Paenibacillus encodes a membrane-localized Spo0B
Isabella N. Lin, Cassidy R. Prince, Heather A. Feaga

TL;DR
This study identifies a membrane-localized variant of the Spo0B protein in Paenibacillus, which is important for sporulation and is widely conserved across the genus.
Contribution
The discovery of a conserved membrane-localized Spo0B variant in Paenibacillus, which is crucial for sporulation phosphorelay interactions.
Findings
Spo0B in Paenibacillus contains a transmembrane domain and is localized to the cell membrane.
Spo0B-TM is present in 92% of surveyed Paenibacillus genomes and is important for phosphorelay interactions.
Spo0B shows low sequence identity across Bacillota compared to other phosphorelay members.
Abstract
Sporulation is a strategy employed by many bacteria to survive harsh environmental conditions. The genus Paenibacillus includes spore-forming species notorious for spoiling pasteurized dairy products and for causing American foulbrood in honeybee larvae, leading to colony collapse. Human pathogens within Paenibacillus are also a growing threat, causing fatal opportunistic infections. Here, we present a comprehensive survey of sporulation genes across 1,460 high-quality Paenibacillus genomes. We find that all members of the sporulation-initiating phosphorelay are well conserved, but that the Spo0B phosphotransferase contains a predicted transmembrane domain. We confirm that this domain localizes Spo0B to the cell membrane and therefore refer to this Spo0B variant as Spo0B-TM. Spo0B-TM is present in 92% of surveyed Paenibacillus genomes. Consistent with its high level of conservation, we…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
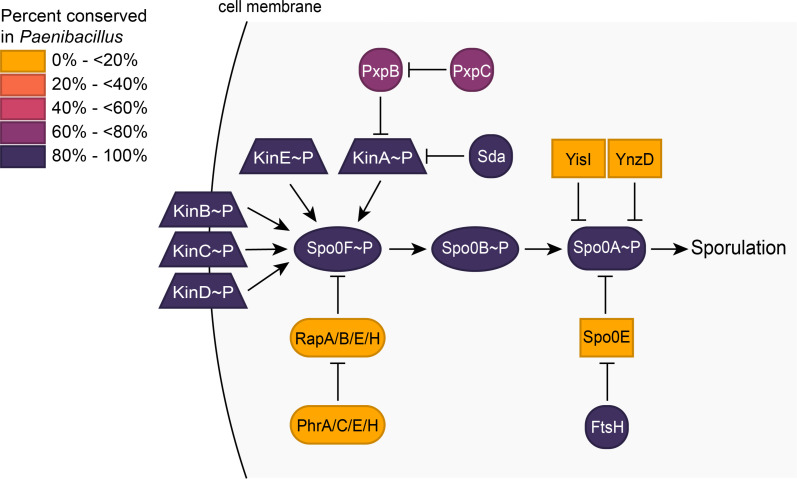 Fig 1
Fig 1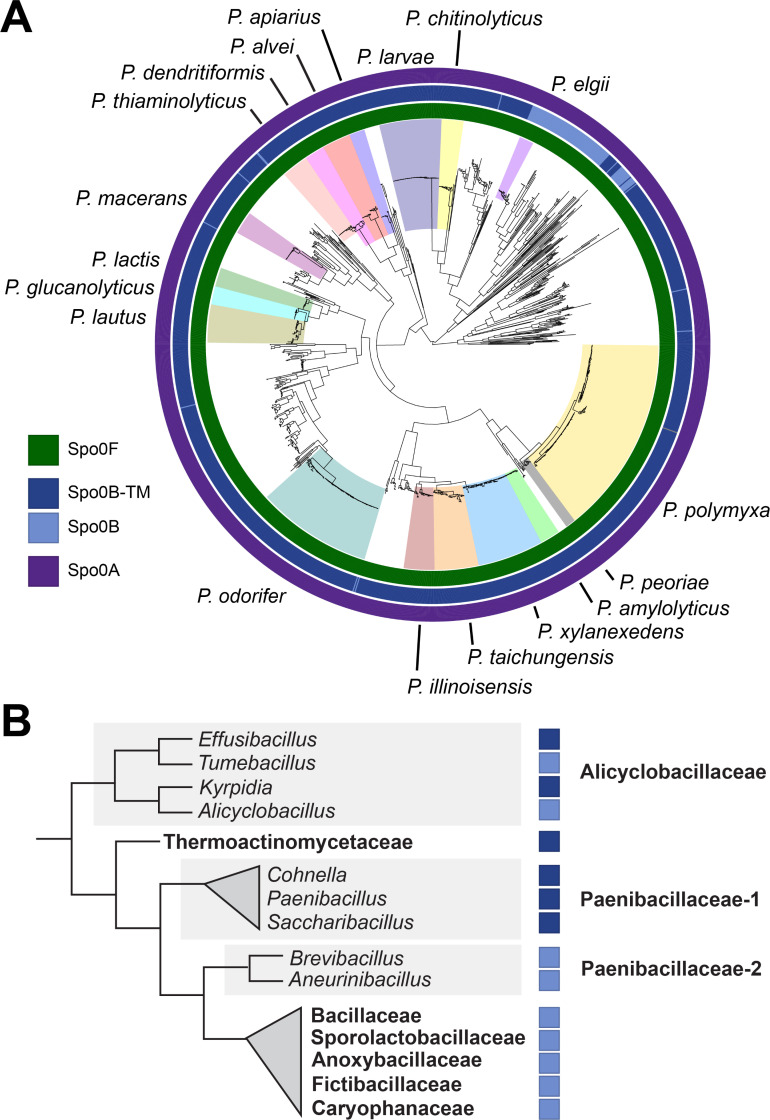 Fig 2
Fig 2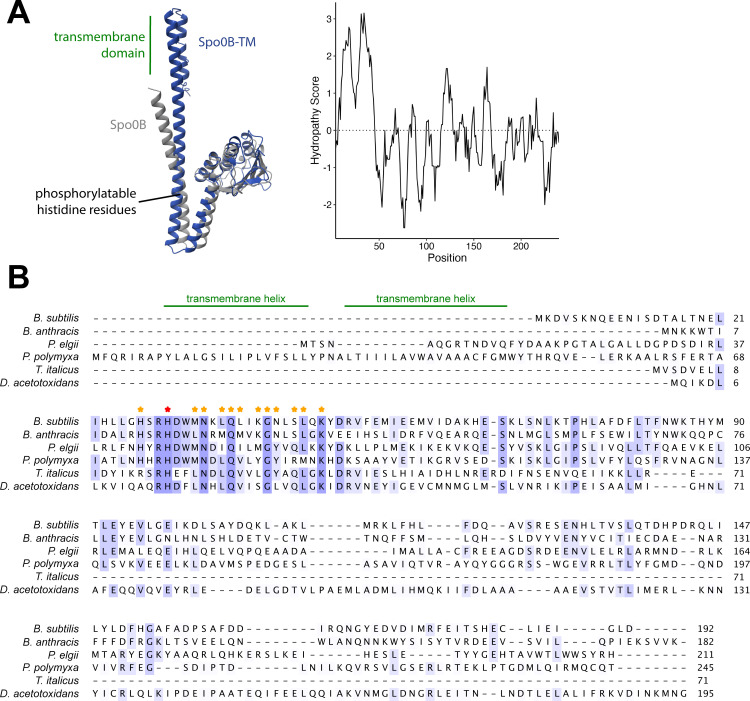 Fig 3
Fig 3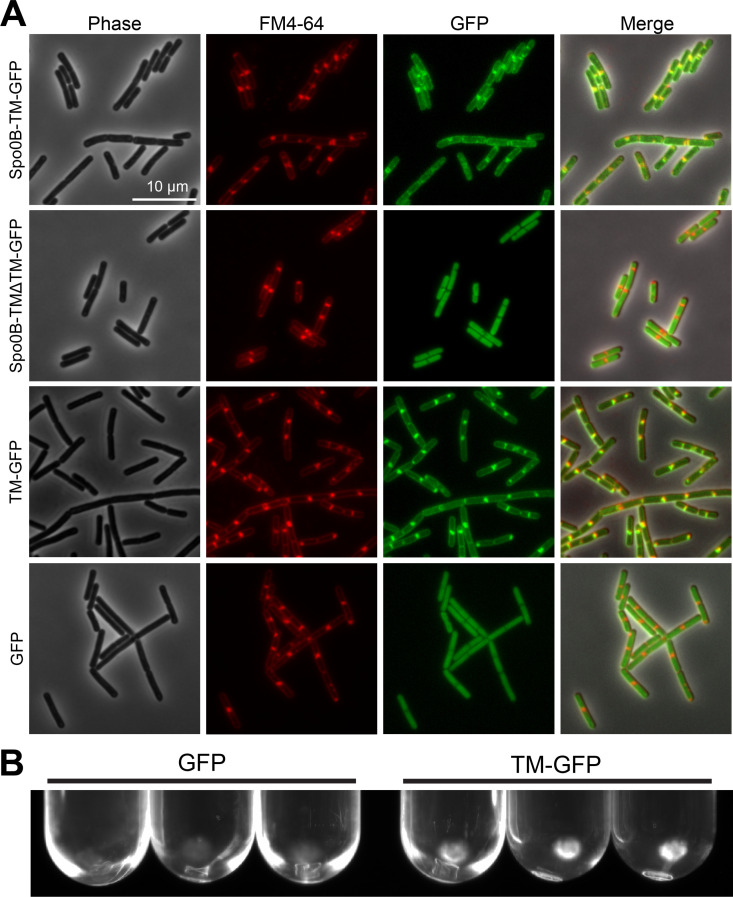 Fig 4
Fig 4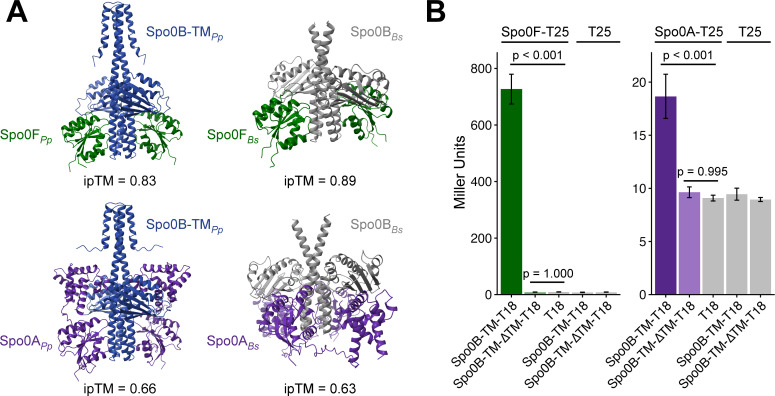 Fig 5
Fig 5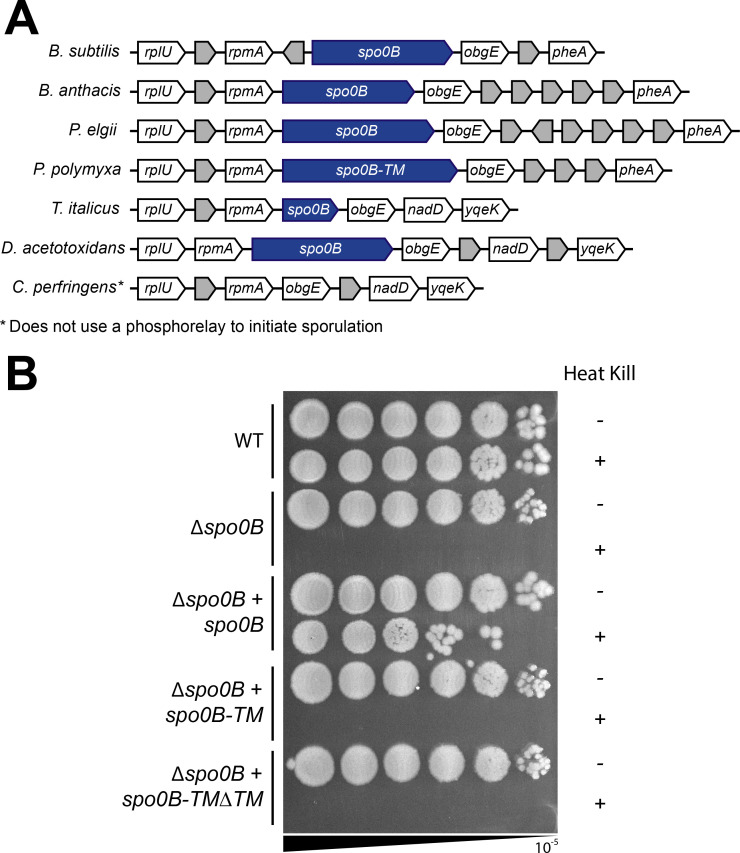 Fig 6
Fig 6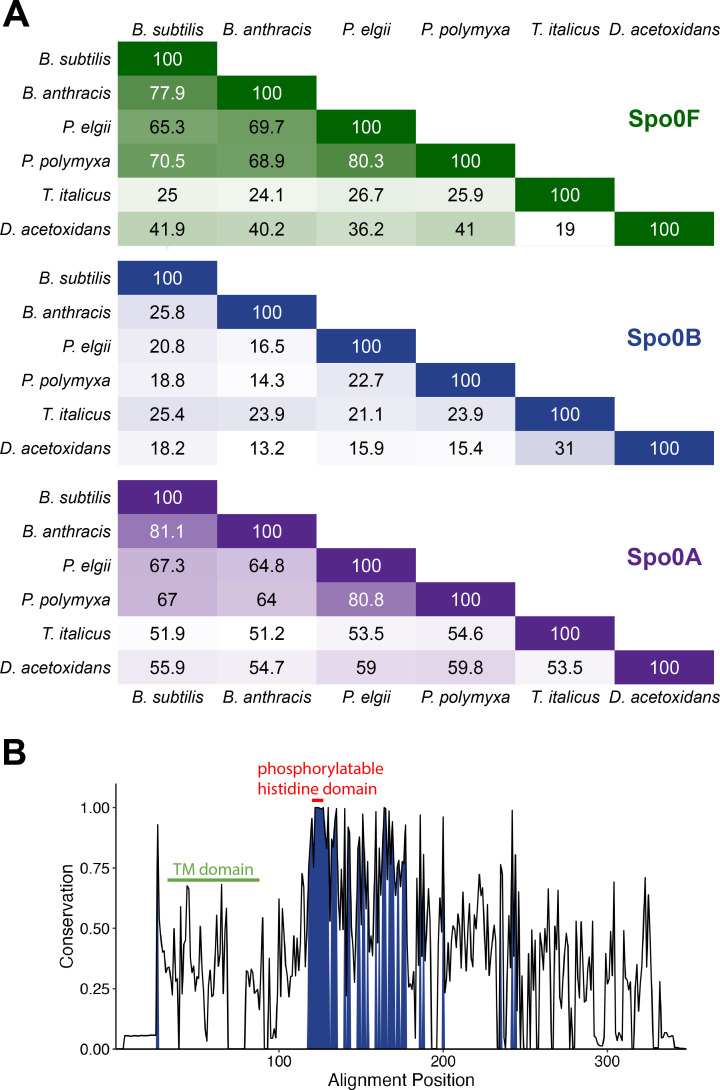 Fig 7
Fig 7| Description or sequence | Source or reference | |
|---|---|---|
|
| ||
| BTH101 | ( | |
| HAF1 | ( | |
| IM39 | ( | |
| CP347 | 168 | This study |
| CP348 | 168 | This study |
| IM37 | 168 | This study |
| IM71 | 168 | This study |
| IM72 | 168 | This study |
| IM73 | 168 | This study |
|
| ||
| pHF549 | pDR111 with P | This study |
| pCP336 | pHF549 | This study |
| pCP338 | pHF549 | This study |
| pKNT25 | BACTH plasmid | ( |
| pUT18 | BACTH plasmid | ( |
| pIM95 | pKNT25 carrying | This study |
| pIM79 | pUT18 carrying | This study |
| pIM75 | pUT18 carrying | This study |
| pIM92 | pKNT25 carrying | This study |
|
| ||
| IM11-F | 5′- | |
| IM12-R | 5′- | |
| IM13-F | 5′- | |
| IM14-F | 5′- | |
| IM15-R | 5′- | |
| IM18-F | 5′- | |
| IM19-R | 5′- | |
- —National Institute of General Medical Scienceshttp://dx.doi.org/10.13039/100000057
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBacterial Genetics and Biotechnology · Bacillus and Francisella bacterial research · Bacterial biofilms and quorum sensing
INTRODUCTION
Endospore formation is a complex developmental process limited to certain classes of Bacillota (1, 2). Through sporulation, vegetative bacteria undergo developmental and metabolic changes to produce highly resistant, dormant spores that are capable of withstanding adverse environmental conditions (3). Spore formation involves a series of morphological stages that are typically triggered by low nutrient density. A vegetative cell asymmetrically divides into a mother cell and a forespore, after which the forespore is fully engulfed and released into the mother cell cytoplasm (4–7). The forespore is encased in two protective shells, the cortex and coat (8–10), and the mature spore is released via lysis of the mother cell (9). Upon sensing germinants, typically nutrients, the spore will germinate and outgrow into a vegetative cell (11, 12).
Endospore formation is well characterized in Bacillus subtilis, where it affects the expression of over 500 genes (13, 14). Sporulation-associated genes continue to be identified and characterized (15–20). A transposon screen by Meeske et al. (21) identified 157 genes required for the sporulation of B. subtilis, including 24 previously unidentified genes. Genes required for sporulation include kinases and phosphotransferases that initiate sporulation and sigma factors that control endospore progression, chromosome segregation, and production of the spore coat. Many of these proteins have dedicated roles in sporulation, while others, such as Spo0A, also play a role in vegetative growth (22–24).
B. subtilis, along with many other Bacilli and Clostridia, initiates sporulation via a phosphorelay system (25–27). Five autophosphorylating sensor histidine kinases (KinA-KinE) respond to high cell density and low nutrient availability by transferring a phosphoryl group to Spo0A via the phosphotransferases Spo0F and Spo0B (4, 27–34). Spo0A, the key transcriptional regulator of sporulation, is activated in its phosphorylated form and directly influences the expression of over 100 genes to govern entry into sporulation (35, 36). In Bacilli and Clostridia, sporulation kinases are highly conserved, as is Spo0A, which is found even in non-sporulating members of Bacillota (1). In contrast, many species within Clostridia lack the phosphotransferases, and instead, multiple kinases phosphorylate Spo0A directly (25, 37–39).
Paenibacillus is a spore-forming genus of bacteria encompassing over 200 species with diverse characteristics (40). Paenibacillus is commonly found in soil, and many species, such as P. polymyxa, P. macerans, and P. elgii, promote the growth of plants through phosphate solubilization or nitrogen fixation (41–43). Species including P. alvei, P. ehimensis, and P. kribbensis contribute to biological control by producing biocidal compounds (44–46). While previously considered non-pathogenic, recent case reports document Paenibacillus spp*.,* including P. alvei and P. thiaminolyticus, to cause sepsis in infants (47–50) and severe infections in adults, typically as an opportunistic pathogen (48, 51–53). Paenibacillus is also the etiological agent of American foulbrood, a devastating honeybee brood disease caused by P. larvae, for which there is currently no treatment and necessitates burning of infected hives (54, 55). Species such as P. odorifer, P. amylolyticus, and P. lactis are the causative spoilage agents of a variety of food products, including pasteurized and chilled items such as dairy and ready-to-eat meals (56–58). Studying the sporulation of diverse Paenibacillus impacts many industries.
Here, we survey 1,460 high-quality Paenibacillus genomes for known sporulation-associated protein-coding genes from B. subtilis. We detected 632 B. subtilis sporulation genes in Paenibacillus, with 350 of these genes conserved in ≥95% of the genomes. While much of the sporulation pathway is conserved between Paenibacillus and B. subtilis, we determined that most Paenibacillus encode a unique Spo0B variant containing an N-terminal domain that localizes it to the cell membrane (Spo0B-TM). Spo0B-TM from P. polymyxa is not functionally interchangeable with Spo0B in B. subtilis, but Spo0B-TM associates with its phosphorelay partners, Spo0F and Spo0A, and the transmembrane domain is likely important for this association. Across Spo0B orthologs in Bacillota, we found that while the phosphorylatable histidine region is strongly conserved in sequence, the rest of the protein is highly variable between species. In the case of Spo0B from Paenibacillus, this sequence variation is likely consequential.
RESULTS
Survey of the Paenibacillus sporulation pangenome identifies core sporulation genes that are shared with other spore-forming Bacillota
To survey the conservation of known sporulation genes throughout the Paenibacillus genus, we downloaded all high-quality annotated Paenibacillus genome assemblies (n = 1,460) from NCBI and performed a BLASTP search using 741 B. subtilis sporulation protein sequences. Genes encoding the protein sequences used in our search included genes predicted to be essential for sporulation in B. subtilis (21), genes classified by SubtiWiki as sporulation genes (59), and genes in the sporulation gene set defined by Galperin et al. (23). The surveyed Paenibacillus genomes represent over 250 unique species (Table S1), with strong representation from P. polymyxa (n = 117), P. larvae (n = 64), and P. odorifer (n = 37). As additional genomes were included in the search, fewer additional sporulation genes were detected, indicating that the search was performed to saturation. Thus, the number and quality of genomes surveyed were sufficient to capture an accurate, extensive representation of likely sporulation genes within the genus that are shared with B. subtilis.
Of the 741 B. subtilis sporulation genes included in our search, 632 were detected in at least one of the Paenibacillus genomes (Table S2), with a mean of 453 sporulation genes detected per genome. To determine which known sporulation genes may be most important for sporulation in Paenibacillus, we split the sporulation pangenome into core, shell, and cloud genomes. Genes found in ≥95% of the surveyed genomes made up the core, genes found in <95% but ≥15% of genomes made up the shell, and the remaining genes made up the cloud. Galperin et al. (23) constructed a list of ~120 universally conserved sporulation genes in Bacillota. We detected all 120 of these genes in Paenibacillus and found 115 of these genes in the core sporulation genome. The five genes missing from the core genome are sweC (detected in 1.4% of genomes), yciB (7.8%), divIC (28%), ylbB (81%), and yloC (89%). Over half of the sporulation genes we identified were core genes (n = 350), of which 14 are essential for life in B. subtilis (60). Meeske et al. (21) identified 142 genes required for sporulation in B. subtilis, excluding genes involved in the TCA cycle. We found 127 of these genes in the sporulation core for Paenibacillus, and the remaining 15 genes are listed in Table S3. The key players in the phosphorelay, KinA-E, Spo0F, Spo0B, and Spo0A, are conserved in >99% of surveyed Paenibacillus genomes (Fig. 1 and 2A; Table S2). The sporulation-specific sigma factors, SigE, SigF, SigK, and SigG (61), are conserved in 100% of the genomes (Table S2). These results indicate that Paenibacillus shares a very similar sporulation pathway with B. subtilis.
Key sporulation initiation proteins in B. subtilis are conserved in Paenibacillus. Heat map of the sporulation initiation phosphorelay of B. subtilis, showing the conservation of the proteins across Paenibacillus. Trapezoids denote kinases; ovals, phosphotransferases; rectangles, phosphatases. Genomes surveyed are listed in Table S1 and a full list of sporulation genes and their conservation can be found in Table S2.
Spo0B has a unique transmembrane domain in most Paenibacillus genomes. (A) A midpoint-rooted maximum likelihood phylogenetic tree of Paenibacillus genomes built with single-copy core genes. Species with at least nine available high-quality genomes are labeled. Color coding indicates the presence of Spo0F, Spo0B, and Spo0A, and the transmembrane domain variant of Spo0B (Spo0B-TM). Paenibacillus genomes used in the analysis are listed in Table S1. (B) A cladogram of sporulating Bacilli with branching based on the Genome Taxonomy Database (GTDB) (62, 63). Taxonomies are derived from NCBI Taxonomy (64, 63). Families with >10 reference genomes and genera with >5 reference genomes are shown. Family names are bolded, and genera are italicized. When >33% of a family or genus encodes Spo0B-TM, this is indicated with a dark blue box in accordance with the legend in 2A.
A Spo0B variant with a predicted transmembrane domain is conserved across Paenibacillus
A previous study of P. polymyxa sporulation histidine kinases examined the Spo0B sequences of six Paenibacillus genomes and found a unique extended N-terminus containing two hydrophobic helices that are predicted to form a transmembrane domain (65). We will refer to Spo0B containing this domain as Spo0B-TM. Of the Paenibacillus genomes surveyed, 92% encode Spo0B-TM (Fig. 2A). The remaining 8% of Paenibacillus genomes encode a Spo0B that lacks the transmembrane domain. Species that do not contain the transmembrane domain in Spo0B clade together and include P. elgii, P. validus, P. mucilaginosus, P. ehimensis, and P. tyrfis (Fig. 2A).
Due to the strong conservation of Spo0B-TM within Paenibacillus, we expanded our search to include all RefSeq reference genomes in the Bacilli class (n = 2,425, including 316 Paenibacillus spp.). Spo0B was detected in 1,385 genomes, and a predicted transmembrane domain was identified in 27% of Spo0B proteins. This variant is restricted primarily to the clade of Bacilli that includes the families of Paenibacillaceae (78% of 416 species with Spo0B contain the transmembrane domain), Thermoactinomycetaceae (95% of 42 genomes), and Alicyclobacillaceae (38% of 21 genomes) (Fig. 2B). The transmembrane domain is completely absent in clade Paenibacillaceae-2, which contains the genera Aneurinibacillus and Brevibacillus, and in the clades of Alicyclobacillaceae containing the genera Tumebacillus and Alicyclobacillus (Fig. 2B). RefSeq accessions and assembly names, taxonomic lineage, and Spo0B and transmembrane domain presence for all Bacilli surveyed are listed in Table S4.
Despite low percent identity at the protein sequence level (19%), Spo0B from B. subtilis and Spo0B-TM from P. polymyxa are predicted to have a high degree of structural similarity when modeled using AlphaFold (Fig. 3A). Alignment of six Spo0B orthologs from Bacilli and Clostridia revealed a highly conserved region around the phosphorylatable histidine residue (Fig. 3B). As expected, the residues that directly interact with Spo0F and Spo0A in the α1 helix of B. subtilis Spo0B (66) are completely or highly conserved (Fig. 3B).
Predicted structure and sequence alignment of Spo0B-TM with other Spo0B proteins from Bacillota. (A) AlphaFold model of Spo0B from B. subtilis and Spo0B-TM from P. polymyxa and Kyte-Doolittle hydropathy plot of Spo0B-TM from P. polymyxa. (B) Protein sequence alignment of Spo0B in B. subtilis and its orthologs across Bacilli (Bacillus anthracis, Paenibacillus elgii, Paenibacillus polymyxa) and Clostridia (Thermoanaerobacter italicus, Desulfofarcimen acetotoxidans). Amino acids are shaded based on conservation, with completely conserved residues shown in dark purple. The transmembrane helices of Spo0B-TM from P. polymyxa are annotated. Residues in the α1 helix of Spo0B from B. subtilis that are known to interact with Spo0F and Spo0A are indicated with stars. The phosphorylatable histidine residue is denoted by the red star. Spo0B sequence accessions are listed in Table S5.
Spo0B-TM localizes to the cell membrane when expressed in B. subtilis
To determine whether the bioinformatically predicted transmembrane domain of Spo0B-TM imparts membrane localization, we fused GFP to the C-terminus of P. polymyxa Spo0B-TM (Spo0B-TM-GFP) and expressed this protein in B. subtilis. As a control, we fused GFP to the C-terminus of P. polymyxa Spo0B-TM from which we deleted the transmembrane domain (Spo0B-TM∆TM-GFP). We then monitored the cellular localization of these proteins using fluorescence microscopy. Spo0B-TM-GFP localized to the cell membrane, exhibiting a membrane localization pattern that was similar to the membrane stain FM4-64 (Fig. 4A). In contrast, Spo0B-TM∆TM-GFP was located throughout the cytoplasm. To further confirm that the transmembrane domain is sufficient to impart localization, we fused only the transmembrane domain to GFP (referred to as TM-GFP). TM-GFP visibly localizes to the membrane, similar to Spo0B-TM-GFP. In contrast, GFP lacking the transmembrane domain is diffuse throughout the cytoplasm (Fig. 4A).
Spo0B-TM localizes to the cell membrane in B. subtilis. (A) Microscope images of B. subtilis expressing P. polymyxa Spo0B-TM fused to GFP (Spo0B-TM-GFP) or P. polymyxa Spo0B-TM lacking the TM domain fused to GFP (Spo0B-TM∆TM-GFP). TM-GFP indicates the TM domain of Spo0B-TM is fused directly to GFP. Membranes were stained with FM4-64. (B) Pellets containing membrane fractions illuminated by blue light.
To further confirm membrane localization of the transmembrane domain from Spo0B, we used a biochemical approach to fractionate the cell membrane (67). Consistent with the results of the microscopy, pellets resulting from ultracentrifugation of cell lysate collected from B. subtilis cells expressing TM-GFP fluoresced under blue light, whereas pellets from B. subtilis cells expressing GFP without the transmembrane domain did not fluoresce (Fig. 4B). Collectively, our results indicate that Spo0B-TM from P. polymyxa exhibits membrane localization and that the transmembrane domain alone is sufficient for localization.
Spo0B-TM interacts with phosphorelay proteins Spo0A and Spo0F from P. polymyxa
To predict if P. polymyxa Spo0F and Spo0A could interact with Spo0B-TM, we modeled the interactions using AlphaFold 3 (Fig. 5A). The interaction between Spo0F and Spo0B-TM had an ipTM score >0.8, indicating a confident, high-quality prediction (68). The interaction between Spo0A and Spo0B-TM had an ipTM score of 0.66, indicating the prediction was made with lower confidence. However, the interaction model of B. subtilis Spo0A and Spo0B, proteins known to interact, similarly yielded an ipTM score of 0.63. Therefore, the low score of the Spo0A and Spo0B-TM interaction should not discredit the modeled interaction. The location of the binding between the two protein pairs across species also appeared similar (Fig. 5A).
Spo0B-TM interacts with both Spo0F and Spo0A from P. polymyxa. (A) AlphaFold models of P. polymyxa and B. subtilis protein interactions. ipTM scores >0.8 represent high confidence predictions. Pp, P. polymyxa; Bs, B. subtilis. Protein sequence accessions used for modeling are listed in Table S5. (B) Quantification of β-galactosidase activity for BACTH constructs using P. polymyxa proteins, representative of nine biological replicates. Error bars represent SEM. P-values indicate the results of a one-way ANOVA followed by Tukey’s test.
We then examined the in vivo interaction of Spo0B-TM with Spo0A and Spo0F from P. polymyxa using the bacterial adenylate cyclase two-hybrid (BACTH) system in Escherichia coli. The BACTH system utilizes the interaction-mediated reconstitution of adenylate cyclase from Bordetella pertussis (69). The catalytic domain of this protein is active only when its two complementary fragments (T18 and T25) are brought together to allow for functional complementation. When interacting proteins are fused to T18 and T25, heterodimerization of the proteins results in functional complementation of the adenylate cyclase fragments. Adenylate cyclase synthesizes cAMP, which induces the production of β-galactosidase in the cell. Therefore, we used β-galactosidase activity as a reporter to quantify protein-protein interactions.
We fused P. polymyxa Spo0F or Spo0A to the T25 fragment and P. polymyxa Spo0B-TM to the T18 fragment of the adenylate cyclase catalytic domain. Spo0B-TM showed strong interaction with Spo0F, with a β-galactosidase activity of 727 ± 53 Miller units (Fig. 5B). The interaction of Spo0B-TM with Spo0A yielded a more modest level of β-galactosidase activity (19 ± 2 Miller units), but this activity was significantly higher than the negative controls (P < 0.001 compared with all negative controls) (Fig. 5B). Interestingly, Spo0B-TM did not exhibit detectable interaction with either Spo0A or Spo0F when the transmembrane domain was deleted (Spo0B-TM∆TM), with β-galactosidase activity comparable to the negative controls (Fig. 5B). These results suggest that Spo0B-TM interacts with Spo0A and Spo0F of P. polymyxa, and that the transmembrane domain is necessary to detect this interaction. One caveat of the BACTH system is that protein levels of Spo0B-TMΔTM-T18 may be reduced compared with Spo0B-TM-T18. In either case, these data suggest that the TM domain of Spo0B has an important role in the function of this protein.
Spo0B-TM is syntenic, but not functionally interchangeable with Spo0B in B. subtilis
While two-component signaling proteins are typically organized in pairs or in adjacent operons (70), the genes encoding the sporulation initiation phosphorelay are scattered throughout the genome. In B. subtilis, spo0B is located downstream of rplU and rpmE, genes encoding ribosomal proteins bL21 and bL31 (71, 72), and upstream of obg, the gene encoding the GTP-binding protein Obg (73). We examined the syntenic regions in Bacilli and Clostridia species and found that the genomic arrangement of spo0B to be well conserved (Fig. 6A).
Spo0B-TM from P. polymyxa does not rescue the sporulation defect of a spo0B deletion in B. subtilis. (A) Synteny diagram of the spo0B genomic region across Bacillus subtilis, Bacillus anthracis, Paenibacillus elgii (spo0B), Paenibacillus polymyxa (spo0B-TM), Thermoanaerobacter italicus, Desulfofarcimen acetotoxidans, and Clostridium perfringens. Genome accessions are listed in Table S5. (B) Sporulation assay of B. subtilis strains. Serial dilutions of culture were plated before and after heating to 80˚C to kill vegetative cells. Spot titers are representative of three biological replicates.
Since the genomic context and the phosphorylatable histidine domain of Spo0B-TM are conserved and Spo0B-TM can interact with other members of the P. polymyxa phosphorelay, we hypothesized that Spo0B-TM may be functional in related spore-formers. To test if spo0B-TM is functionally interchangeable with spo0B in B. subtilis, we expressed P. polymyxa spo0B-TM under the control of the native spo0B promoter and determined whether it could complement the Δspo0B mutant of B. subtilis. Spo0B is essential for sporulation in B. subtilis, and as expected, the ∆spo0B mutant failed to produce spores (Fig. 6B). Complementation with spo0B expressed under its native promoter restored sporulation efficiency to nearly wild-type levels (Fig. 6B). The incomplete complementation may be because deleting spo0B has a polar effect on the downstream gene obg, a gene that is essential for both growth and sporulation (74, 75). Complementation with spo0B-TM, regardless of the presence or absence of its transmembrane domain, did not rescue the sporulation defect of the ∆spo0B mutant, as no heat-resistant spores were detected in either strain (Fig. 6B). Therefore, spo0B-TM and spo0B-TM∆TM from P. polymyxa are not functionally interchangeable with spo0B in B. subtilis, despite the highly conserved region surrounding the phosphorylatable histidine.
Spo0B exhibits the lowest sequence identity of members of the phosphorelay
Since neither Spo0B-TM nor Spo0B-TM∆TM from Paenibacillus could rescue sporulation in a ∆spo0B mutant of B. subtilis, we next examined the sequence conservation of Spo0B proteins across Bacillota. Despite a high degree of structural similarity predicted by AlphaFold (Fig. 3A), B. subtilis and P. polymyxa Spo0B share only 19% protein sequence identity (Fig. 7A). In contrast, Spo0F and Spo0A from these species share 70% and 67% protein sequence identity, respectively. Consistent with the low sequence identity of Spo0B, a search using BLASTN with B. subtilis spo0B as the query identified no significant Spo0B homologs in Paenibacillus. However, a search with BLASTP yielded hits for Spo0B in 1,450/1,460 genomes. Using HMMER, we were able to identify Spo0B in all but one Paenibacillus genome when we used the protein sequences of Spo0B from P. polymyxa and P. validus as the query.
Spo0B protein sequences are not well conserved. (A) Pairwise percent identity matrix of Spo0F, Spo0B, and Spo0A orthologs, calculated using Clustal Omega. (B) Protein sequence conservation plot of Spo0B-TM and Spo0B from Paenibacillus (n = 1,437). Residues shared by ≥75% of sequences are shaded dark blue. Features are labeled with respect to the Spo0B-TM sequence from the reference genome of P. polymyxa. Protein sequence accessions are listed in Table S5.
The sequence of Spo0B is known to be poorly conserved among Bacillus species (76, 77), and we determined this to be true among Paenibacillus species as well (Fig. 7B). In an alignment of 1,437 Spo0B-TM and Spo0B sequences from Paenibacillus (length = 246 ± 12 amino acids), six residues were 100% conserved (Fig. 7B). Five of these residues are part of the highly conserved region around the phosphorylatable histidine residue. Only 46 residues were conserved in >75% of sequences (Fig. 7B). The sequence of the transmembrane domain was not strongly conserved, although its hydrophobic properties are maintained, suggesting that the importance of this domain is membrane localization. Altogether, these data suggest that the phosphorylatable histidine domain is the most fixed feature of this protein, while the remaining sequence can evolve more freely.
DISCUSSION
Bacillota diversified from other phyla very early in evolutionary history (78), and endospore formation is likely an ancestral trait of this phylum. Thus, the ability to form spores encompasses diverse bacteria that inhabit a broad range of environments. Sporulation genes have largely been inherited vertically, with approximately 120 genes shared across endospore formers within Bacillota (23). Although previous studies included single representatives of Paenibacillus, we detected most of these genes in our expanded survey, which provides further support for their universal conservation. Core genes are involved in all stages of sporulation from initiation to germination, indicating that the fundamental process of sporulation is shared in most Bacillota. Nevertheless, even within this core sporulation genome, there is unexpected diversity that likely reflects differences in environmental contexts and signals triggering the decision to sporulate. The Paenibacillus genus was originally classified within Bacillaceae but has been subsequently reclassified (63). Our analysis of sporulation gene conservation across Paenibacillus spp. may aid in further resolution of their phylogenetic relationships.
The ancestral sporulation initiation pathway is hypothesized to have been a phosphorelay (25), in which the phosphotransferases Spo0F and Spo0B aid in the transfer of the phosphoryl group from a histidine kinase to Spo0A (Fig. 1). Spo0B, a cytoplasmic dimeric histidine transferase, may have evolved from a histidine kinase that lost its N-terminal signal detection domain and C-terminal ATP-binding domain (79). Spo0B ortholog sequences tend to vary greatly, even between species in the same genus (65, 76, 77) (Fig. 3B and 7). One of the unusual features of Spo0B identified in Paenibacillus is an N-terminal extension predicted by Park et al. (65) to encode a transmembrane domain. This domain is absent from Spo0B in B. subtilis. Park and colleagues found this domain in Spo0B sequences in a total of six Paenibacillus genomes. We confirmed that this predicted transmembrane domain does indeed facilitate membrane localization of P. polymyxa Spo0B-TM (Fig. 4). We identified Spo0B in all but one of the Paenibacillus genomes we surveyed (n = 1,460), and of these, 92% encode Spo0B-TM (Fig. 2A). Outside of Paenibacillus spp., we detected Spo0B-TM in other species in the Paenibacillaceae family and even in the neighboring families Thermoactinomycetaceae and Alicyclobacillaceae (Fig. 2B). The high degree of conservation in these families suggests that Spo0B-TM was present in their common ancestor, and the transmembrane domain was subsequently lost in certain lineages.
What is the importance of the transmembrane domain of Spo0B in Paenibacillus? Species from Paenibacillus lineages that encode Spo0B without the transmembrane domain have been experimentally shown to sporulate (80–82), which suggests that Spo0B lacking the transmembrane domain is functional in these lineages. Nevertheless, the high conservation of the transmembrane domain suggests it imparts a strong selective advantage in this genus. One possibility is that Paenibacillus may rely more heavily on the membrane-localized kinases (KinB, KinC, and KinD) than the cytoplasmic kinases (KinA and KinE) to initiate sporulation. B. subtilis is more dependent on KinB for sporulation in minimal media (83). KinA-E are highly conserved in Paenibacillus (Fig. 1; Table S2). If there are environmental conditions experienced by Paenibacillus that increase reliance on membrane-localized kinases to initiate sporulation, then it may be advantageous to localize a member of the phosphorelay to the membrane as well.
The transmembrane domain of Spo0B-TM may also enhance its interaction with Spo0A and Spo0F. Using a BACTH assay, we found that deleting the transmembrane domain eliminated detectable interaction with both Spo0A and Spo0F of P. polymyxa (Fig. 5B). In B. subtilis, Spo0B forms dimers through the interaction of the N-terminal helical domains, and dimerization is essential to form the four-helix bundle interaction site necessary for phosphoryl transfer (84). Therefore, the additional helices supplied by the transmembrane domain could help to stabilize dimer formation. Increased dimerization may enable detection by the BACTH assay, either through increased stabilization of the Spo0B protein itself or through stabilization of binding interactions with its phosphorelay partners.
Among members of the sporulation initiation phosphorelay, we found that Spo0B exhibits the most sequence diversity (Fig. 7A). The phosphorelay is an expanded version of a typical two-component system, and the interaction surface residues of the proteins in these systems resist evolutionary change. For example, 20 out of 21 interaction residues of Spo0F in B. subtilis, Bacillus anthracis, and Bacillus halodurans are identical, while only 50% of the remaining residues are identical (85). Accordingly, the most well-conserved region of Spo0B is the region surrounding the phosphorylatable histidine residue (Fig. 3B). Spo0B orthologs have a much lower sequence conservation, and in some instances, have even acquired additional function. Spo0B in B. anthracis has pleiotropic functions, including phosphotransferase, autophosphorylation, and ATPase activity as a result of having acquired ATP-binding and hydrolysis domains (77). The physiological consequence of gaining ATPase function has not been determined, but it has been proposed that this expanded functionality could be important for vegetative growth or even pathogenesis.
Our study highlights the importance of studying sporulation proteins in non-model organisms, since variations can be identified even in highly conserved sporulation genes. Paenibacillus species inhabit diverse environments and cause a broad range of industrial and public health impacts, from milk spoilage to beehive collapse to human infections. And yet, the majority of what we know about endospore formation comes from B. subtilis, mainly due to its genetic tractability. The growing number of Paenibacillus genomes available will greatly facilitate future surveys of this kind. Moreover, the wealth of environmental isolates that can be obtained from environmental sources makes Paenibacillus an attractive and underappreciated model of sporulation with important industrial and clinical applications.
MATERIALS AND METHODS
Strains and media
B. subtilis strains were derived from 168 trpC2 (86) and grown in Lysogeny Broth (LB) at 37°C with aeration. Genomic DNA from the ∆spo0B::kan strain from the BKK collection (87) was used to create the ∆spo0B strains by transformation into 168 trpC2. Plasmids used can be found in Table 1, and novel plasmid sequences are available on the project GitHub at https://github.com/isabella-n-lin/paenibacillus_sporulation/blob/main/paeni_plasmid_seqs.fasta. Antibiotics were used at final concentrations of 100 µg/mL ampicillin, 50 µg/mL kanamycin, and 100 µg/mL spectinomycin. P. polymyxa ATCC 842 was obtained from the American Type Culture Collection and grown in Brain Heart Infusion (BHI) broth at 30°C with aeration. All experiments were performed with three biological replicates, unless otherwise stated.
Paenibacillus sporulation pangenome and phylogenetic tree building
All available Paenibacillus genome assemblies as of 21 April 2025 were downloaded from the NCBI Reference Sequence (RefSeq) collection as annotated by the NCBI Prokaryotic Genome Annotation Pipeline (PGAP) (89). Genomes with CheckM (90) contamination greater than 5% or completeness less than 95% were removed. Any genomes suppressed by NCBI by 28 July 2025 were removed, resulting in 1,460 genomes. Genome assembly numbers can be found in Table S1. The phylogenetic tree was made with GToTree v1.8.14 (91), using the prepackaged single-copy gene set for bacteria (74 target genes). The tree was visualized using the ggtree v3.10.1 package (92) and midpoint rooted using the phangorn v2.12.1 package (93). A comprehensive list of sporulation genes was compiled from previous work (21, 23) and SubtiWiki (59) (Table S2). B. subtilis sporulation protein sequences were obtained from SubtiWiki (59). Sporulation genes were detected in Paenibacillus genomes using BLASTP v2.16.0 (94) and an E-value cutoff of 0.05.
Spo0B search and transmembrane-domain prediction
HMMER v3.4 (phmmer) (95) was used to search for Spo0B across all proteins annotated by the NCBI PGAP pipeline in the previously described Paenibacillus data set (Table S1) and all reference genomes in the class of Bacilli from the NCBI RefSeq database (downloaded on 5 November 2025) (Table S4). Spo0B sequences from P. polymyxa and P. validus were used as the phmmer query for Paenibacillus spp., and Spo0B sequences from P. polymyxa, P. validus, B. subtilis, and Brevibacillus brevis were used as the query for Bacilli. Spo0B protein accessions for the queries can be found in Table S5. DeepTMHMM v1.0.42 (96) was used to predict transmembrane domains.
Spo0F, Spo0B, and Spo0A sequence analysis and structure modeling
The Kyte-Doolittle hydropathy plot was created using ProtScale on the ExPASy server (97) using the Spo0B sequence WP_019688407.1. AlphaFold models were created using AlphaFold 3 (98) and visualized using ChimeraX v1.9 (99). Protein sequences of Spo0F, Spo0B, and Spo0A were obtained from NCBI (100), and the accession numbers can be found in Table S5. Percent identity was calculated using Clustal Omega v1.2.4 (101). The alignment in Fig. 3B was made using Clustal Omega v1.2.4 (101) and visualized using the pyMSAviz v0.5.0 package (102). The conservation plot in Fig. 7B was created using the bio3d v.2.4-5 package (103). Spo0B sequences for the conservation plot were aligned using MAFFT v7.520 (104), using a gap opening penalty of 3 and gap extension penalty of 0.3.
Membrane localization microscopy
B. subtilis overnight cultures were normalized to an OD600 of 0.05 and grown to an OD of 1.5 at 37°C with aeration in LB with 1% xylose for induction of GFP-tagged proteins. For microscopy, cells were harvested, washed with 1× phosphate-buffered saline, and stained with fluorescent dye FM4-64 (Invitrogen T3166) for 5 min at room temperature. Stained cells were mounted on 1% agarose pads for imaging. Microscopy was performed with a Nikon ECLIPSE Ni-E upright microscope with GFP-FITC and RFP-TRITC filter cubes and a Prime SBI Express camera. Exposure times for GFP, FM4-64, and phase microscopy were 700, 100, and 26 ms, respectively. Images were processed using ImageJ v1.53k (105). GFP fluorescence is autoscaled in each image.
Membrane fractionation
B. subtilis overnight cultures were normalized to an OD600 of 0.05 and grown to an OD of 1.5 at 37°C with aeration in LB with 1% xylose for induction of GFP-tagged proteins. Cells were centrifuged at 10,000 × g for 15 min at room temperature (Sorvall LYNX 4000, Fiberlite F12-6x500 LEX rotor), resuspended in Buffer A (50 mM Tris-HCl, pH 7.5, 150 mM NaCl, 5 mM MgCl2, 10% glycerol v/v), and flash-frozen by dripping into liquid nitrogen. Cells were lysed by cryomilling at 15 Hz for 3 min, repeated 3 times (Retsch Mixer Mill MM 400), and lysates were clarified by centrifuging at 10,000 rpm for 10 min at 4°C (Beckman Coulter Avanti J-15R, rotor JA-10.100). To pellet membranes and their associated proteins, clarified lysates were centrifuged at 45,000 rpm for 1 h at 4°C (Beckman Coulter Optima XE-90 Ultracentrifuge, TI70 rotor). Membrane pellets were visualized using a BioRad ChemiDoc Imaging System and the Alexa Fluor 488 setting.
Bacterial two-hybrid system
To investigate the protein interactions of Spo0B-TM from P. polymyxa, we used the commercially available BACTH System Kit from Euromedex. Strains were derived from the non-reverting adenylate cyclase-deficient E. coli strain BTH101. Plasmid gene inserts were amplified from P. polymyxa ATCC 842. All proteins of interest were C-terminally tagged with either T18 or T25 fragments.
For the β-galactosidase assay, strains were grown in LB supplemented with ampicillin, kanamycin, and 1% glucose for 6 h at 30°C. The cultures were used to inoculate LB supplemented with ampicillin, kanamycin, and 0.5 mM IPTG and grown for 18 h at 30°C. The cells were lysed using lysis buffer (100 mL Z-buffer (0.06M Na_2_HPO_4_, 0.04M NaH_2_PO_4_-H_2_O, 0.01M KCl, 0.001M MgSO_4_-7H_2_O), 270 μL β-mercaptoethanol, 50 μL 10% sodium dodecyl sulfate) and 20 μL chloroform and incubated at 30°C. The reaction was started upon the addition of o-nitrophenyl-β-D-galactopyranoside (4 mg/mL in Z-buffer) to the lysed cells. The reaction was stopped with 1M Na_2_CO_3_ when a pale yellow color developed. Absorbances at 600, 420, and 550 nm were measured with a BioTek Synergy H1 microplate reader, Gen 5 3.11, for cell density, o-nitrophenol production, and background, respectively. β-galactosidase activity was measured in Miller units. P-values were calculated with the R stats v4.3.2 package using a one-way ANOVA followed by Tukey’s test. Experiments were performed using nine biological replicates.
Sporulation assays
Sporulation was induced by nutrient exhaustion in Difco sporulation medium (DSM). Strains were grown in DSM on a roller drum at 37°C for 3 h, then normalized to OD600 of 0.05 and incubated at 37°C, shaking at 220 rpm for 24 h. The cultures were serially diluted in Tbase + 1 mM MgSO_4_ and plated on LB agar plates before and after heating for 20 min at 80°C.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Galperin MY. 2013. Genome diversity of spore-forming firmicutes. Microbiol Spectr 1:TBS–0015 doi:10.1128/microbiolspectrum.TBS-0015-201226184964 PMC 4306282 · doi ↗ · pubmed ↗
- 2Yutin N, Galperin MY. 2013. A genomic update on clostridial phylogeny: gram-negative spore formers and other misplaced clostridia. Environ Microbiol 15:2631–2641. doi:10.1111/1462-2920.1217323834245 PMC 4056668 · doi ↗ · pubmed ↗
- 3Setlow P, Christie G. 2023. New thoughts on an old topic: secrets of bacterial spore resistance slowly being revealed. Microbiol Mol Biol Rev 87:e 0008022. doi:10.1128/mmbr.00080-2236927044 PMC 10304885 · doi ↗ · pubmed ↗
- 4Tan IS, Ramamurthi KS. 2014. Spore formation in Bacillus subtilis. Environ Microbiol Rep 6:212–225. doi:10.1111/1758-2229.1213024983526 PMC 4078662 · doi ↗ · pubmed ↗
- 5Khanna K, Lopez-Garrido J, Pogliano K. 2020. Shaping an endospore: architectural transformations during Bacillus subtilis sporulation. Annu Rev Microbiol 74:361–386. doi:10.1146/annurev-micro-022520-07465032660383 PMC 7610358 · doi ↗ · pubmed ↗
- 6Khanna K, Lopez-Garrido J, Zhao Z, Watanabe R, Yuan Y, Sugie J, Pogliano K, Villa E. 2019. The molecular architecture of engulfment during Bacillus subtilis sporulation. elife 8:e 45257. doi:10.7554/e Life.4525731282858 PMC 6684271 · doi ↗ · pubmed ↗
- 7Broder DH, Pogliano K. 2006. Forespore engulfment mediated by a ratchet-like mechanism. Cell 126:917–928. doi:10.1016/j.cell.2006.06.05316959571 PMC 3266857 · doi ↗ · pubmed ↗
- 8Shuster B, Khemmani M, Abe K, Huang X, Nakaya Y, Maryn N, Buttar S, Gonzalez AN, Driks A, Sato T, Eichenberger P. 2019. Contributions of crust proteins to spore surface properties in Bacillus subtilis. Mol Microbiol 111:825–843. doi:10.1111/mmi.1419430582883 PMC 6417949 · doi ↗ · pubmed ↗