Research Advances and Disease Modeling in Respiratory Organoids
Lanhe Chu, Dian Chen, Simin Jiang, Huanyu Long, Xiaojuan Liu, Yahong Chen

TL;DR
Respiratory organoids are advanced biological models that improve disease research and drug development by mimicking human respiratory systems more accurately than traditional methods.
Contribution
This review highlights recent advances in respiratory organoid models and their transformative potential in studying and treating respiratory diseases.
Findings
Respiratory organoids provide human-specific microenvironments for studying disease mechanisms and drug responses.
They enable functional drug screening and personalized therapy development for respiratory diseases.
Integration with precision medicine could redefine respiratory disease research.
Abstract
Organoid culture represents a sophisticated biological model that surpasses traditional two-dimensional (2D) methods and animal models in physiological relevance and cost-effectiveness. Current organoid systems derive from adult, fetal, and induced pluripotent stem cells, providing innovative platforms for studying organ development, disease pathogenesis, and drug discovery. Recent technological advances now enable respiratory organoids to significantly contribute to respiratory disease research. This review comprehensively synthesizes the development of respiratory organoid models and their applications in studying major respiratory diseases, including pulmonary fibrosis, chronic obstructive pulmonary disease (COPD), and lung cancer. It further evaluates the transformative potential of these models in advancing respiratory disease research. Respiratory organoids uniquely model disease…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
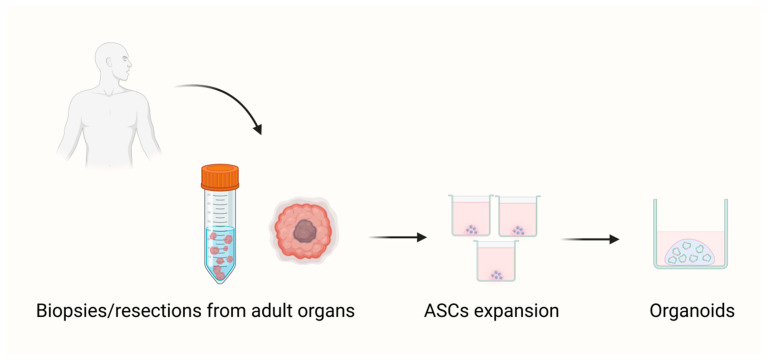 Figure 1
Figure 1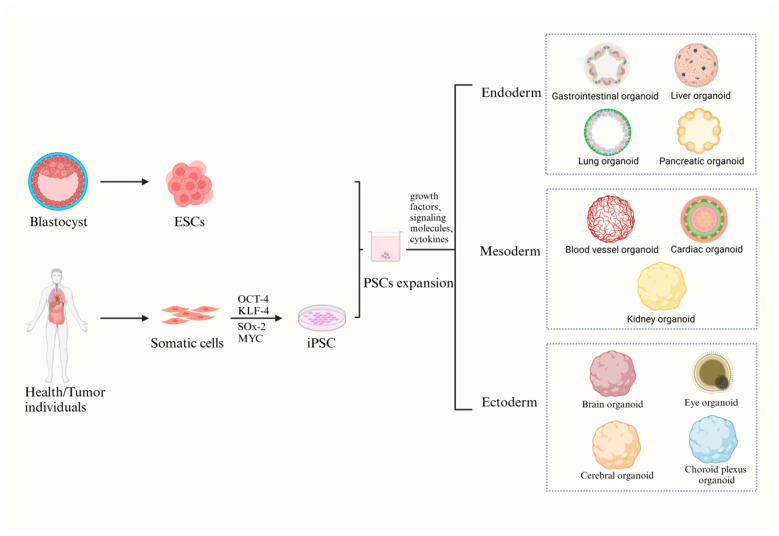 Figure 2
Figure 2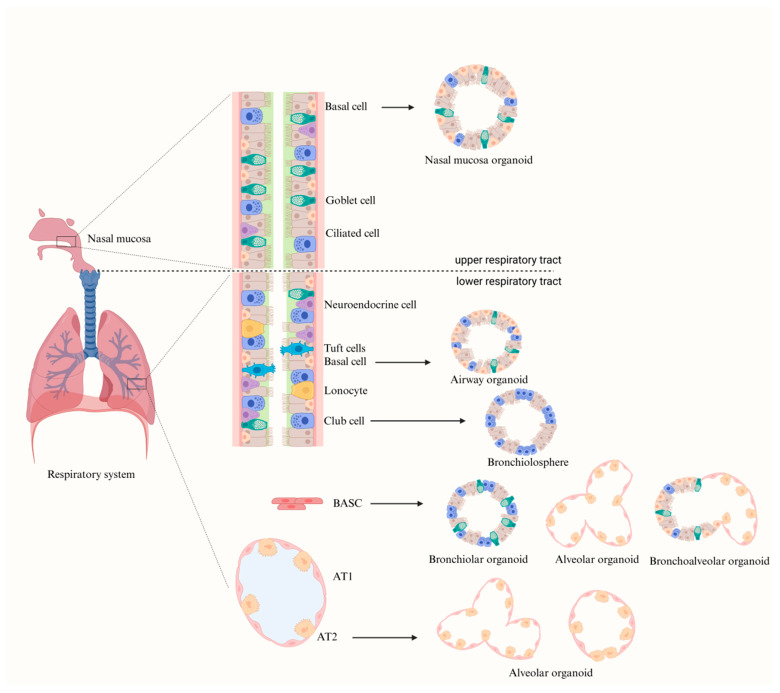 Figure 3
Figure 3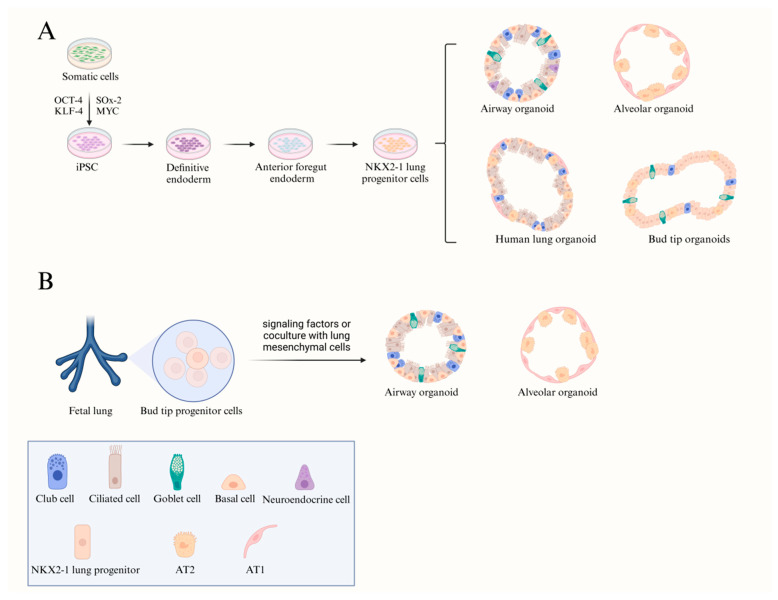 Figure 4
Figure 4- —Natural Science Foundation of China
- —National Key R&D Program of China: “Frontier Biotechnology” Key Special Project
- —National Key Laboratory of Vascular Homeostasis and Remodeling Open Project
- —State Key Laboratory of Natural and Biomimetic Drugs (Peking University)
- —National Natural Science Foundation of China (NSFC) International Cooperation and Exchange Program
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsNeonatal Respiratory Health Research · Cancer Cells and Metastasis · Inhalation and Respiratory Drug Delivery
1. Introduction
Organoids were originally described as aberrant cell growth or intracellular structures in early three-dimensional (3D) cell culture studies [1]. As research proceeded, the concept was expanded to refer to 3D culture systems derived from stem cells or tissue-specific cells that can self-organize and differentiate into structures mimicking the architecture and function of real organs [2]. These “mini-organs” have become powerful tools for exploring human biology, disease mechanisms, and therapeutic strategies. In healthcare, organoids are widely used in disease modeling, drug discovery and screening, precision medicine, and regenerative medicine. In basic research, they facilitate studies of development, organ function, and pathology. By recapitulating key features of native organs, organoids provide an invaluable platform for advancing biomedical science [3].
2D cell/tissue culture models have long been used to study human diseases and development, yet they are limited by their lack of physiological relevance. In vivo, cells exist within complex microenvironments and are influenced by diverse biochemical and mechanical cues—conditions that 2D cultures fail to reproduce [3,4,5,6]. Consequently, cells in 2D cultures gradually lose their native morphology and heterogeneity, and long-term passaging leads to marked genomic and metabolic divergence from the original cell lines. Animal models such as genetically engineered mouse models (GEMMs) and patient-derived xenografts (PDXs) offer improved physiological fidelity but are costly, time-consuming, and limited by interspecies differences [7,8]. Organoid culture systems overcome many of these drawbacks. Derived from stem or tissue-specific cells, organoids retain the genetic and functional traits of their tissue of origin while maintaining long-term stability in vitro [9,10,11]. By mimicking the in vivo microenvironment, they provide a superior platform for studying disease mechanisms and therapeutic responses.
Thus far, three different types of organoids have been described based on the source of their starting material: (a) adult stem cells (ASCs), (b) embryonic stem cells (ESCs), and (c) induced pluripotent stem cells (iPSCs) (Figure 1 and Figure 2).
Both ESC/iPSC and ASC-derived organoids can form 3D organ-like structures that partially recapitulate the architecture and function of native organs, though important differences remain (Table 1). Together, these systems provide complementary platforms for scientific research and personalized medicine. Ongoing efforts aim to establish diverse ESC/iPSC- and ASC-based “human disease-in-a-dish” models.
Respiratory organoids, as an emerging 3D model, offer distinct advantages for studying respiratory biology. Unlike traditional 2D lung cell cultures that form basic epithelial layers with progenitor traits, organoids better capture complex cell interactions, extracellular matrix, and physiological signals [25,26,27]. ALI (Air-Liquid Interface) culture system is a valuable in vitro model that effectively mimics the airway epithelial cell microenvironment [28]. However, it has significant limitations, including a limited source of primary cells, a complex culture process, and low expansion capacity. Primary airway basal stem cells (ABSCs) exhibit poor passaging stability, with their differentiation ability declining after 2–3 passages, which restricts long-term experimentation [29]. Additionally, the model’s complexity and reliance on transwell membranes hinder scalability for large applications [30]. Respiratory organoids derived from ASCs offer a short culture duration and long-term maintenance, closely mimicking adult tissues, which is advantageous for regenerative medicine and chronic disease modeling [31]. In contrast, organoids from iPSCs involve more complex, time-consuming, and costly processes, often yielding less mature, fetal-like cells [31]. Due to their diversity, scalability, and reproducibility, respiratory organoids are promising for studying respiratory disease mechanisms, lung regeneration, and personalized therapies [17,19,32,33]. In this review, we summarize recent advances in respiratory organoid technology and its applications in respiratory diseases, highlighting future opportunities in this rapidly evolving field.
2. Composition of the Respiratory System and Modeling of Respiratory Organoids
The respiratory system is a complex organ system that can be divided into the upper respiratory tract (URT) and lower respiratory tract (LRT). The URT includes the nostrils, nasal cavity, sinuses, nasopharynx, oropharynx, and the portion of the larynx above the vocal cords, while the LRT comprises the larynx below the vocal cords, trachea, bronchi, bronchioles, and alveoli [34]. Its primary physiological function is the exchange of oxygen and carbon dioxide [35]. The respiratory tract exhibits region-specific cellular compositions and physiological characteristics. Airway epithelium from the nasal cavity to the terminal bronchioles consists mainly of four cell types: ciliated, goblet, basal, and Club (Clara) cells. Together, they maintain mucociliary clearance and protect against inhaled particles and pathogens [36,37]. The epithelial cells of the URT form the first barrier against environmental agents and pathogens. Mucus secreted by glands and goblet cells forms the airway surface liquid (ASL), which supports mucociliary clearance; this process is more efficient in the URT due to its higher density of ciliated cells [37,38]. The LRT epithelium is more complex. In addition to the aforementioned cells, secretory cells regulate airway hydration, maintaining proper mucus flow and clearance [39,40]. The alveoli, as the functional units of gas exchange, are composed of type I (AT1) and type II (AT2) alveolar epithelial cells [41,42,43]. AT1 cells cover about 96% of the alveolar surface and form a very thin blood-gas barrier for efficient exchange [44], while AT2 cells secrete surfactant to reduce surface tension and prevent alveolar collapse [41,43]. AT2 cells also function as facultative progenitor cells, capable of self-renewing and differentiating into AT1 cells during repair [45,46].
During embryonic development, respiratory epithelial cells originate from the endoderm and eventually develop into multiple airway compartments, including the alveoli, bronchioles, and proximal airways [47]. These regions contain specific stem and progenitor cell populations, such as basal cells, Club cells, bronchioalveolar stem cells (BASCs), pulmonary neuroendocrine cells (PNECs), and AT2 cells, that collectively sustain repair and homeostasis [47,48,49]. Basal cells are abundant in the bronchi and gradually decrease distally [50,51]. Club cells are primarily located in distal airways and possess secretory functions [52]. Basal cells exhibit strong proliferative capacity and can differentiate into ciliated, goblet, and Club cells after injury [53,54]. Under steady-state conditions, Club cells show limited self-renewal ability but can differentiate into ciliated and goblet cells; in response to viral infection, they can dedifferentiate into basal cells [55,56,57]. Currently, organoids derived from basal cells, including tracheal, bronchial, and nasal organoids, have been successfully established, containing multiple airway cell types [58,59,60]. Furthermore, BASCs at the mouse bronchial-alveolar junction co-express Club cell markers (Club Cell Secretory Protein, CCSP) and AT2 cell marker (surfactant protein C, SPC). These cells possess self-renewal abilities and can differentiate into Club cells, AT1 cells, and AT2 cells under both homeostatic and injury conditions [61,62]. PNECs emerge during early lung development, initially appearing in proximal conducting airways and later in distal branches, characterized by calcitonin gene-related peptide (CGRP) expression [63]. Lineage tracing studies in mice have revealed that PNECs can self-renew and differentiate into Club and ciliated cells following injury, although their removal does not markedly affect regeneration, suggesting additional repair mechanisms exist [64]. A variant form of Club cells (vClub) in the bronchioles can self-renew and replenish lost Club cells [65]. Finally, AT2 cells in the distal lung, owing to their robust self-renewal and differentiation capacities, serve as ideal sources for generating alveolar-like organoids [66].
The specific stem and progenitor cell populations in the respiratory epithelium can generate 3D structures that closely recapitulate in vivo cellular organization and interactions through differentiation or co-culture. These structures include nasal, trachea, bronchi, and alveolar organoids. Currently, three types of stem cells are commonly used for constructing respiratory organoids: ASCs (Figure 3), iPSCs, and fetal lung stem cells (FSCs) (Figure 4). Such models are widely used to study respiratory pathophysiology and to explore novel therapeutic strategies.
2.1. Nasal Mucosa Organoid
Early investigations predominantly utilized traditional cell lines, such as primary human nasal epithelial (HNE) cells, to examine nasal diseases. However, these models often failed to accurately reflect the physiological characteristics of the human nasal epithelium [67]. With advancements in cell culture methodologies, researchers have successfully reconstructed 3D nasal epithelium in vitro, providing new platforms for studying respiratory viral infections and other airway diseases. Liu et al. developed an in vitro organoid model of human nasal epithelial cells using nasal biopsy samples and characterized its differentiation and imaging features [67]. This model has been employed in cystic fibrosis research and other respiratory disease studies. Following this, Brewington et al. [68] and Gamage et al. [69] also utilized nasal biopsy samples and nasal brushings to create 3D models of nasal epithelial cells. However, these methods are invasive, requiring clinical sampling of nasal or airway tissues, which limits their accessibility for routine research and drug screening. To overcome this, Chiu et al. developed a non-invasive two-phase culture system using nasal epithelial cells from inferior turbinate swabs of healthy donors to model SARS-CoV-2 infection [70]. This system involves an amplification phase for long-term expansion (over six months) and a differentiation phase to generate mature nasal organoids or 2D monolayers. The differentiated organoids contained ciliated cells, basal cells, goblet cells, and club cells, and expressed high levels of ACE2, faithfully mimicking native nasal epithelium. Moreover, adjusting the differentiation medium to mildly acidic conditions (pH 6.6) enhanced epithelial barrier formation, closely simulating the physiological nasal surface environment. Another group established a fully non-invasive human nasal mucosa organoid (HNO) model applicable to both pediatric and adult donors [71]. Stem cells isolated from nasal lavage or turbinate swabs were cultured in 3D and differentiated under air–liquid interface (ALI) conditions, forming polarized, pseudostratified epithelia with ciliated, basal, secretory, and goblet cells. The cultures exhibited motile cilia, maintained long-term proliferative capacity, and could be cryopreserved, making them valuable for respiratory disease modeling. An “amplification-differentiation” culture approach has also been developed using human nasal polyp samples [72]. Following sequential amplification (7 days) and differentiation (14 days), this method generated organoids closely resembling normal nasal mucosa, providing a promising tool for studying nasal disease pathogenesis and drug screening [73]. However, conventional organoid cultures using Matrigel limit cell migration and promote inward growth, resulting in apical surfaces being enclosed within spheroids [30,74]. This orientation may not fully recapitulate the in vivo epithelial structure, as the apical surface of airway epithelium should face the atmosphere and interact with the contents of the lumen. To address this, researchers developed a human nasal mucosa organoid model with an outward-facing apical surface using nasal progenitor cells embedded in a composite hydrogel (CAH gel). This system accurately reproduced epithelial polarity, cellular diversity, and revealed that epithelial-derived matrix metalloproteinases (MMPs) regulate epithelial fate and polarity [75]. Since basal cells are also present in nasal epithelium, organoids or “nasal balls” can be derived from these cells, offering a convenient patient-derived platform for drug screening. Nonetheless, organoid characteristics may vary depending on the anatomical origin of the basal cells within the nasal cavity [76].
2.2. Airway Organoid
Nasal organoids, derived from donor nasal epithelial cells, contain major airway epithelial cell types and share similarities with airway organoids. However, nasal mucosa organoids retain the unique characteristics of the upper airway epithelium. In 2016, Satoshi Konishi et al. established the first stepwise 3D differentiation protocol to generate multiciliated airway cells (MCACs) from hiPSCs [77]. Using carboxypeptidase M (CPM) to isolate ventral foregut endodermal cells, they formed 3D progenitor aggregates (‘spheroids’). These airway progenitor spheroids could then be directed to differentiate into a complex 3D epithelium containing MCACs and other airway lineages (excluding alveolar cells). Then the researchers identified optimal culture conditions for proliferating spheroids and inducing FOXJ1 expression (a representative marker for MCACs), which included FGF10, CHIR99021 (a WNT agonist), KGF, and DAPT (a γ-secretase inhibitor that blocks the Notch pathway). This pioneering work paves the way for future applications aimed at modeling airway diseases or developing airway reconstruction methods, such as artificial tracheas. Subsequent studies revealed that high concentrations of FGF-10 could induce the generation of human lung organoids (HLOs) from hiPSCs. These organoids contained airway-like epithelial components marked by TP63 and FOXJ1, as well as mesenchymal and epithelial cells that simultaneously expressed SFTPC and HOPX, which are markers of alveolar cells. In contrast, a culture system based on FGF-7, CHIR, and ATRA could induce lung progenitor cells to form bud-tip-like organoids (BTOs) with airway-like regions expressing SOX2^+^, MUC5AC^+^, and SCGB1A1^+^, as well as tip-like structures expressing SOX2^+^, SOX9^+^, SFTPC^+^, and ID2^+^ [78]. Based on these findings, SOX2^+^, SOX9^+^, CPM^+^^,^ and NKX2-1^+^ tip progenitor cells were directed to form airway organoids [79]. Nikolić and colleagues et al. later developed fetal lung bud tip (LBT)–derived organoid cultures using a combination of growth factors (EGF, FGF7, FGF10), BMP inhibitors (Noggin, SB431542), and WNT activators (RSPO1, CHIR) [80]. These maintained NKX2-1 expression and lung progenitor identity without full differentiation. Subsequent exposure to airway differentiation media or modulation of SMAD signaling (via TGF-β, BMP4, A8301, and Noggin) produced proximal airway organoids containing basal, goblet, Club, and ciliated cells [81].
In 2017, Tan et al. [82] first reported the generation of airway organoids by co-culturing human bronchial epithelial cells, lung fibroblasts, and pulmonary microvascular endothelial cells. They found that randomly seeded mixed cell populations could self-organize into branching epithelial-endothelial structures. In 2018, the same group established the first adult stem cell-derived airway organoids from normal lung tissue. These organoids, composed exclusively of epithelial cells, were capable of stable long-term expansion for over a year in an expansion medium containing key growth factors R-spondin, Noggin, FGF7, and FGF10 [30]. Based on a differentiation protocol previously reported by Satoshi Konishi et al. [77], they induced proximal differentiation in these continuously expanding organoids, thereby generating airway organoids that could mimic the airway epithelium. The team continued to explore distal differentiation systems for alveolar cultures, successfully establishing a respiratory organoid culture system with bidirectional differentiation potential for both proximal and distal airway regions [70]. Additionally, Sachs et al. developed a long-term expanding human airway organoid model from human bronchoalveolar lavage fluid and lung tissue specimens from cystic fibrosis and lung cancer patients [83]. By modulating TGF-β, FGF, and WNT signaling pathways, these basal cell-derived airway organoids contained basal cells, ciliated, and secretory/Club cells, and maintained stability for over a year. They preserved tissue pathological features and cancer gene mutations, making them suitable as in vitro models for malignant tumors and infectious lung diseases. Notably, they further optimized the organoid culture system by removing Noggin from the medium and adding DAPT/BMP4 to the standard airway organoid culture media [84]. Researchers also validated the regenerative and multipotent properties of basal cells in bronchial organoids derived from human bronchial tissue. Another study reported that basal cells derived from human bronchial tissue could differentiate into other cell types, including goblet cells and ciliated cells, in bronchial organoids. This differentiation ability was found to be closely associated with the Notch signaling pathway [85]. Co-culturing Club cells with mesenchymal subpopulations has been employed to evaluate the ability of mesenchymal cells to support stem cell regeneration and differentiation. Under these conditions, co-culturing generated organoids called bronchiolar organoids, which contained both Club and ciliated cells. However, due to the absence of basal and goblet cells, these organoid models are mainly used to study the biological characteristics of Club and ciliated cells [85,86]. Furthermore, mouse BASCs have been shown to co-express both bronchiolar and alveolar cell markers. By co-culturing BASCs with lung endothelial cells, researchers successfully generated bronchiolar and bronchioalveolar organoids, providing new model systems for studying lung structure and function [87].
Current airway organoid models have limitations in simulating the 3D structure and physiological features of small airways. Researchers have addressed this by using cell encapsulation techniques, where bronchial epithelial cells from patients are encapsulated in tubular alginate scaffolds, successfully creating a physiologically relevant bronchial organoid model. This model exhibits typical epithelial morphology and tubular structures with visible lumens, closely mimicking the physiological characteristics of the human distal airway [88]. It also facilitates efficient viral infections via intraluminal injection, overcoming challenges seen with traditional infection methods. Previously, organoid construction relied on lung tissue specimens or iPSCs, which were limited by specimen and complex procedures. A recent study reported generating airway organoids from bronchoalveolar lavage fluid (BALF) [89]. After dissociating and filtering BALF cells, they were cultured in airway or alveolar organoid systems, generating stable organoids within 7–10 days, with passaging maintained for at least 6 months. The system also allows for freezing and recovery. Single-cell sequencing analysis revealed that BALF-derived airway organoids were primarily composed of basal cells, with smaller proportions of secretory/goblet cells, ciliated cells, neuroendocrine cells, and tuft cells. This technique offers a new research strategy for modeling lung diseases using accessible human lung samples. However, organoids grown in extracellular matrices, including airway organoids, typically exhibit apical polarity, with the apical surface enclosed within the organoid lumen. To address this, researchers reversed the polarity in airway organoids by removing the extracellular matrix, creating an “apically outward” model [90]. This model, with its apical orientation, offers a more physiologically relevant approach to studying respiratory infections and airway diseases.
2.3. Alveolar Organoid
In 2013, Barkauskas and colleagues first demonstrated that AT2 cells are the primary stem cells of the alveolus through both in vivo and in vitro experiments. By placing lineage-labeled AT2 cells in a 3D culture environment, they generated an “alveolar layer” with self-renewing capabilities, which contained AT2 cells and cells expressing AT1 markers [66]. Co-culturing with primary PDGFRα-positive lung mesenchymal cells promoted more efficient alveolar layer growth and differentiation [66]. Within this AT2 cell population, Wnt-responsive alveolar epithelial progenitor (AEP) cells play a crucial role in distal lung regeneration after acute injury. These AEPs remain stable during homeostasis but proliferate rapidly post-injury to regenerate much of the alveolar epithelium. Further studies identified TM4SF1 as a surface marker for human AEP cells, aiding the identification of AEP subpopulations in normal human lungs [91]. Co-culture with human fetal lung fibroblasts enabled these subpopulations to generate functional alveolar layers, offering insights into human alveolar regeneration and disease modeling [91]. Additionally, the team also developed immortalized human AT2 cell lines using SV40 large T antigen lentiviral transfection and a RHO/ROCK pathway inhibitor (Y-27632), which formed alveolar organoids when co-cultured with mouse lung fibroblasts [25]. Mesenchymal stem cells (MSCs) secreted factors like TSP1 and MMP-9, which regulated lung progenitor cell function and promoted alveolar differentiation in co-culture with lung progenitor organoids [92]. Karen Hoffmann and colleagues [93] identified a subpopulation of HTII-280^+^/EpCAM^+^ alveolar epithelial progenitor cells in adult lungs using single-cell RNA sequencing. These cells, enriched in stem cell markers, generated dual-differentiated bronchioalveolar organoids in 3D culture, with their differentiation direction influenced by GSK-3β inhibition (CHIR99021), which promoted alveolar differentiation via WNT signaling [93,94,95]. Recently, a method was developed to generate AT2-derived alveolar organoids without fibroblasts [95]. Specifically, by using Matrigel containing Jagged-1 and a specific mixture of medium containing FGF7, Noggin, SB431542, and CHIR99021, alveolar organoids derived from human AT2 cells were successfully established. These advances deepen our understanding of alveolar biology and provide models for studying lung diseases and regenerative processes.
In addition, BASCs have been shown to regenerate both bronchiolar and alveolar epithelia under various injury conditions [61,96]. Endothelial cell-derived BMP4/NFATc1/TSP1 signaling enhances BASC proliferation and differentiation into an alveolar phenotype [87]. Co-culturing BASCs with lung endothelial cells forms bronchioalveolar organoids, but these lack AT1 cells in distal alveolar regions [87]. Adding lung mesenchymal cells to the culture generates organoids with mature alveolar-like and proximal airway-like regions. The distal regions express AT1 and AT2 markers (SFTPC, HOPX), while proximal regions show basal, goblet, and ciliated cell markers (TP63, FOXJ1, MUC5AC). Introducing alveolar macrophages into organoids aids differentiation and maturation, creating a complex niche. This model provides a new platform for studying lung infections, injuries, and repair [97]. Further research also reported that lung alveolar organoids can be derived from basal cells. Salahudeen et al. [98] cultivated organoids from Krt5^+^ basal cells, and single-cell sequencing revealed that these organoids had proliferating ITGA6+ and ITGB4+ cells, with TNFRSF12A^hi cells showing high clonogenic activity. This offers crucial insights into the source and culture optimization of distal lung alveolar organoids.
In recent years, significant progress has been made in the development of alveolar organoids derived from ESCs and iPSCs. Researchers have successfully constructed organoids derived from fetal lung tip (LBT) cells in a culture medium containing FGF-7, FGF-10, CHIR, DCI (dexamethasone, cAMP, and intracellular cAMP activator IBMX), T3 (triiodothyronine), and the Notch inhibitor DAPT. Through co-culturing these LBT-derived organoids with freshly isolated human lung mesenchymal cells, differentiation of the organoids towards the alveolar lineage was induced [80]. Furthermore, when tip progenitor cells sourced from fetal lung tissue (from 8.5 to 19 weeks of gestation) were cultured with isolated RSPO2^+^ mesenchymal cells, RSPO2 signaling was observed to facilitate the maintenance of the tip and its differentiation towards the alveolar lineage. The enhanced RSPO2^−^ mediated signaling supports distal (alveolar) differentiation of the tip organoids via WNT signaling pathways [81]. Dye et al. first reported human lung organoids (HLOs) from hPSCs, using SAG to activate HH signaling, boosting NKX2.1 expression and forming organoids with both proximal and distal airway structures [99]. These organoids included multiple alveolar cell types, effectively mimicking human lung structure. Later studies used ESCs/iPSCs to create alveolar organoids with diverse epithelial cells like AT1, AT2, and basal cells [100]. Culturing these organoids under controlled oxygen and pressure conditions and using co-cultures has led to complex, multi-cellular structures resembling in vivo alveoli [101]. For example, iPSC-derived alveolar epithelial cells (iAECs) and their co-cultures with macrophages have been used to model the capillary-alveolar barrier [102]. Seo et al. [103] created human iPSC-derived alveolar organoids with macrophages (iMACs) by injecting iMACs into human hiPSC-derived alveolar organoids. These iMACs, which persisted for up to 14 days, showed increased IL-1β and TNF-α gene expression under LPS-induced inflammation. Lung mesenchyme provides structural support for lung tissue and facilitates differentiation. Yamamoto et al. proposed embedding alveolar organoids in extracellular matrix hydrogels with fetal lung fibroblasts, which supported SFTPC^+^ alveolar organoid expansion and AT2 stemness [104]. Tamai et al. [105] used NKX2-1^+^ lung progenitor cells and iPSC-derived mesenchymal cells (iMES) to generate alveolar organoids. iMES, expressing R-spondin-2 and R-spondin-3, promoted differentiation of SFTPC^+^ EPCAM^+^ AT2 cells, emphasizing epithelial–mesenchymal interactions in lung organogenesis. By optimizing culture medium components like CHIR99021, FGF10, KGF, and DAPT, researchers have successfully established a long-term expansion system for iPSC-derived alveolar organoids [104]. Additionally, customized hyaluronic acid hydrogels with micro-chambers have been used to produce uniform alveolar organoids from human iPSC-derived AT2 cells in a matrix-free system. These ESC/iPSC-derived alveolar organoid models offer valuable platforms for studying lung development, lineage tracing, and cell interactions in disease processes [106].
3. Applications of Respiratory Organoid Models to Study Lung Diseases
3.1. Pulmonary Fibrosis
Respiratory organoids derived from human ESC/iPSC and ASC can simulate key features of pulmonary fibrosis, including idiopathic pulmonary fibrosis (IPF) and cystic fibrosis (CF) [107]. Although significant breakthroughs have been made in these studies, clinical treatment options for pulmonary fibrosis remain limited. Organoid technology has played an important role in uncovering disease mechanisms and advancing drug development.
3.1.1. Idiopathic Pulmonary Fibrosis
Idiopathic pulmonary fibrosis (IPF) is a chronic interstitial lung disease primarily associated with alveolar epithelial cell injury, aberrant fibroblast activation, and excessive extracellular matrix (ECM) deposition. By using CRISPR/Cas9-based gene editing, TGF-β1, or bleomycin stimulation, researchers have generated IPF-like organoids that display fibroblast proliferation, senescence, and ECM accumulation [108]. For example, Strikoudis et al. developed HPS1^−/−^ organoids that spontaneously developed fibrosis, while Kim et al. established a TGF-β1–induced hPSC-derived alveolar organoid model for drug testing [108,109]. Chen et al. constructed lung bud organoids (LBOs) from hPSCs and demonstrated that introducing HPS1 mutations produced early-onset fibrotic features [110]. Patient-derived lung spheroids (3D multicellular structures) also recapitulate fibrotic remodeling and show patient-specific responses to antifibrotic drugs, supporting their use as preclinical models [111]. In addition, mutations in HPS2 or HPS4 can induce fibrotic changes in LBOs, while IL-11 knockout attenuates fibrosis in HPS4^−/−^ organoids, highlighting IL-11 as a potential therapeutic target [110,112]. Exosomes released from lung spheroid cells, as well as 3D spheroids cultured from hPSCs, have shown regenerative and anti-fibrotic potential, suggesting novel directions for lung tissue repair [113]. To further explore the molecular drivers of IPF, hiPSC-derived alveolar organoids treated with a fibrosis-inducing cytokine mix (TNFα, PDGF-AB, TGF-β, LPA) exhibited decreased surfactant protein C (SPC) and increased vimentin, KRT17, and KRT8 expression, indicating epithelial injury and maladaptive repair [114]. Pathways such as WNT, BMP, IL-6/STAT3, and FGF7 signaling regulate mesenchymal activity and AT2 cell renewal, while impaired glutamine metabolism reduces alveolar regenerative capacity [115]. These findings emphasize the interplay between epithelial dysfunction and mesenchymal activation in fibrosis. Due to the limitations of alveolar epithelial cell in vitro culture and expansion conditions, specific lung fibrosis models targeting alveolar epithelial cells are rare. Takahiro Suezawa et al. [116] developed fibroblast-dependent alveolar organoids from hPSCs and human fetal lung fibroblasts, enabling long-term expansion and drug screening for IPF. In this model, a TGF-β-driven positive feedback loop in senescent AT2 cells accelerated fibroblast activation, providing a mechanistic basis for disease progression. Takahiro Suezawa et al. developed fibroblast-dependent alveolar organoids from hPSCs and human fetal lung fibroblasts, enabling long-term expansion and drug screening for IPF. In this model, a TGF-β-driven positive feedback loop in senescent AT2 cells accelerated fibroblast activation, providing a mechanistic basis for disease progression. hPSC-derived lung spheroids and organoids more closely resemble natural lung tissue in cell distribution and internal structure. However, they remain fetal-like in maturity, which limits their ability to fully model IPF, which involves age-related epithelial senescence and chronic disease remodeling [117].
Unlike lung fibrosis organoids derived from ESC/iPSC, Surolia et al. directly cultured alveolar organoids from the lung tissue of IPF patients, which replicated in vivo fibrotic characteristics [111]. Similarly, Wilkinson et al. developed a human fetal mesenchymal organoid model that exhibited tissue contraction and strong α-SMA and collagen I expression following TGF-β stimulation, recapitulating early fibrotic remodeling [118]. AT2 cells play a crucial role in alveolar injury repair and fibrosis progression. In human and mouse alveolar organoids, Cebpa downregulation promoted fibrogenic gene expression, whereas Cebpa overexpression restored Sftpc and suppressed fibrosis markers, highlighting its protective role. Consistent with this, activation of the WNT/β-catenin pathway via FZD-specific agonists enhances epithelial renewal and alleviates fibrotic phenotypes. Metabolic regulation also contributes to IPF pathogenesis. Glutamine—a key substrate for energy and biosynthesis—is significantly reduced in AT2 cells from IPF patients and bleomycin-treated mice. Inhibition of glutamine metabolism impairs proliferation and differentiation of AT2-derived alveolar organoids, limiting epithelial regeneration [115]. Additionally, IPF lungs show an increase in abnormal basal-like KRT17^+^ cell populations. Airway basal cells from IPF patients form excessive bronchial spheroids with pro-fibrotic properties, in which SRC signaling is enriched. Pharmacologic SRC inhibition reduces organoid formation and fibrosis progression in vitro and in vivo [119]. Long-term accumulation of basal cells also helps establish a pro-fibrotic niche, as basal cell-derived WNT7A activates fibroblasts and impairs AT2 renewal [120]. These adult stem cell-derived organoid models offer simpler, faster construction and a more faithful representation of human fibrotic tissue. Co-culture systems combining adult epithelial organoids with fibroblasts have become powerful tools for dissecting epithelial–mesenchymal crosstalk in pulmonary fibrosis. Using primary AT2 cells and lung fibroblasts from mice and humans, researchers demonstrated that bleomycin-induced DNA damage triggers p53 activation and a self-sustaining TGF-β feedback loop within AT2 cells, driving inflammation-independent fibrosis. Targeting this autocrine loop may represent a therapeutic strategy for non-inflammatory fibrotic disease [121]. Further supporting this concept, Sui et al. showed that deletion of ANT1 in primary AT2 cells led to cellular senescence and loss of regenerative capacity, resulting in fewer alveolar organoids [122]. Moreover, in AT2–fibroblast co-culture organoids, activation of the TGF-β1/sFRP2 non-canonical WNT signaling axis promoted differentiation into KRT17^+^ basal-like cells, marking the early stages of IPF pathogenesis [123]. Inhibition of ROCK1/2 reversed TGF-β–induced fibroblast differentiation and restored organoid growth, suggesting ROCK signaling as a potential antifibrotic target [124]. Finally, Tan et al. developed airway organoids using adult bronchial epithelial cells, fibroblasts, and microvascular endothelial cells that formed branched tubular structures resembling late fetal lung stages. Although their long-term viability remains limited, these models represent an important step toward building multicellular airway fibrosis organoids [82].
Organoid techniques using patient-derived cells have improved predictions of IPF treatment responses. For instance, iPSCs from IPF patients replicate the fibrotic phenotype, aiding drug efficacy evaluation. iPSC-derived respiratory organoids treated with pirfenidone showed a 35% reduction in collagen (p < 0.01) [125]. In a study of 20 IPF patients, organoids from bronchial biopsies assessed drug response: those with >50% collagen reduction from pirfenidone had an 85% 2-year survival rate, versus 45% for <50% reduction (HR = 0.21, 95% CI: 0.06–0.74, p = 0.016) [111]. A study on a 58-year-old male with IPF unresponsive to standard treatment used ASC-derived organoids to screen 500 compounds, identifying a TGF-β inhibitor that reduced ECM deposition by 60% (p < 0.001) [126]. Off-label treatment improved his FVC by 25% in 3 months and stabilized his condition for a year. These cases demonstrate the translational potential of organoid models in pediatric fibrotic lung diseases.
3.1.2. Cystic Fibrosis
Cystic fibrosis (CF) is a genetic disorder linked to CFTR gene defects [127]. CF organoids have advanced CFTR gene research significantly. Patient-derived PSC organoids can mimic the genetic defect’s impacts on lung development. McCauley et al. [128] created CF-specific airway organoids derived from iPSCs, revealing Wnt-driven airway patterning, and developed a “low Wnt” culture protocol to generate organoids from NKX2-1^+^ progenitor cells. These organoids showed CFTR-dependent swelling upon forskolin stimulation (a forskolin-induced swelling test widely used to assess CFTR-dependent fluid secretion and predict drug responses in CF patients) [129,130]. Sachs et al. [83] developed airway organoids from CF patient ASCs, replicating core CF features, and exhibited significant swelling after CFTR modulation and TMEM16A activation. Previous studies have used the FIS test to assess CFTR modulator responses in distal airway organoids, but results vary due to limited swelling in well-differentiated spherical structures [68,131,132]. Nasal mucosa organoids, however, are more sensitive in detecting CFTR function, making them ideal for personalized CF therapies. In 2020, Liu et al. [132] cultured nasal mucosa organoid models from CF patients and healthy subjects, confirming CFTR gene expression and differences in lumen formation. They also introduced an automated CFTR activity assay using baseline lumen area measurement to evaluate organoid differentiation [67]. Due to the low throughput of functional CFTR assays in previous nasal mucosal organoids studies [131,132,133], there is a need for a scalable FIS detection method using airway organoids from nasal brushing samples. Researchers have improved organoid culture by using 2D differentiated nasal epithelial cell monolayers and optimizing conditions with growth factors like neuregulin-1β and IL-1β to enhance CFTR modulator response detection [134]. Validation showed stable detection of genotype-specific CFTR modulator responses, including the FDA-approved triple regimen (VX-661/VX-445/VX-770) [135]. The short-circuit current (Isc) remains the gold standard for measuring CFTR activity, accurately assessing activity levels and fluid transport effects. Measuring CFTR activity via short-circuit current in vitro is complex. To simplify this, the ANDERSON team [136] developed an automated nasal mucosa organoid swelling assay. This method accurately assesses CFTR activity and is suitable for drug screening. CF patient-derived organoids have been used to establish biobank samples for studies on CFTR repair, including precise gene correction using CRISPR/Cas9 [137]. Nasal mucosa organoids are cost-effective, efficient, and directly relevant to CFTR function, making them valuable for basic CF research, drug development, high-throughput screening, and personalized medicine.
3.2. Chronic Obstructive Pulmonary Disease
Chronic obstructive pulmonary disease (COPD) is a chronic airway inflammatory disease characterized by persistent respiratory symptoms and irreversible airflow obstruction. Significant risk factors for the development of COPD include exposure to harmful particles and gases, notably cigarette smoke and air pollutants. The development of COPD organoid models is commonly facilitated through exposure to cigarette smoke extract (CSE) [138] or particulate matter 2.5 (PM2.5) [139]. Furthermore, organoids derived from the nasal and bronchial epithelial cells of COPD patients demonstrate disease characteristics akin to those observed in COPD, including goblet cell hyperplasia, elevated MUC5AC expression, and a reduction in ciliated cells. The utilization of organoid technology has significantly advanced our understanding of the mechanisms governing the self-renewal, survival, and differentiation of epithelial cells in COPD.
Chan et al. [140] successfully established a COPD organoid model derived from lung adult stem cells of COPD patients. Through single-cell transcriptomic analysis, it was observed that COPD organoids, in comparison to healthy controls, exhibited a reduced presence of basal cells and ciliated cells, alongside an increase in goblet cells. This finding indicates that the model was highly consistent with existing in vitro and in vivo COPD models in terms of phenotype, transcriptome, and functionality. Furthermore, their research showed that COPD organoids exhibited enhanced infection efficiency with SARS-CoV-2, rendering them an ideal model for studying host–pathogen interactions. Utilizing organoid models derived from COPD patients, researchers identified an important intermediate cell population involved in the abnormal developmental trajectory of basal cells in COPD patients. Transcriptomic analysis further suggested that critical signaling pathways, including those related to mitochondrial dysfunction, endoplasmic reticulum stress, ciliary remodeling, extracellular matrix remodeling, and pro-inflammatory cytokine signaling, are disrupted in COPD pathogenesis. During the processes of basal cell regeneration and differentiation, cellular metabolism undergoes dynamic regulation. The researchers observed that basal cells preferentially metabolize lactate through glycolysis during early regeneration [141]. In contrast, during lumen formation, there is a metabolic shift towards the tricarboxylic acid (TCA) cycle [141]. Furthermore, they demonstrated that dysfunction in mitochondrial pyruvate metabolism is a crucial mechanism contributing to basal cell hyperplasia, impaired differentiation of ciliated cells, and the accumulation of intermediate cells. Mesenchymal stem cells, known for their multipotency, secrete regenerative growth factors and immunomodulatory/anti-inflammatory factors, including microRNAs [142,143,144,145]. These properties have led to their widespread application in the treatment of various diseases. In animal models of emphysema, Mesenchymal stem cells have been shown to effectively enhance lung function and improve lung tissue structure, while facilitating the repair of distal alveolar damage. Organoids formed by co-culturing alveolar epithelial cells from non-COPD and COPD patients with lung mesenchymal stem cells suggest that mesenchymal stem cells derived from COPD patients can affect the formation capacity of organoids. Co-culture results showed the formation of larger diameter alveolar organoids with lower early-stage AT2 marker expression, indicating impaired self-renewal and differentiation of alveolar progenitor cells [146]. These findings suggest that abnormal mesenchymal stem cell function may be one reason for the failure of lung tissue repair in COPD.
Key molecular signaling pathways, including Notch, TGF-β, and WNT signaling, play a crucial role in the differentiation of AT2 cells into AT1 cells and in the repair of alveolar epithelium. Disruption of the WNT/β-catenin pathway can lead to dysregulation of distal alveolar epithelial progenitor cell homeostasis and the development of emphysema. Smoking has been shown to downregulate the expression of the WNT receptor protein FZD in both human and murine alveolar epithelial cells, thereby inhibiting WNT/β-catenin signaling activation and impairing the regenerative repair function of COPD organoids [147]. In studies involving murine respiratory organoids exposed to particulate matter (PM2.5), researchers have demonstrated that impaired AT2 to AT1 cell transition may be a mechanism by which PM2.5 contributes to COPD pathogenesis [139]. Given the structural differences between human and mouse airways, in vivo animal models are limited in their ability to fully replicate human airway architecture. Consequently, researchers have employed human respiratory organoid models to identify a unique secretory cell lineage in human distal lung tissue, referred to as RAS cells. RAS cells have the capacity to differentiate into human AT2 cells via Notch and WNT signaling pathways, thereby contributing to alveolar regeneration. However, chronic smoke exposure induces abnormal differentiation of RAS cells into AT2 cells, with this abnormal RAS-AT2 differentiation trajectory potentially playing a crucial role in the pathogenesis of COPD [148].
Furthermore, organoid technology also holds significant advantages in drug screening applications. In a study by Wu et al. [138] transcriptomic data from COPD patients and COPD mouse models exposed to CSE were utilized to identify and investigate potential druggable gene targets for COPD, including the E-series prostaglandin receptor (EP) and prostacyclin receptor (IP). In their study, the researchers developed mouse and human airway and alveolar organoids by co-culturing mouse CD31^−^/CD45^−^/Epcam^+^ lung epithelial cells with CCL-206 mouse lung fibroblasts, and human CD31^−^/CD45^−^/Epcam^+^ lung epithelial cells with MRC5 human lung fibroblasts. These organoids were subsequently exposed to different concentrations of CSE. The findings revealed that alveolar organoids exhibited significantly greater sensitivity to cigarette smoke compared to airway organoids, with CSE exposure markedly inhibiting the formation of alveolar organoids. Moreover, the organoids were utilized for potential drug target screening, demonstrating that EP4 and IP agonists exerted beneficial therapeutic effects on distal alveolar organoids compromised by CSE exposure. Additionally, Rita Costa et al. screened effective WNT/β-catenin activators such as amlexano using mouse COPD organoids and in vivo animal models, proving that amlexano could be a novel drug for improving COPD emphysema [149].
3.3. Respiratory Cancers
3.3.1. Lung Cancer
Research suggests that cancer may originate from endogenous stem cells, within which a subset of stem cells, termed “cancer stem cells” (CSCs), is believed to regulate tumor growth and tumorigenesis [150]. Lung cancer is the most common type of cancer and the leading cause of cancer-related death among men. This type of cancer encompasses a variety of subtypes, each characterized by highly diverse mutation patterns, which complicates both research and treatment efforts. According to the CSCs theory, lung cancer is conceptualized as a stem cell disease resulting from the malignant transformation of adult lung stem cells [151]. Currently, the in vitro disease models employed in cancer research predominantly include 2D cell culture cancer cell lines, patient-derived tumor xenografts (PDXs), and tumor organoids (patient-derived organoids, PDOs). Due to their ease of manipulation and cost-effectiveness, the growth of cancer cell lines in 2D cultures limits their ability to accurately represent the heterogeneity inherent in lung cancer. PDXs are developed by implanting patient tumors into immunodeficient mice, thereby providing a substantial degree of structural fidelity to the original cancer. Due to high costs, lengthy modeling periods, and low success rates, PDXs have limitations. In contrast, PDOs, cultured from patient cancer tissues, offer advantages like higher success rates, shorter modeling times, and lower costs.
Organoid systems derived from cancer tissues have been applied to the primary lung cancer cell cultures [152]. Zhang et al. successfully established patient-derived non-small cell lung cancer (NSCLC) tumor spheres [153]. These tumor spheres contained cancer cells with a high nuclear-to-cytoplasm ratio, deep-staining nuclei, and irregular nuclear membranes, with adenocarcinoma marker Thyroid Transcription Factor-1 (TTF-1) staining positive, consistent with the patient’s original tumor sample, further confirming their adenocarcinoma origin [153]. Serial passage experiments showed that the cytological characteristics of these tumor spheres remained stable, and clinical markers were consistently retained throughout the passage process [153]. Research conducted by Weeber et al. [154] and Sato et al. [18] has corroborated that tumor organoids, aside from their reduced construction times, can maintain genetic and morphological stability even after long-term expansion. These findings provide reliable evidence for the application of tumor organoids. Furthermore, by manipulating organoids derived from normal tissues, it is possible to simulate lung cancer. For instance, Park et al. generated organoids from human bronchial epithelial cells and employed gene editing techniques to upregulate the medulloblastoma and B-cell lymphoma genes [155]. These organoids were subsequently transplanted into immunosuppressed mice, resulting in tumors that closely resembled small cell lung cancer (SCLC). Additionally, Kim et al. [156] developed a basic culture medium devoid of Wnt3a and Noggin to inhibit normal cell growth, thereby creating a library of 80 lung cancer organoid samples encompassing five subtypes of lung cancer: adenocarcinoma, squamous cell carcinoma, small cell carcinoma, adenosquamous carcinoma, and large cell carcinoma. These lung cancer organoids (LCOs) preserved the genetic traits and tissue structure of the original cancer, expressing specific tumor markers and maintaining stable morphology in vitro for up to six months. Using patient samples, primarily epithelial cells, the success rates for culturing were highest for 2D primary lung cancer cells (100%), followed by LCOs (87%), and PDXs (3%). LCOs required only 4 weeks to cultivate, compared to 3–6 months for PDXs, and closely resembled patient tissue morphology. These data demonstrate that LCOs have outstanding advantages in terms of modeling success rate, time, and cost. Additionally, LCOs successfully generated PDXs when transplanted into immunodeficient mice. Loss of liver kinase B1 (LKB1) function is common in adenocarcinoma. Zhang et al. showed that deleting LKB1 in KRAS-positive organoids can convert KRAS-positive adenocarcinomas into squamous cell carcinoma (SCC), a process confirmed in both spontaneous and transplant models [157,158]. These findings suggest new lung cancer treatment strategies, with targeted therapies for LKB1 loss showing promise in preclinical studies. Additionally, organoids from Bcl11a knockout mice confirmed BCL11A as an oncogene in SCC [159].
Beyond their advantages in lung cancer modeling and mechanistic research, respiratory organoids also hold substantial promise in the realm of lung cancer drug discovery and screening. Traditional 2D cell culture systems have often led to the identification of anticancer drugs that exhibit limited efficacy in clinical trials [160]. In contrast, patient-derived organoids, which retain the specific functions of the primary tumor, offer a robust platform for high-throughput drug screening and serve as an important tool for assessing the efficacy of personalized treatments [83]. For example, Barrera-Rodríguez et al. developed a multicellular tumor spheroid model to investigate alterations in drug sensitivity and the molecular mechanisms of multicellular drug resistance [161]. Hai et al. [162] used CRISPR/Cas9 technology to perform gene editing on mouse-derived organoids, successfully knocking out several tumor suppressor genes. By comparing these organoids with patient-derived cell lines, researchers found them to closely resemble human lung squamous cell carcinoma genetically and phenotypically. They proposed a combination therapy hypothesis using immune checkpoint blockade and DNA damage-inducing treatments. Organoids serve as an ideal model for assessing patient-specific drug responses, reacting to drugs based on genomic changes, and providing insights into drug resistance mechanisms. For example, lung cancer organoids with the BRCA2 p.W2619C mutation were more sensitive to olaparib than those with the BRCA2 p.M965I mutation, with olaparib disrupting the structure of the former and inhibiting PDX tumor growth [156]. Wang et al. successfully constructed a patient-derived organoid model from an advanced adenocarcinoma harboring the HER2-A775_G776YVMA insertion mutation [163]. Utilizing this organoid model, the researchers evaluated the efficacy of HER2-targeted therapeutics (afatinib, T-DM1, and pyrotinib). Their findings indicated that pyrotinib exhibited the most significant tumor suppression effects at clinically relevant concentrations both in vivo and in vitro. Furthermore, the researchers assessed the effectiveness of neratinib, afatinib, and gefitinib, proposing that the combination of neratinib and trastuzumab should be considered for clinical trials targeting HER2-mutant lung cancer. Additionally, by combining patient-derived organoids with high-throughput screening systems, the researchers examined the efficacy of EGFR and HER2 inhibitors [164]. In summary, these organoid-based drug screening systems provide important evidence for personalized cancer treatment strategies.
Patient-derived lung cancer organoids exhibit a high fidelity in replicating the tumor’s immune microenvironment. Through the co-culture of lung cancer organoids, autologous peripheral blood lymphocytes in a medium supplemented with interferon-γ, IL-2, and anti-PD-1 antibodies, there is a significant promotion of the expansion and enrichment of tumor-reactive CD4^+^T cells and CD8^+^T cells [165,166]. This methodology maintains the integrity of the tumor’s T cell receptor repertoire and effectively emulates the mechanism of action of immune checkpoint inhibitors [167], thereby facilitating the advancement and implementation of T cell therapies in the treatment of lung cancer.
3.3.2. Nasopharyngeal Carcinoma
Nasopharyngeal carcinoma (NPC) starts in nasopharyngeal epithelial cells. Common NPC cell lines often suffer from HeLa cell contamination, lack of molecular diversity, and loss of the Epstein–Barr virus (EBV) genome over time. Respiratory organoids are increasingly being recognized as a promising model for NPC research. The research team led by Li Gang successfully developed the inaugural patient-derived nasopharyngeal carcinoma organoid (NPCO) model and biobank [73]. Compared to NPCOs, organoids derived from normal nasal mucosa exhibit three morphological types: regular vesicles, irregular vesicles, and irregular liquid shapes, whereas NPCOs consistently manifest a regular spherical form. The study further revealed that recurrent NPCOs exhibit a higher proportion of Bmi-1-positive cells compared to primary NPCOs, suggesting that Bmi-1 expression may play a role in NPCO generation. These nasopharyngeal cancer organoids effectively preserve the pathological and physiological characteristics and genetic heterogeneity of the original tumor. Consequently, they hold significant potential for advancing the development of novel anticancer drugs, investigating the mechanisms underlying NPC development, recurrence, metastasis, and resistance, and formulating personalized treatment strategies.
4. Conclusions and Future Direction
Organoids, resembling real tissues and organs, are ideal in vitro models for research. This review outlines methods for creating respiratory organoid models and recent progress in lung disease research. Their use has been crucial for understanding lung epithelial stem/progenitor cell changes in disease, advancing personalized medicine, and exploring lung regeneration. However, current organoid systems still have limitations in replicating in vivo cell maturity, interactions, and complexity.
Currently, organoid cultivation primarily relies on mouse-derived extracellular matrix substitutes like Matrigel, which can vary in terms of reproducibility during research. These substitutes may also contain unknown pathogens, limiting their direct application in clinical transplantation. Clinical-grade collagen could address these issues [168], and bioengineering strategies may further optimize cultivation techniques [169].
A key challenge for using organoid culture methods in regenerative medicine is their inability to mimic multi-organ pathology due to missing cell lineages from different organs. Advanced cellular engineering, such as co-culture of respiratory organoids with niche cells or other organoids, is crucial to address this. Progress is being made with multi-lineage organoids; for example, merging organoids from various brain regions of hPSCs has allowed researchers to replicate neuronal migration, projection, and neurodevelopmental disorders, enhancing the study of multicellular interactions [170,171]. These “assemblies” are multi-regional organoids combining various cell lineages to model complex liver and gallbladder diseases [172,173]. Advances in sequencing have allowed the integration of previously unknown lung cell types, such as CFTR-rich ionocytes, into organoids, crucial for understanding cystic fibrosis [174]. Using ALI technology, researchers co-cultured neuroendocrine cells with fetal lung fibroblasts to create respiratory organoids with rare neuroendocrine cells [175], closely resembling natural lung epithelial cells. Advanced techniques like single-cell RNA sequencing and spatial transcriptomics validate these organoids, enhancing disease modeling and paving the way for future multi-organ organoids and regenerative medicine applications. Current tumor organoids lack essential elements like biomechanical stimulation, matrix components, blood vessels, and immune cells [176], limiting their use in cancer research. Integrating these could enhance organoid systems, offering better solutions for cancer research and treatment. Recent strategies involve organ-on-a-chip technology to simulate multi-organ interactions and 3D bioprinting with hydrogel to create complex respiratory organoids [177,178]. These innovations pave the way for precise cancer research and personalized treatment.
Tissue maturity is a third key limitation of organoid technology in research and clinical applications. PSC-derived organoids often lack cellular maturity, hindering accurate simulation of human organs. Their fetal-like characteristics can lead to discrepancies in replicating adult disease pathophysiology, as they may not fully capture the complex tissue structure and function of adult lungs. In conditions like IPF and COPD, age-related changes such as fibrosis and cellular senescence are crucial but less evident in immature organoids from ESCs or iPSCs. Thus, while useful for studying early disease mechanisms, these models require cautious interpretation for adult-onset diseases. To address this, researchers use strategies like pre-treating organoids with small molecules such as BDNF (brain-derived neurotrophic factor) to speed up maturation [179]. Additionally, enhancing oxygenation and nutrient diffusion, particularly through vascularization, improves tissue maturity and extends organoid survival [3]. A study demonstrated the coordinated development of human heart-lung development by inducing multi-lineage differentiation of hiPSCs, highlighting the cardiac lineage’s role in alveolar maturation [180]. Wimmer et al. developed a method to transform hiPSCs into mesodermal cells and then into vascular organoids using growth factors like VEGF-A and FGF2 [23]. When transplantation into murine models, these organoids successfully developed a complete vascular system, thereby providing a valuable model for investigating diabetic vascular complications and facilitating the development of therapeutic interventions. Moreover, researchers have vascularized human respiratory organoids derived from hiPSC lung progenitor cells and endothelial cells, demonstrating differentiation into AT2 and AT1 cells, as well as eliciting inflammatory responses [181]. In a pioneering study, scientists have, for the first time, developed highly vascularized lung and intestinal organoids from human iPSCs [181]. This was accomplished by engineering a 3D system that mimics human embryonic development, allowing for the co-differentiation of mesoderm and endoderm to form blood vessels and lung/gut epithelium within a single spheroid. This achievement overcomes the limitations of traditional organoid models, which typically lack functional vasculature and organ-specific stroma, thereby providing a novel platform for the study of organ development, disease pathogenesis, and regenerative medicine. Our recent study reveals that BMPR2 mutation impairs late-stage lung progenitor differentiation and airway epithelial maturation. BMPR2-mutant airway organoids exhibit impaired goblet and ciliated cell development, with increased basal/club cell markers, suggesting arrested differentiation [182]. These insights highlight BMPR2’s role in lung development and its potential link to pulmonary hypertension, offering valuable perspectives for understanding lung development, inflammatory lung injuries, and advancing organoid technology.
In addition, limited reproducibility is a major challenge in organoid research, hindering the creation and control of advanced organoids. Key factors affecting the reproducibility of organoids include batch differences, production scalability, cell composition stability, and structural complexity [3]. Developing automated culture and analysis platforms is crucial to address these issues, enabling standardized large-scale organoid production and high-throughput drug screening [183]. While mouse respiratory organoids show high complexity, identifying equivalent human cells could lead to more complex, personalized organoids from patient samples. This approach will enhance model accuracy for studying human lung diseases. Future use of integrated organoid chip-based culture platforms is anticipated to improve precise modeling of chronic lung diseases. Coupled with high-resolution analytical technologies, these platforms will expand opportunities to explore complex pulmonary pathophysiological mechanisms.
Respiratory organoid technology is widely used in biomedical research, drug screening, and gene therapy. Although its application is still being explored, resolving current challenges will improve its relevance to the human respiratory system. The use of AI and machine learning can enhance image analysis and predict drug responses. Integrating single-cell RNA sequencing and spatial transcriptomics offers perspectives for achieving high-resolution characterization of organoid heterogeneity, lineage trajectories, and disease-associated cell states. Combining organoids with microfluidic platforms and organ-on-chip systems can better simulate physiological conditions. These advancements will deepen our understanding of chronic lung disease and improve patient outcomes.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Clevers H. Modeling Development and Disease with Organoids Cell 20161651586159710.1016/j.cell.2016.05.08227315476 · doi ↗ · pubmed ↗
- 2Zhao Z. Chen X. Dowbaj A.M. Sljukic A. Bratlie K. Lin L. Fong E.L.S. Balachander G.M. Chen Z. Soragni A. Organoids. Nature reviews Methods Primers 202229410.1038/s 43586-022-00174-y 37325195 PMC 10270325 · doi ↗ · pubmed ↗
- 3Tang X.-Y. Wu S. Wang D. Chu C. Hong Y. Tao M. Hu H. Xu M. Guo X. Liu Y. Human organoids in basic research and clinical applications Signal Transduct. Target. Ther.2022716810.1038/s 41392-022-01024-935610212 PMC 9127490 · doi ↗ · pubmed ↗
- 4Saraswathibhatla A. Indana D. Chaudhuri O. Cell-extracellular matrix mechanotransduction in 3D Nat. Rev. Mol. Cell Biol.20232449551610.1038/s 41580-023-00583-136849594 PMC 10656994 · doi ↗ · pubmed ↗
- 5Baker B.M. Chen C.S. Deconstructing the third dimension: How 3D culture microenvironments alter cellular cues J. Cell Sci.20121253015302410.1242/jcs.07950922797912 PMC 3434846 · doi ↗ · pubmed ↗
- 6Scalise M. Marino F. Salerno L. Cianflone E. Molinaro C. Salerno N. De Angelis A. Viglietto G. Urbanek K. Torella D. From Spheroids to Organoids: The Next Generation of Model Systems of Human Cardiac Regeneration in a Dish Int. J. Mol. Sci.2021221318010.3390/ijms 22241318034947977 PMC 8708686 · doi ↗ · pubmed ↗
- 7Bresnahan E. Ramadori P. Heikenwalder M. Zender L. Lujambio A. Novel patient-derived preclinical models of liver cancer J. Hepatol.20207223924910.1016/j.jhep.2019.09.02831954489 · doi ↗ · pubmed ↗
- 8CorròC. Novellasdemunt L. Li V.S.W. A brief history of organoids Am. J. Physiol. Cell Physiol.2020319 C 151C 16510.1152/ajpcell.00120.202032459504 PMC 7468890 · doi ↗ · pubmed ↗