transport efficiency and optimizing trehalose handling in sugar-rich dietary contexts.
At the metabolic level, the changes in trehalose biosynthesis and degradation pathways in response to transporter activity reflect a coordinated homeostatic system. The downregulation of endogenous trehalose synthesis in the presence of abundant dietary trehalose suggests a resource-efficient switch that curtails ATP-intensive biosynthesis in favor of direct trehalose uptake (Fig. 2, a to e). An increased level of ATP compared to a low level of ADP or AMP suggests low demand for ATP hydrolysis in the exogenous supply of trehalose, highlighting energy conservation (Fig. 2f). This trade-off supports the notion that insects prioritize the least metabolically costly route to meet carbohydrate requirements, a trait advantageous for survival in fluctuating nutritional environments.
RNAi-mediated silencing reduces HaST46 transcript abundance and triggers compensatory responses at both transcriptomic and enzymatic levels. The upregulation of HaST46 in dsRNA-treated larvae on the trehalose diet suggests trehalose feeding could trigger a compensatory transcriptional response (feedback upregulation) of HaST46 (Fig. 4, a and b). The upregulation of trehalose biosynthetic genes in HaST46 knockdown suggests a potential compensation for reduced trehalose uptake by enhancing its endogenous production. Furthermore, HaST46 overexpression indicated gut prevalent localization of elevated HaST46 molecules, supporting its preferred localization in the gut (Fig. 5, a and b). Together, these findings highlight role of HaST46 in trehalose uptake in the gut and putative involvement in trehalose metabolism regulation in response to dietary trehalose. To elucidate the role of HaST46 in trehalose metabolism and physiology by integrating RNAi and whole-body overexpressions; consequently, future tissue-specific overexpression to get insights into spatial role of HaST46 in trehalose transport. Given that trehalose metabolism is unique to insects and absent in vertebrates, HaST46 offers a promising molecular target for the selective disruption of energy metabolism in lepidopteran pests. This study focuses on a specific lepidopteran species, potentially restricting the wider relevance of the identified trehalose transport dynamics among insects with different gut region and feeding behaviors. The temporal regulation of transporter expression during different developmental stages was not studied, leaving potential shifts in sugar uptake due to ontogeny unaddressed. While the present study offers an extensive overview of transporter expression throughout the gut, analyses targeting specific regions such as the FG, midgut, and HG may clarify the exact locations of trehalose absorption and reabsorption. Subsequent studies should prioritize clarifying the directional flow and mechanistic pathway of trehalose transport, especially in HaST46, which stands out as a significant transporter candidate. Site-directed mutagenesis along with in silico modeling and functional tests may uncover essential amino acid residues involved in substrate recognition, gating, and ion coupling, offering a molecular-level understanding of how HaST46 facilitates selective trehalose transport. These structure-function analyses would enhance our insight into the regulation of trehalose transport and could help formulate approaches to disrupt energy balance in lepidopteran pests.
Conclusion
According to this study, HaST46 is an important gut-localized trehalose transporter that supports metabolic balance in H. armigera. It plays important role in dietary trehalose absorption as per internal metabolic demand by showing preference for trehalose and its regulated expression depending on the availability of dietary trehalose. Functional study, silencing and overexpression, showed that trehalose levels affected the expression of HaST46 and trehalose metabolism genes, thus indicating a coordinated balance between external absorption and endogenous synthesis. These findings highlight the critical function of HaST46 in controlling trehalose metabolism and demonstrate the potential of this protein as a specific molecular target for disrupting energy regulation in lepidopteran pests. Future structural and functional aspects in depth will highlight the mechanistic importance of HaST46 in trehalose transport.
Supplementary Material
jkaf303_Supplementary_Data
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Al Hinai MS, Rehman A, Siddique KHM, Farooq M. 2025. The role of trehalose in improving drought tolerance in wheat. J Agron Crop Sci. 211:e 70053. 10.1111/jac.70053. · doi ↗
- 2Azzouz H et al 2005. Effects of plant protease inhibitors, oryzacystatin I and soybean bowman–birk inhibitor, on the aphid macrosiphum Euphorbiae (homoptera, aphididae) and its parasitoid aphelinus abdominalis (hymenoptera, aphelinidae). J Insect Physiol. 51:75–86. 10.1016/j.jinsphys.2004.11.010.15686649 · doi ↗ · pubmed ↗
- 3Barbole RS, Sharma S, Patil Y, Giri AP, Joshi RS. 2024. Chitinase inhibition induces transcriptional dysregulation altering ecdysteroid-mediated control of spodoptera frugiperda development. Iscience. 27:109280. 10.1016/j.isci.2024.109280.38444606 PMC 10914475 · doi ↗ · pubmed ↗
- 4Becker A, Schlöder P, Steele JE, Wegener G. 1996. The regulation of trehalose metabolism in insects. Experientia. 52:433–439. 10.1007/BF 01919312.8706810 · doi ↗ · pubmed ↗
- 5Chaudhari BY, Pradhan AG, Joshi RS. 2025. Metabolic gatekeepers: dynamic roles of sugar transporters in insect metabolism and physiology. Insect Mol Biol. 34:1–18. 10.1111/imb.12963.39394882 · doi ↗ · pubmed ↗
- 6Chen C et al 2020. T Btools: an integrative toolkit developed for interactive analyses of big biological data. Mol Plant. 13:1194–1202. 10.1016/j.molp.2020.06.009.32585190 · doi ↗ · pubmed ↗
- 7Chown SL, Gaston KJ. 1999. Exploring links between physiology and ecology at macro-scales: the role of respiratory metabolism in insects. Biol Rev Camb Philos Soc. 74:87–120. 10.1111/j.1469-185X.1999.tb 00182.x. · doi ↗
- 8Danks HV . 2007. The elements of seasonal adaptations in insects. Can Entomol. 139:1–44. 10.4039/n 06-048. · doi ↗