Isolation and genomic analysis of secondary metabolism in cave Actinomycetota from biofilms and Ceuthophilus
Ilyana O. Bachmann, Brian O. Bachmann

TL;DR
This paper describes the isolation and genomic study of Actinomycetota from cave biofilms and crickets, revealing new bacterial diversity and secondary metabolism.
Contribution
The study provides novel genomic insights into cave Actinomycetota and their secondary metabolism potential.
Findings
Nine Actinomycetota strains were isolated from cave biofilms and crickets.
Genomic analysis revealed diverse genera with low DNA-DNA hybridization to known species.
The study highlights the secondary metabolism potential of cave Actinomycetota.
Abstract
We report the isolation and genomic sequencing of nine Actinomycetota obtained from both cave biofilms and trogloxenic Ceuthophilus (cave crickets). Strains were isolated from samples via actinomycetota-selective media and sequenced using nanopore sequencing. Provisional taxonomic inference of strains was performed by whole genome phylogenetic analysis, revealing a broad range of genera, with most strains clustering with known genera with digital DNA-DNA hybridization values less than 70%, and one strain not clustering with known genera. An analysis of secondary metabolism reveals the quantity and diversity of secondary metabolism in cave Actinomycetota.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
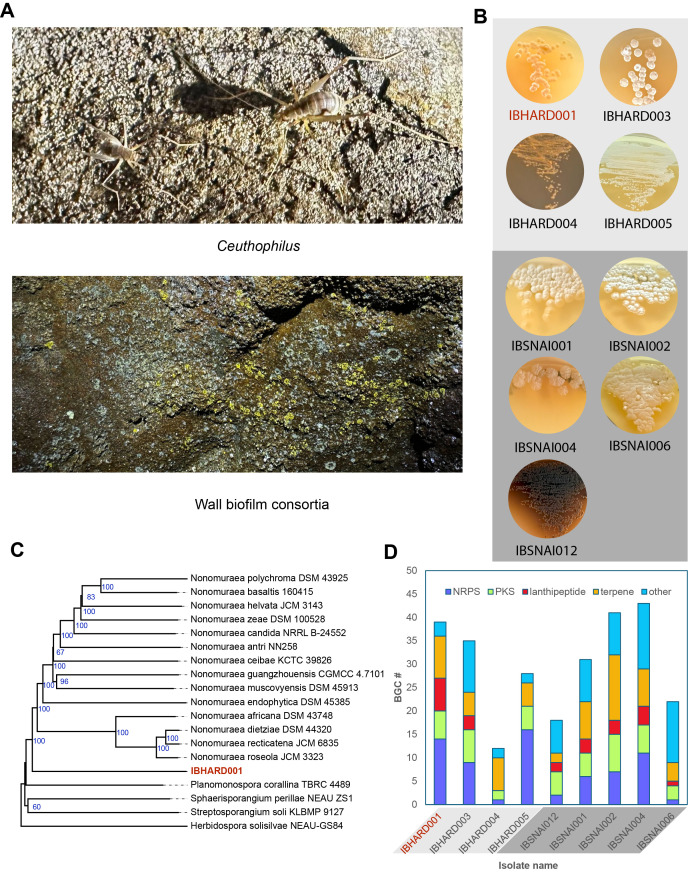 Figure 1
Figure 1|
| ||||||||
|
|
|
|
|
|
|
|
|
|
|
IBHARD001 |
248,811 |
97 |
9.5 |
2 |
8,494 |
|
|
PRJNA1308599 |
|
IBHARD003 |
221,983 |
75 |
8.3 |
2 |
7,251 |
|
28.6 |
PRJNA1308605 |
|
IBHARD004 |
380,186 |
99 |
6.8 |
1 |
6,546 |
|
66.8 |
PRJNA1308606 |
|
IBHARD005 |
266,653 |
102 |
6.3 |
1 |
5,866 |
|
44.4 |
PRJNA1308607 |
|
IBSNAI012 |
352,887 |
102 |
6.6 |
1 |
6,126 |
|
68.5 |
PRJNA1308591 |
|
IBSNAI001 |
316,549 |
101 |
8.5 |
4 |
7,810 |
|
69.7 |
PRJNA1308129 |
|
IBSNAI002 |
168,615 |
92 |
9.9 |
8 |
9,065 |
|
34.9 |
PRJNA1308159 |
|
IBSNAI004 |
69,803 |
45 |
9.4 |
9 |
8,707 |
|
46.7 |
PRJNA1308588 |
|
IBSNAI006 |
378,350 |
98 |
9.9 |
2 |
8,943 |
|
73.5 |
PRJNA1308590 |
|
1
IBHARD strains were isolated from Hardin’s cave
| ||||||||
- —National Cancer Institute (United States)https://ror.org/040gcmg81
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsSubterranean biodiversity and taxonomy · Actinomycetales infections and treatment · Genomics and Phylogenetic Studies
Description
The Actinomycetota phylum has been historically prolific in producers of bioactive secondary metabolites with clinical application (Yi, Shi, Cao, & Pan, 2025). Many antibiotics and anticancer drugs are bacterial secondary metabolites, derived from secondary metabolites, or address a target of a secondary metabolite (Newman & Cragg, 2020). For example, while first discovered in fungal strains, the β-lactam antibiotics, including penicillins, cephalosporins, clavams, and carbapenems, are also produced by various strains of Actinomycetota (Lewis et al., 2024). Anticancer secondary metabolites produced by Actinomycetota have roles in the treatment of a variety of cancer types. Daunomycin, used in the treatment of Acute Myeloid Leukemia and other cancers (Lewis et al., 2024), is produced by *Streptomyces * (Hutchinson, 1997). Carfilzomib, a derivative of epoxomicin, is a treatment for multliple myeloma produced by Streptomyces (Schorn et al., 2014). However, after decades of mining for bioactive secondary metabolites, the rates of discovery of novel compounds have slowed (Pye, Bertin, Lokey, Gerwick, & Linington, 2017).
Most Actinomycetota have been isolated from terrestrial ecosystems. The slowing rate of compound discovery is correlated with the slowing rate of discovery of novel Actinomycetota. It has been demonstrated that novel microbial taxa are more likely to produce novel chemistry (Yi et al., 2025). Moreover, metagenomic studies suggest a large reservoir of Actinomycetota remains to be cultivated (Bull & Goodfellow, 2019). Novel taxa can be discovered by developing new culture techniques or isolating strains from previously uninvestigated ecosystems.
In previous work, we have isolated novel bioactive secondary metabolites from cave actinomycetota (Covington, Spraggins, Ynigez-Gutierrez, Hylton, & Bachmann, 2018; Derewacz et al., 2014; Earl et al., 2018). Herein, we investigate the potential of cave ecosystems for the discovery of Actinomycetota strains with the potential to produce secondary metabolites. Our goal was to investigate two sources, cave microbial biofilms and trogloxenic insects, in particular cave crickets ( Ceuthophilus ), via Actinomycetota selective culture conditions, nanopore sequencing of isolated strains, and analysis of secondary metabolic biosynthetic gene clusters.
Two caves were targeted for this study. Hardin’s Cave, privately owned, and Snail Shell Cave, managed by the Southeastern Cave Conservancy. Hardin's Cave is a 3.5-mile relatively dry cave. Snail Shell cave comprises over nine miles of surveyed passages and is characterized by extensive flowing water passages, many of which require a boat for passage. Both cave ecosystems utilize allochthonous carbon input from the forest soil ecosystem. To collect samples from Snail Shell cave (Figure A), we developed a biofilm collection methodology that targeted biofilm from cave walls. Ceuthophilus samples were generated from whole crickets to encourage the isolation of spore-forming Actinomycetota. Three soil samples and three crickets were used in isolation workflows.
A wide variety of methods have been developed for the isolation of Actinomycetota from environmental samples. We selected two methods to enrich for the isolation of actinomycetes. As described in Methods, we adapted a standard soil dilution and plating methodology using humic acid vitamin (HV) agar with added antibiotics to suppress fungi and Gram-negative microorganism growth (Hayakawa & Nonomura, 1987). Second, we adapted a methodology to enrich the presence of motile spores (Hayakawa, Otoguro, Takeuchi, Yamazaki, & Iimura, 2000; Hop et al., 2011).
Colonies formed in HV agar primary plates, and fast-growing colonies were picked and subcultured in as little as one week (Figure B), and slow-growing colonies were subcultured in as much as three weeks. Subculture plates of International Streptomyces Protocol 1 (ISP1) medium agar, typtic soy broth (TSB) agar, and Bennett’s agar were streaked to subculture picked colonies from primary plates onto secondary plates. Photographs of isolated Actinomycetota are shown in Figure B. Single colonies were used to subculture onto tertiary plates of the best medium for growth, and these were used to inoculate liquid cultures in ISP1 or Bennett’s medium. From 25 mL cultures, we prepared glycerol stocks and used the majority for genomic sequencing.
Genomes of nine selected isolated strains sequenced by nanopore sequencing using Oxford Nanopore sequencing at Plasmidsaurus Inc. We evaluated two methodologies for sequencing samples. Genomic DNA from four liquid cultures was isolated using a classic protocol optimized for bulk isolation of genomic DNA from actinomycetes. Isolated DNA was of high purity (1.96- 2.00 A 260 /A 280 , 1.97 -2.07 A 260 /A 230 ) and submitted for sequencing. Alternatively, for six liquid cultures, we submitted cell pellets for automated DNA isolation using a ZymoBIOMICS™ 96 MagBead DNA Kit and nanopore sequencing. As shown in Table 1, we obtained high-quality assemblies for all samples. All genomes were of sufficient quality to permit taxonomic estimation and evaluation of secondary metabolic potential.
Taxonomic estimation via whole gene taxonomic analysis using the Type (Strain) Genome Serve (TYGS), which estimates taxonomic assignments and phylogenies for submitted strains in comparison to a continuously updated database of type strains. Taxonomic distance is proportional in part to the Digital DNA-DNA Hybridization (sDDH) score shown, with all strains below the cutoff for preliminary designation as novel species. Figure C shows an example of phylogenetic analysis of strain IBHARD001, isolated from a cave cricket and analyzed by whole genome taxonomy using TYGS. This shows the tree inferred with FastME 2.1.6.1 (Lefort et al., 2015) from GBDP distances calculated from genome sequences. The branch lengths are scaled in terms of GBDP distance formula * d 5 * and indicate that this strain does not cluster with any of the species that it is most similar to, suggesting it may be a member of a previously undescribed genus. Future studies will be required to validate this preliminary assignment.
Secondary metabolic potential of genomically sequenced strains was performed using the antiSMASH program, which uses GenBank-formatted data to analyze for the presence of secondary metabolic biosynthetic gene clusters (BGCs). Gene clusters are classified into biosynthetic classes based on the predicted presence of diagnostic genes for each class. From antiSMASH analysis of the nine genomes, we tabulated the number of BGCs in four major classes. As shown in Panel D, isolates contain between 12 and 43 predicted BGCs, the number of which roughly scales with genome size. Number and types of biosynthetic classes varied between strains.
This pilot study defines the potential for the discovery of new taxa with secondary metabolic potential from cave ecosystems and trogloxenic insects. Future studies with a broader range of culture conditions and sample treatments can expand the scope of microorganismal discovery. Preliminary species and genera can be validated by follow-up studies. Promising strains containing biosynthetic gene clusters of interest can enter into a molecular discovery workflow to assess and validate the potential of novel cave taxa for generating novel bioactive secondary metabolites as leads for development in drug discovery programs.
Methods
Cave environmental sample collection
Sediment samples were collected from Snail Shell Cave, a hydrologically active cave located in Rockvale, Tennessee. The samples were collected immediately after the twilight zone, in the dark zone, from cave walls that demonstrated evidence of microbial colonization. A one cm ^2^ sterile abrasive pad (Scotch-Brite) was used to collect biofilm sediments by scrubbing the wall in a circle with a ten cm diameter. Five sediment samples were collected and transported in sterile Falcon tubes, transferred into sterile petri dishes, and dried at 30 °C for two weeks. Dry weights were 215 - 553 mg. Sediment was removed from the pads aseptically by stretching and bending the pads. Cave-dwelling crickets were collected from Hardin’s Cave, located in Nashville, Tennessee. The crickets were captured in sealable plastic bags and crushed within ten minutes of capture. The crickets were then cut into fine pieces in a Petri dish, at 30 °C for one week to induce sporulation and further cut with a sterile razor blade to produce a coarse powder.
Selective isolation method
Sediments and crickets were plated using two selective isolation methods. For both methods, the samples were first resuspended in 100 mL beakers (60 x 46 mm diameter) in 50 mL phosphate buffer (50 mM, pH = 7.0) containing 10 % soil extract, stirring with a glass rod for 30 seconds, followed by 90 minutes of rest at 30 °C. In the “motile spore preparation” method (Hayakawa et al., 2000; Hop et al., 2011), 8 mL of the upper portion of the sample solution was transferred into a 15 mL Falcon tube (16 x 105 mm). The tubes were centrifuged for 10 minutes at 1500 x g and allowed to rest at 30 ºC for 30 minutes to enable motile spores to circulate. From the middle of the volume, 3 mL was removed to a sterile Eppendorf tube and ten-fold serially diluted twice. In the “standard dilution method,” the above 90-minute rested solution was briefly stirred, allowed to settle for 30 seconds, and the suspension was diluted with sterile H 2 O, 1:100, 1:1K, and 1:10K dilutions.
All samples were plated on Humic Acid Vitamin agar (Hayakawa & Nonomura, 1987), ISP1, and Bennett’s Agar supplemented with a final concentration of nalidixic acid (20 mg/L), trimethoprim (10 mg/L), and cycloheximide (50 mg//L) to inhibit the growth of fungi and Gram-negative bacteria. The plates were incubated for 2 - 4 weeks at 30 ºC, and colonies of interest were picked and inoculated onto secondary purification plates by streaking onto ISP1 and Bennett’s media Agar. Individual colonies were used to streak tertiary isolation plates. Loops of mycelia from tertiary plates were used to inoculate ISP1 and Bennett’s liquid cultures (25 mL in a 250 mL Erlenmeyer flask). After 5-7 days of incubation with orbital shaking (230 rpm). A 0.5 mL portion of the culture was combined with an equal volume of sterilized 20% glycerol, 10% lactose, and stored at -80 °C. The remaining cultures were pelleted by centrifugation at 3348 x g for 20 minutes and stored at -80 ºC.
Isolation of genomic DNA
Genomic DNA was isolated using a procedure adapted from standard protocols (Kieser, J., Buttner, Chater, & Hopwood, 2000). The frozen bacterial pellets were resuspended in 5 mL TE25S buffer. 100 µL of Lysozyme solution (100 mg mL ^-1^ in nuclease-free H 2 O) and 200 µL/mL RNAse A (0.004 g in TE25S Buffer) were added. The sample was incubated for 60 minutes at 37 ºC, with occasional inversions. Then 50 µL Proteinase K solution (0.02 g in 1 mL H 2 O) was added and then mixed. 300 µL of SDS solution was added, and the sample was incubated for one hour at 55 ºC in a water bath, with occasional inversion. Then 1 mL of 5M NaCl and 650 µL CTAB/NaCl were added and mixed in, and the sample was incubated for 10 minutes at 55ºC. The samples were cooled to about 37 ºC, and 5 mL chloroform/isoamyl alcohol (24:1) was added and mixed via inversion for 30 minutes. The samples were centrifuged for 15 minutes at 3750 rpm, and the top layer was transferred to a fresh Falcon tube, being careful not to pull the interface layer. Then 0.6 volume of isopropanol was added and mixed by inversion. After about 3 minutes, the DNA was precipitated. Then the DNA was spooled with a glass hook and rinsed in 70% Ethanol. The DNA was transferred to a clean Eppendorf tube and air-dried for about 2.5 minutes. The DNA was then dissolved in 250 µL EB Buffer and heated to 55 ºC for 30 minutes. The DNA was incubated at four ºC for 3 days to fully rehydrate and homogenized by pipetting up and down using a 200 μL micropipette.
Nanopore sequencing
Genomic DNA was isolated from IBSNAI001, IBSNAI002, IBSNAI004, and IBSNAI006 using a modified CTAB protocol from 25 mL cultures, which were diluted to 50 ng/μL. Pellets of IBHARD001, 003, 004, 005, and IBSNAI012 were generated from 25 mL liquid cultures. Bacterial Genome Sequencing for IBSNAI001, IBSNAI002, IBSNAI004, and IBSNAI006 was performed on isolated DNA by Plasmidsaurus Inc. using Oxford Nanopore Technology with custom analysis and annotation. Bacterial Genome Sequencing for IBHARD001, 002, 003, 004, 005, and for IBHARD012 was performed from pellets (30 - 50 mg), genomic DNA isolated using ZymoBIOMICS™ 96 MagBead DNA Kit (D4308/D4302) and sequenced as above.
Taxonomic estimation
The genome sequence data were uploaded to the Type (Strain) Genome Server (TYGS), a free bioinformatics platform available under https://tygs.dsmz.de, for a whole genome-based taxonomic analysis (Meier-Kolthoff & Goker, 2019). The analysis also made use of recently introduced methodological updates and features. Information on nomenclature, synonymy, and associated taxonomic literature was provided by TYGS's sister database, the List of Prokaryotic names with Standing in Nomenclature (LPSN, available at https://lpsn.dsmz.de) (Meier-Kolthoff, Carbasse, Peinado-Olarte, & Goker, 2022). The results were provided by the TYGS on 2025-08-11.
Data availability
GenBank accession numbers are provided in Table 1.
**: **
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Blin K Shaw S Vader L Szenei J Reitz ZL Augustijn HE Cediel-Becerra JDD de Crécy-Lagard V Koetsier RA Williams SE Cruz-Morales P Wongwas S Segurado Luchsinger AE Biermann F Korenskaia A Zdouc MM Meijer D Terlouw BR van der Hooft JJJ Ziemert N Helfrich EJN Masschelein J Corre C Chevrette MG van Wezel GP Medema MH Weber T 202577 anti SMASH 8.0: extended gene cluster detection capabilities and analyses of chemistry, enzymology, and regulation.Nucleic Acids Res 53W 10305-1048 W 32W 3810.1093/nar/gkaf 33440276974 PMC 12230676 · doi ↗ · pubmed ↗
- 2Bull AT Goodfellow M 2019121 Dark, rare and inspirational microbial matter in the extremobiosphere: 16 000 m of bioprospecting campaigns.Microbiology (Reading)165121350-08721252126410.1099/mic.0.00082231184575 · doi ↗ · pubmed ↗
- 3Covington BC Spraggins JM Ynigez-Gutierrez AE Hylton ZB Bachmann BO 2018917 Response of Secondary Metabolism of Hypogean Actinobacterial Genera to Chemical and Biological Stimuli.Appl Environ Microbiol 84190099-224010.1128/AEM.01125-1830030223 PMC 6146984 · doi ↗ · pubmed ↗
- 4Derewacz DK Mc Nees CR Scalmani G Covington CL Shanmugam G Marnett LJ Polavarapu PL Bachmann BO 2014721 Structure and Stereochemical Determination of Hypogeamicins from a Cave-Derived Actinomycete.J Nat Prod 7780163-38641759176310.1021/np 400742 p 25046128 PMC 4334282 · doi ↗ · pubmed ↗
- 5Earl DC Ferrell PB Jr Leelatian N Froese JT Reisman BJ Irish JM Bachmann BO 201812 Discovery of human cell selective effector molecules using single cell multiplexed activity metabolomics.Nat Commun 91393910.1038/s 41467-017-02470-829295987 PMC 5750220 · doi ↗ · pubmed ↗
- 6Farris James S. 197291 Estimating Phylogenetic Trees from Distance Matrices The American Naturalist 1069510003-014764566810.1086/282802 · doi ↗
- 7Hayakawa Masayuki Nonomura Hideo 198711 Humic acid-vitamin agar, a new medium for the selective isolation of soil actinomycetes Journal of Fermentation Technology 6550385-638050150910.1016/0385-6380(87)90108-7 · doi ↗
- 8Hayakawa M Otoguro M Takeuchi T Yamazaki T Iimura Y 200081 Application of a method incorporating differential centrifugation for selective isolation of motile actinomycetes in soil and plant litter.Antonie Van Leeuwenhoek 7820003-607217118510.1023/a:102657942626511204769 · doi ↗ · pubmed ↗