Supplementation with commercially available probiotic bacterial strains reduces fat accumulation in Caenorhabditis elegans by regulating the lipid metabolism pathway
Ryuichi Saito, Rika Inomata, Dian-Sheng Wang, Satoshi Shimazaki

TL;DR
This study shows that certain probiotic bacteria reduce fat accumulation in a type of worm by influencing lipid metabolism.
Contribution
The novel finding is that EFT110, BSTOA, and CBTOA reduce fat accumulation in C. elegans by regulating lipid metabolism genes.
Findings
EFT110, BSTOA, and CBTOA significantly reduce fat accumulation in C. elegans.
These probiotics regulate gene expression in lipid metabolic pathways.
Abstract
Enterococcus faecium T-110 (EFT110), Bacillus subtilis TO-A (BSTOA), and Clostridium butyricum TO-A (CBTOA) are probiotic bacteria used as supplements to improve intestinal symptoms in humans, chickens, pigs, cattle, and fish. However, their effects on hosts remain unclear. Our previous study showed that EFT110, BSTOA, and CBTOA extend the lifespan of Caenorhabditis elegans . In this study, we investigated the effects of these three strains on fat accumulation in nematodes; and observed that each bacterium significantly reduced fat accumulation by regulating gene expression in lipid metabolic pathways.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
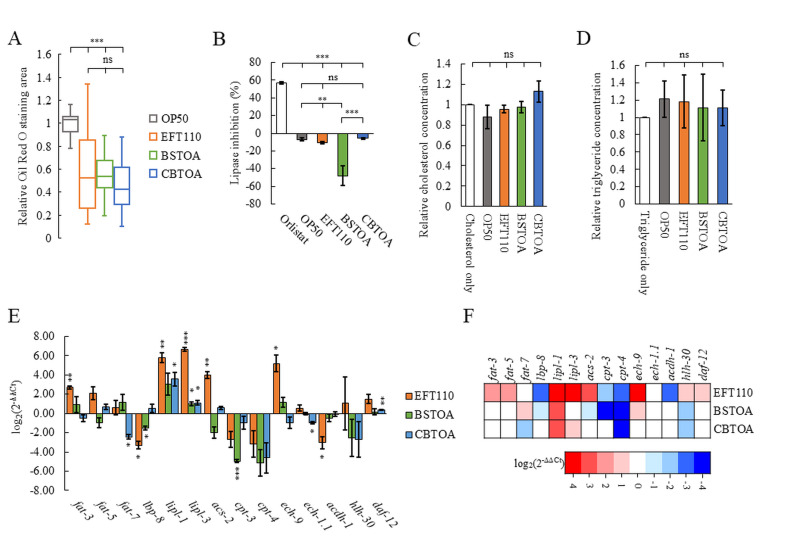 Figure 1
Figure 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGenetics, Aging, and Longevity in Model Organisms · Rabbits: Nutrition, Reproduction, Health · Microbial Metabolism and Applications
Description
EFT110, BSTOA, and CBTOA are commercially available as combined-probiotic preparation (BIO-THREE®) used since 1963 to improve intestinal symptoms in humans. These strains are used as dietary supplements and as probiotic additives in livestock and aquaculture. These strains effectively alleviate constipation, diarrhea, irritable bowel syndrome, inflammatory diseases, and infectious diseases (Chen et al., 2010, Huang et al . , 2014; Inatomi et al . , 2017; Nagamine, 2021; Shiozaki et al., 2014, Tsuda et al . , 2007) owing to reduced dysbiosis (Inatomi and Honma 2021; Shiozaki et al., 2014), enhanced short-chain fatty acid production (Honda et al., 2025; Inatomi and Honma 2023; Xu et al., 2021), and inhibition of inflammation-related gene expression (Takaoka et al., 2024). We previously reported that these strains extend the lifespan of * Caenorhabditis elegans * (Saito et al., 2023), but their broader health effects remain unclear. Obesity is a major risk factor for diabetes, hypertension, coronary artery disease, stroke, and cancer (Apovian, 2016), with incidence rising rapidly (Phelps et al., 2024). Interest in probiotic strategies has grown, as many studies report reduced fat accumulation by probiotics. Therefore, we hypothesized that EFT110, BSTOA, and CBTOA could reduce fat accumulation.
Under laboratory conditions, nematodes are typically fed * Escherichia coli * OP50 ( OP50 ), producing a microbiota composed only of OP50 . Nematodes have a short lifespan (approximately 15 days), and their transparent bodies allow real-time tissue imaging. Notably, over 70% of lipid metabolism genes have human orthologs (Zhang et al., 2013), and their fat droplets contain triglycerides, phospholipids, sphingolipids, and sterols, similar to humans. Thus, nematodes are widely used as well-established models in obesity research (Ashrafi, 2007). In this study, we assessed fat accumulation in nematodes ingesting EFT110, BATOA, or CBTOA; and evaluated the ability of these probiotic strains to reduce host fat storage.
After 24 h of feeding, fat accumulation was significantly reduced compared to nematodes fed OP50 (set at 1.00 ± 0.024), with relative ratios of 0.563 ± 0.077, 0.541 ± 0.039, and 0.459 ± 0.053, respectively ( Figure 1A ). Dietary fat is typically hydrolyzed by lipase into free fatty acids and monoglycerides for absorption. Although the mechanism in nematodes remain unclear, the observation that orlistat reduces fat accumulation, as in humans, suggests a similar intestinal lumen-based hydrolysis and uptake (Savova et al., 2022). Orlistat, an anti-obesity drug, inhibited porcine pancreatic lipase-catalyzed hydrolysis of p-nitrophenyl butyrate by 56.9 ± 1.31% ( Figure 1B ). In contrast, EFT110 and CBTOA showed no inhibition, while BSTOA significantly enhanced lipase activity by 47.9 ± 10.8% ( Figure 1B ).
To evaluate the fat-degrading activity of the probiotic strains, each strain was incubated for 24 h in M9 buffer supplemented with cholesterol or triglycerides. None degraded cholesterol or triglycerides under our conditions ( Figure 1C and D). These results suggest that these bacteria alone cannot degrade fats in the intestinal lumen and the host lipase is required for the fat degradation. Therefore, we hypothesized that these strains reduce fat by modulating fat synthesis, storage, or degradation pathways in nematodes. To test this, we performed RT-qPCR on 31 lipid metabolism genes in nematodes fed each strain (DuMez-Kornegay et al . , 2024;Karyn et al . , 2008; Lu et al . , 2022;Noble et al . , 2013;Nomura et al . , 2010; Savova et al . , 2022; Yue et al . , 2021; Yu et al . , 2021; Sternberg et al., 2024). Figures 1E and 1F show genes whose expression levels changed by more than two-fold (log₂ 2 ^-ΔΔCt^
1 or <-1) or showed significant differences compared to the nematodes fed OP50 . In the EFT110-fed group, expression of
fat-3 * (fatty acid desaturase, involved in fatty acid desaturation), * lipl-1 * and * lipl-3 * (lipase, associated with fat catabolism), * acs-2 * (acyl-CoA synthetase, involved in fatty acid b-oxidation and synthesis), and * ech-9 * (enoyl-CoA hydratase, involved in fatty acid b-oxidation) were significantly increased, whereas * lbp-8 * (fat-binding protein, implicated in fatty acid transport) and * acdh-1 * (acyl-CoA dehydrogenase, involved in fatty acid b-oxidation) were decreased. In the BSTOA-fed group, * lipl-3 * increased, whereas * lbp-8 * and * cpt-3 * (carnitine palmitoyl transferase, fatty acid β-oxidation) decreased. Although * lipl-3 * expression was higher in BSTOA fed nematodes, the fold change was <2 (log₂ 2 ^-ΔΔCt^ =0.980 ± 0.184), suggesting a limited role in BSTOA-induced fat reduction. In the CBTOA-fed group, * lipl-1 * , * lipl-3 * , and * daf-12 * (nuclear receptor, fat metabolism) increased, whereas * fat-7 * (fatty acid desaturase, fatty acid desaturation) and * ech-1.1 * (enoyl-CoA hydratase, fatty acid β-oxidation) decreased. Although * ech-1.1 * and * daf-12 * expression was significantly changed in CBTOA fed nematodes, the fold changes were <2 (log₂ 2 ^-ΔΔCt^ =-0.955 ± 0.127 and 0.374 ± 0.026, respectively), suggesting a limited role in CBTOA-induced fat reduction. Most altered genes were linked to fat degradation, indicating these strains may promote fat reduction via lipid catabolism though the gene profiles suggest distinct mechanisms. Identifying active factors and expanding gene expression analyses may clarify the mechanisms underlying fat reduction. Moreover, as nematodes lack organs such as adipocytes, liver, and pancreas, it remains necessary to determine whether similar effects occur in other host organisms. Such validation could broaden the potential applications of these probiotics.
Methods
** * C. elegans maintenance and bacterial culture conditions * **
C. elegans * N2 Bristol and OP50 were purchased from the Caenorhabditis Genetics Center (CGC). Worms were maintained at 20°C on 60 mm nematode growth medium (NGM) agar plates (3 g NaCl, 17 g agar, 2.5 g peptone, 25 mM KPO 4 buffer, 1 mM MgSO 4 , 1 mM CaCl 2 , 5 µg·mL ^−1^ cholesterol in ethanol, and 975 mL H 2 O) seeded with OP50 . All experiments were conducted using adult nematodes 3 days post-hatching. OP50 or BSTOA were cultured in 5 mL of Luria-Bertani (1 g tryptone, 0.5 g yeast extract, 0.5 g NaCl, and 100 mL sterile water, pH 7.0) or Tryptic Soy Broth, respectively, under aerobic conditions at 37°C by shaking for 24 h. TOAT110 or CBTOA were cultured statically in 5 mL de Man Rogosa Sharpe (MRS) or gifu aerobic medium (GAM) broth, respectively, under anaerobic conditions for 24 h. Cells were harvested by centrifugation, washed three times with M9 buffer (3 g KH 2 PO 4 , 6 g Na 2 HPO 4 , 5 g NaCl, 0.02% gelatin, 1 mM MgSO 4 , and 1 L H 2 O), and collected as pellets.
** Oil Red O staining and quantitation **
Oil Red O staining was performed as described (Aarnio et al., 2011; Eyleen et al., 2009; Wang and Ching, 2021). Briefly, 2× MRWB (160 mM KCl, 40 mM NaCl, 357.5 mg EGTA, 266.3 mg HEPES, 50 mL H 2 O, pH 7.4) was supplemented immediately with 0.2% β-mercaptoethanol, 2% paraformaldehyde, 1 mM spermidine, and 0.4 mM spermine, before use. Nematodes were cultured on 60 mm plates with modified NGM without peptone onto 200 mL of bacterial suspension was uniformly spread (10 mg mL⁻ ^1^ in M9 buffer), at 20°C for 24 h. After incubation, worms were washed with M9, resuspended in 100 mL M9 and 100 mL of 2× MRWB, and incubated on a rocking shaker at room temperature (RT) for 1 h. Next, the samples were washed with M9 and incubated in 1 mL of 60% isopropanol at RT for 15 min for dehydration. Subsequently, 1 mL of Oil Red O solution was added, and the nematodes were incubated on a rocking shaker at RT for 24 h. An Oil Red O solution was prepared by dissolving 125 mg Oil Red O in 41.75 mL of 60% isopropanol. Stained nematodes were placed on 2% agarose pad mounted on a glass slide and imaged using a stereomicroscope equipped with a camera (WRAYCAM-EL310; WRAYMER, Osaka, Japan). The stained regions were quantified using ImageJ software (version 1.54f). To enhance the contrast, they were inverted and split into RGB channels. The blue channel was used to set a consistent threshold for all areas based on the pixel count.
** Cholesterol degradation assay **
Cholesterol degradation was analyzed following previously described protocol (Tomaro-Duchesneau et al., 2014). Briefly, 10 mg bacterial cells and 100 mL cholesterol (10 mg mL⁻ ^1^ in ethanol) were added to 5 mL of M9 buffer and incubated with shaking at 37°C for 24 h. After incubation, 10 mL ethanol and 5 mL 33% KOH were added, vortexed, cooled to RT, and extracted with 30 mL hexane. The upper layer was collected, evaporated to dryness, and dissolved in 1 mL of ethanol. Cholesterol concentration was measured using LabAssay ^TM^ Cholesterol Kit (FUJIFILM Wako Pure Chemical Corporation, Osaka, Japan). A standard curve was generated with kit standards, and cholesterol levels in samples were calculated accordingly.
** Triglyceride degradation assay **
Triglyceride degradation was assessed using a modified method from Zhao et al. (2022). Briefly, 10 mg of bacterial pellets were incubated with 50 µL tributyrin (10 mg mL⁻¹) in 5 mL M9 buffer at 37°C with shaking for 24 h. Subsequently, 200 µL culture was mixed with 10 mL ethanol, vortexed 30 s, and centrifuged (4,000 × g, 5 min). One milliliter of supernatant was collected, and triglyceride concentration was determined using the LabAssay ^TM^ Triglyceride Kit (FUJIFILM Wako) with a standard curve from kit standards.
** Lipase inhibition assay **
Lipase inhibition was tested using a modified protocol from Núñez et al. (2023). Porcine pancreatic lipase (2.5 mg mL ^-1^ ) was prepared in Tris-base buffer (0.1 M Tris-HCl, pH7.0, 5 mM CaCl 2 ). After centrifugation (2,000 ×g , 3 min) 40 µL bacterial suspension or orlistat control (100 mg mL⁻ ^1^ ) was mixed with 40 µL enzyme solution on ice. Substrate solution (20 mL of 10 mM p-Nitrophenyl butyrate in Tris-base buffer) was added and absorbance (Abs) at 400 nm measured every 5 min at 37°C. Reactions without substrate served as blanks. Lipase inhibition was calculated:
Lipase inhibition (%) = [(Abs of lipase only - Abs of sample with added bacteria) / Abs of lipase only] ×100
** RNA purification and RT-qPCR analyses **
One hundred nematodes; fed bacteria for 1 day at 20°C were collected in lysis solution [0.5% Triton X-100, 0.5% Tween 20, 0.25 mM EDTA, 2.5 mM Tris-HCl, pH 8.0, 8% RNAsecure™ RNase Inactivation Reagent (Thermo Fisher Scientific Inc., WA, USA), and 1 mg mL ^−1^ proteinase K] and flash-frozen in liquid nitrogen (Jiang et al., 2019). Worm lysates were prepared by heating at 65°C for 15 min, followed by heating at 85°C for 1 min. To these lysates, 100% ethanol (2.5 times the volume of the sample) and 3 M sodium acetate (10% volume of the sample) were added, and stored overnight at −30°C. After centrifugation (14,000 ×g , 20 min), RNA was purified, and cDNA synthesized using PrimeScript™ RT reagent Kit with gDNA Eraser (Takara Bio Inc., Kusatsu, Japan). RT-qPCR was performed with TB Green® Premx Ex Taq™ II (TaKaRa Bio) on the Quantstudio 1 real-time PCR System (Thermo Fisher Scientific Inc., WA, USA). Primers are listed in the Reagents section. Relative expression was calculated by the 2 ^-ΔΔCt^ method, normalized to four housekeeping genes ( * tba-1 , cdc-42 , eif-3 .C, pmp-3 * ) (Taki and Zhang, 2013).
** Statistical analysis **
Statistical analyses conducted in R software (version 4.4.0). Statistical significance was set at p < 0.05. Values are annotated as ns (not significant), * p < 0.05, ** p < 0.01, or *** p < 0.001.
Reagents
Primers of each gene using RT-qPCR. Forward and reverse primers are denoted as For or Rev, respectively.
tba-1 *
For: CGTACACTCCACTGATCTCTG, Rev: CTTTGGAACGACATCTCCTCTG
cdc-42 *
For: CTATCGTATCCACAGACCGAC, Rev: CTGGATCATCCCTGAGATCG
eif-3 .C *
For: TGCCACTGTTTCGCTCAAGAAG, Rev: GTGCATGATGAGACAGTCGGTG
pmp-3 *
For: CTACACTTTCACCGCCCAATGAC, Rev: TCCACATCCACTGTCTCCAGTTATC
pod-2 *
For: ATTGAGCAGTATTACGAGACGCAG, Rev: CATAAACGTGTTGAGCTGCTGTTC
fasn-1 *
For: GAACATCAATGTCTTCTCTGCCACC, Rev: TCCGTAGTAGAATGGTTGGTTGTC
elo-2 *
For: CATGGGCTCGTTGGTCTTTGG, Rev: GGGCGTGGAGTTTGGATGTTC
fat-3 *
For: CCACACGCAACATGACTCCATC, Rev: AGTAGTCATCGACGAGGTAAGGAAG
fat-5 *
For: GATGGGTATTCCTCCTGCACAC, Rev: CCATGAGAGGGTGGCTTTGTAG
fat-7 *
For: TGTTTCACACTTCACGCCACTTG, Rev: TGGGAATGTGTGGTGGAAATTGTG
lbp-1 *
For: GACAGCACCAAACACGAGATCAC, Rev: AGGTACTCGTAGGTCTCTTCCTTG
lbp-8 *
For: GGTGACACTTGGCATTTCAATC, Rev: AACCAAAGTGTTGTAGGATCGCTC
lipl-1 *
For: GTTGAAGCTGGAGTTTGTGACGATG, Rev: GAGTAGAAGTTCCTGCGGGTGTATG
lipl-3 *
For: TGGACTATGAACCTGCCCGAG, Rev: TGTGCTTCATGCTGTAAGTGTTCC
atgl-1 *
For: TGAAGAAACGTGCGAGTGCAAATG, Rev: GGAATATCACTGTCACCACACGTC
acs-2 *
For: CCAAAGAGACACAAGCACAGCAAC, Rev: GCTCTGGGATGTCATGGGATTTG
acs-5 *
For: CCGGATTTGAACTACTTTGCGAAGG, Rev: CTTCATCAAACAACTCGGCGGTTC
acs-22 *
For: GCCTGTGATGGATGTGGAAGATG, Rev: CCTTGACGACAATACCAGCCATTC
acox-1.4 *
For: TGATACGGATAAGATGGCGGCTG, Rev: GGTCCATGAACGGAAAGGGTTG
ech-9 *
For: GTGGAACTATGGGAAAGGCAATGG, Rev: TCCCGAGCATACATAACTTCCAAC
ech-1.1 *
For: TGCATCCAATACTTCTGCTCTTCC, Rev: TAGTCCTAGCTGAGCAGCAGTTG
acdh-1 *
For: GAATCCGAGCTTCATCCACTTGTC, Rev: CTGAGCCAGTCCAATCATTTGTGC
sbp-1 *
For: GATTGGATTGCTCGCTGGAAGTG, Rev: TGCTAGTTCCATCCGGGTCATC
hlh-30 *
For: GAGTCGCCACCAAATCACACG, Rev: GTTCGGCGATGTGATCTGCATG
daf-12 *
For: TGTTGGCTGTTCTCTTCTCTGTCC, Rev: CTGCTCTCCGAACAACGATTCC
nhr-49 *
For: AGATGACGCACCCACAAGATATG, Rev: AACTCGGAGAGCAGAGAATCCAC
nhr-76 *
For: TCGACGATATTCTGTTCTCTGCTGC, Rev: GTATAGTGCGGCAAGGGTTTCG
nhr-80 *
For: TGAGGAATTTGTGGCGCTGAAG, Rev: GTTGGGTGATTAGGTGACGTTTCG
ser-6 *
For: CTGTCCGTCAGCCAATCAAATACC, Rev: GAGTATGACAGCCAGAAGTAGGGAG
mod-1 *
For: GTGAAAGCAATGGATGTGTGGATG, Rev: CGCGTTTCTTACGCTGTTCTGAC
daf-15 *
For: CAGGAAGTAAACAACAGACAGGACC, Rev: TCGACTCGACCATCAGCATAAC
let-363 *
For: AGCTGTAAGAATCGCCGGAATTG, Rev: GTTTGGTTGACCTTCCAGTGCTTC
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Aarnio Vuokko Lehtonen Marko Storvik Markus Callaway James C. Lakso Merja Wong Garry 2011 Caenorhabditis Elegans Mutants Predict Regulation of Fatty Acids and Endocannabinoids by the CYP-35A Gene Family Frontiers in Pharmacology 21663-981210.3389/fphar.2011.00012 PMC 310855821687507 · doi ↗ · pubmed ↗
- 2Apovian CM 201661 Obesity: definition, comorbidities, causes, and burden.Am J Manag Care 227 Suppl 1088-0224 s 176s 18527356115 · pubmed ↗
- 3Ashrafi Kaveh 2007 Obesity and the regulation of fat metabolism Worm Book 1551-850710.1895/wormbook.1.130.1PMC 478088018050496 · doi ↗ · pubmed ↗
- 4Chen Chien-Chang Kong Man-Shan Lai Ming-Wei Chao Hsun-Chin Chang Kuei-Wen Chen Shih-Yen Huang Yhu-Chering Chiu Cheng-Hsun Li Wen-Chen Lin Pen-Yi Chen Chih-Jung Lin Tzou-Yien 201021 Probiotics Have Clinical, Microbiologic, and Immunologic Efficacy in Acute Infectious Diarrhea Pediatric Infectious Disease Journal 2920891-366813513810.1097/inf.0b 013e 3181 b 530bf 20135748 · doi ↗ · pubmed ↗
- 5Du Mez-Kornegay Rachel N. Baker Lillian S. Morris Alexis J. De Loach Whitney L. M. Dowen Robert H. 2024328 Kombucha Tea-associated microbes remodel host metabolic pathways to suppress lipid accumulation PLOS Genetics 2031553-7404 e 1011003 e 101100310.1371/journal.pgen.101100338547054 PMC 10977768 · doi ↗ · pubmed ↗
- 6Honda Shotaro Eguchi Hiromichi Okino Yoichi Wang Dian-Sheng 2025324 The Probiotic Strain Clostridium butyricum TO-A Produces Butyrate by Utilizing Lactate and Acetate International Journal of Molecular Sciences 2671422-00672951295110.3390/ijms 2607295140243571 PMC 11988312 · doi ↗ · pubmed ↗
- 7Huang Yung-Feng Liu Po-Yen Chen Ying-Yao Nong Bao-Ren Huang I-Fei Hsieh Kai-Sheng Chen Kow-Tong 201411 Three-Combination Probiotics Therapy in Children With Salmonella and Rotavirus Gastroenteritis Journal of Clinical Gastroenterology 4810192-0790374210.1097/mcg.0b 013e 31828 f 1c 6e 23632352 · doi ↗ · pubmed ↗
- 8Inatomi Takio Amatatsu Masaaki Romero-Pérez Gustavo A. Inoue Ryo Tsukahara Takamitsu 20171222 Dietary Probiotic Compound Improves Reproductive Performance of Porcine Epidemic Diarrhea Virus-Infected Sows Reared in a Japanese Commercial Swine Farm under Vaccine Control Condition Frontiers in Immunology 81664-322410.3389/fimmu.2017.01877 PMC 574391529312349 · doi ↗ · pubmed ↗