The Complete Mitochondrial Genome of Nearly Threatened Kwang‐Yang Asian Frog Nanorana quadranus (Anura: Dicroglossidae) and Its Phylogenetic Analyses
Jia Liu, Bin Zuo, Yan‐Bo Sun

TL;DR
This paper reports the first complete mitochondrial genome of a near-threatened frog species from the Qinghai-Tibet Plateau, providing genetic data for its conservation and evolutionary studies.
Contribution
The first complete mitogenome of Nanorana quadranus is sequenced and analyzed for phylogenetic placement.
Findings
The mitogenome is 20,173 bp long and includes 13 PCGs, 21 tRNAs, 2 rRNAs, and a control region.
Phylogenetic analysis shows N. quadranus clusters with N. taihangnica within the Dicroglossidae family.
The study provides foundational genetic data for future conservation and evolutionary research on this species.
Abstract
Nanorana quadranus (Liu, Hu & Yang, 1960) is a high‐altitude amphibian species native to the Qinghai‐Tibet Plateau. It plays a vital role in alpine wetland ecosystems and is listed as near threatened (NT) in the IUCN Red List. Here, we present the first complete mitogenome of N. quadranus using the Illumina NovaSeq 6000 sequencing platform. The total length of the mitogenome is 20,173 bp (Accession No. PV546862). It encompasses 13 protein‐coding genes (PCGs), 21 transfer RNAs (tRNAs), 2 ribosomal RNAs (rRNAs), and one control region. Phylogenetic analysis revealed that N. quadranus forms a clade with N. taihangnica , supporting its taxonomic placement within the Dicroglossidae family. This study provides foundational genetic data for further research on the evolution and conservation of this species. We report the first complete mitochondrial genome of the near‐threatened frog…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 FIGURE 1
FIGURE 1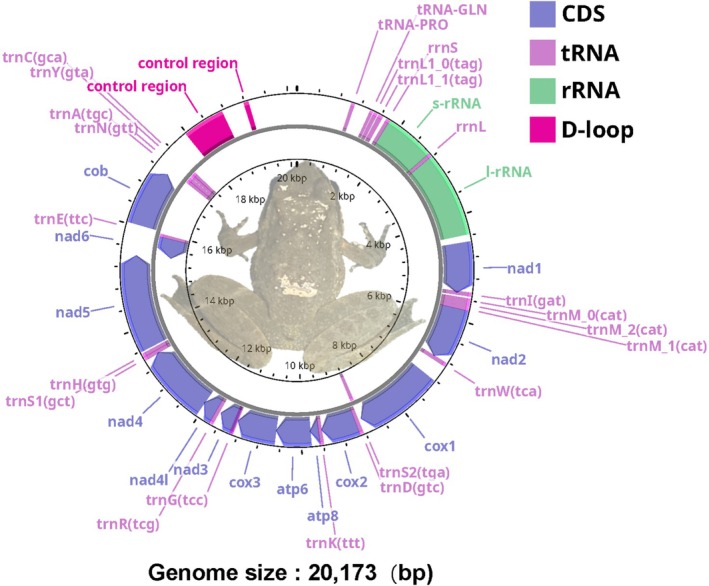 FIGURE 2
FIGURE 2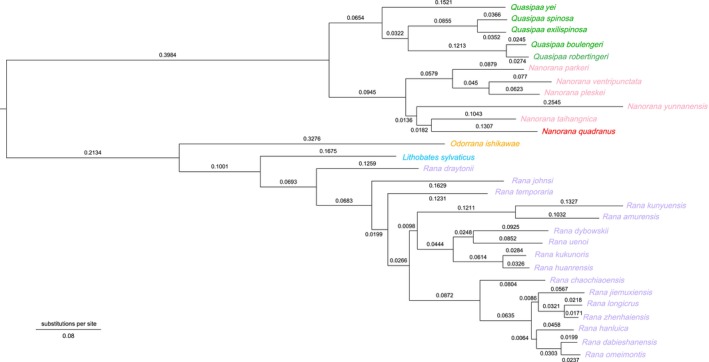 FIGURE 3
FIGURE 3| Family | Genus | Species | Accession number | References |
|---|---|---|---|---|
| Dicroglossidae |
|
| (Jiang et al. | |
|
| (Chen et al. | |||
|
| This study | |||
|
| (Chen et al. | |||
|
|
|
Unpublished (Zhang et al. | ||
|
|
| (Shan et al. | ||
|
| (Wu et al. | |||
|
| Unpublished | |||
|
| (Zhou et al. | |||
|
| (Chen et al. | |||
| Ranidae |
|
| Unpublished | |
|
| Unpublished | |||
|
| Unpublished | |||
|
| Unpublished | |||
|
| (Li, Lei, and Fu | |||
|
| Unpublished | |||
|
| (Dong et al. | |||
|
| Unpublished | |||
|
| (Chen et al. | |||
|
| (Wang et al. | |||
|
| (Li, Yin, et al. | |||
|
| Unpublished | |||
|
| Unpublished | |||
|
| (J. J. Chen | |||
|
| Unpublished | |||
|
| (Huang et al. | |||
|
| (Kurabayashi et al. | |||
|
| (Ni et al. |
- —Yunnan Fundamental Research Projects
- —National Key Research Development Program of China
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGenomics and Phylogenetic Studies · Amphibian and Reptile Biology · Ichthyology and Marine Biology
Introduction
1
The genus Nanorana (Anura: Dicroglossidae) comprises small to medium‐sized frogs adapted to harsh high‐altitude environments. Nanorana quadranus , endemic to the eastern Qinghai‐Tibet Plateau, is a keystone species in alpine wetlands (Liu et al. 1960). In living specimens, distinct yellow lateral stripes are present along the body flanks; dorsal tubercles are relatively sparse and irregularly distributed (Figure 1). Due to anthropogenic landscape modification and ongoing climatic shifts, the species has been classified as Near Threatened on the IUCN Red List of Threatened Species (IUCN 2020). Although previous studies focused on its reproductive biology (Xiao et al. 2024) and taxonomic disputes with Quasipaa species (Chen et al. 2017), the absence of mitogenomic data hindered resolution of its evolutionary history and population genetic analysis. This study bridges this gap by presenting the first complete mitogenome of N. quadranus , enabling insights into its phylogenetic position.
Morphological photograph of Nanorana quadranus (photographed by Bin Zuo at Mianyang City, Sichuan Province, China).
Materials and Methods
2
Ethics Statement
2.1
All animal procedures were approved by the Yunnan University Ethics Committee (Approval No. YNU20230645) and complied with the National Wildlife Protection Law of China.
Sample Collection and Sequencing
2.2
An adult male N. quadranus specimen was collected from Qianfo Town, Mianyang City, Sichuan Province, China (latitude: 31.710, longitude: 104.262). Species identification was first conducted based on morphological characteristics following Liu et al. (1960) and regional taxonomic keys (Liu et al. 1960). In addition, the presence of a swollen vent (cloacal swelling) was compared with descriptions in the AmphibiaChina database (http://www.amphibiachina.org/species/445) to further support the taxonomic assignment (Che and Wang 2016). The specimen voucher, designated as CB237‐001R0002, is securely stored in the Ministry of Education Key Laboratory for Transboundary Ecosecurity of Southwest China at Yunnan University (Yanbo Sun, [email protected]).
Genomic DNA was extracted from the muscle tissue. Library construction was performed using the VAHTS Universal Plus DNA Library Prep Kit for Illumina (ND617). The required concentration was ≥ 1 ng/μL, with the fragment size centered around 430–530 bp and an average fragment size between 420 and 580 bp. After passing the quality check, the library was sequenced on the Illumina NovaSeq 6000 platform with a PE150 sequencing strategy.
De Novo Assembly and Annotation of Mitochondrial Genomes
2.3
For the raw sequencing data, we performed quality control using fastp (v0.21.0) with parameters: ‐q 10 ‐u 50 ‐y ‐g ‐Y 10 ‐e 20 ‐l 100 (S. F. Chen 2023). The mitochondrial genome sequence of N. taihangnica was used as a reference (Chen et al. 2014). The mitochondrial genome of N. quadranus was assembled using MITObim (v1.9.1) with parameters: ‐start 1 ‐end 10 ‐readpool ‐quick ‐kbait 21 (Hahn et al. 2013). After assembly, the mitochondrial genome was annotated using the MITOS (Bernt et al. 2013) and MitoFinder v1.4.2 (Allio et al. 2020). We manually merged the annotation results from both tools and performed visualization using Proksee (Grant et al. 2023). We further analyzed the tRNAs of the mitochondrial genomes of N. quadranus and N. taihangnica using tRNAscan‐SE (v.2.0.12) (Chan et al. 2021) with the parameters ‐M vert ‐thread 20. To validate the assembly reliability of the D‐loop region, we mapped the raw sequencing reads to the mitochondrial sequence using BWA v0.7.17 (Li and Durbin 2009), calculated per‐base coverage, and determined the coverage of both the D‐loop and non‐D‐loop regions.
Phylogenetic Analysis
2.4
Phylogenetic relationships were inferred using 13 PCGs from 29 amphibian mitogenomes (including N. quadranus ; Table 1). The PCGs sequences were retrieved and concatenated with FasParser software (Sun 2018). Multiple sequence alignment of each PCG was conducted in Muscle v5 (Edgar 2022). Finally, a maximum‐likelihood tree was constructed using IQ‐TREE v2.4 (‐alrt 2000 ‐bb 2000 ‐nt 40 ‐bnni) (Minh et al. 2020). To further support the taxonomic identification of our specimens as N. quadranus , we downloaded cox1 (partial cds) sequences of N. quadranus (NCBI accession number: PP572988.1) and N. taihangnica (NCBI accession number: PP572987.1) from NCBI. These sequences were aligned with the full‐length cox1 gene extracted from our newly assembled mitochondrial genome, and a phylogenetic tree was constructed using the same methods described above. In addition, pairwise genetic distances of cox1 were calculated using the K80 (Kimura 2‐parameter) model.
Results
3
Taxonomic Identification
3.1
The phylogenetic analysis further supports that the specimens in this study belong to N. quadranus , as the cox1 gene from our newly assembled mitochondrial genome clustered closely with published N. quadranus sequences, while N. taihangnica was positioned as an outgroup (Figure S1).
Mitogenome Organization
3.2
The mitochondrial genome of Nanorana quadranus showed exceptionally high sequencing quality. The coverage across the mitochondrial genome was 99.13%, with an average sequencing depth of approximately 10,310×. The mean base quality score was 37.7, and the mean mapping quality score was 55.7, indicating highly accurate base calls and reliable read alignments. The total length of the N. quadranus mitogenome is 20,173 bp (GenBank Accession No. PV546862), with a GC content of 41.13%, comprising 13 PCGs, 21 tRNAs (with the Threonine tRNA missing), 2 rRNAs, and one control region (Figure 2). Although the D‐loop assembly is represented by two separate intervals due to repetitive sequences, it corresponds to a single biological control region. Due to the high variability of the D‐loop region, mitochondrial genome annotation often splits a single D‐loop region into multiple segments. For example, in Nanorana yunnanensis (KF199150.2), three separate D‐loop regions were annotated, illustrating that the high variability of the D‐loop region poses challenges for accurate annotation. The D‐loop region shows a mean coverage of 44,692×, while the non‐D‐loop regions have a mean coverage of 8813×. Reads continuously span the entire D‐loop, supporting that the ~20 kb mitochondrial genome represents genuine biological sequence rather than an assembly artifact. Sequencing depth analysis indicates that, despite variations at the boundaries, these two annotated D‐loop regions maintain stable coverage (Figure S2). Gene order and orientation are conserved relative to other anurans (Chen et al. 2014). Twelve PCGs and 15 tRNAs are encoded on the heavy strand, while ND6 and 6 tRNAs reside on the light strand (Figure 2).
Mitochondrial genome map of the Nanorana quadranus . Different gene functions are represented by colored outside circles, and transcriptional direction is represented by arrows.
Phylogenetic Analysis
3.3
The phylogenetic tree (Figure 3) robustly supports the monophyly of Dicroglossidae (Bootstrap support [BS] = 100%). N. quadranus clusters with N. taihangnica (BS = 100%), aligning with previous classifications (Chen et al. 2017).
Maximum‐likelihood analysis of N. quadranus and 29 related species based on 13 PCG sequences with 1000 bootstraps. Branch length values (substitutions per site) are labeled near the branch. The species analyzed in this study are represented by red fonts. The sequences from the following species were used: Nanorana parkeri NC_026789 (Jiang et al. 2016), Nanorana pleskei NC_016119 (Chen et al. 2011), Nanorana taihangnica NC_024272 (Chen et al. 2014), Nanorana ventripunctata NC_039094, Quasipaa boulengeri NC_021937 (Shan et al. 2014), Quasipaa exilispinosa NC_056269 (Wu et al. 2020), Quasipaa robertingeri KY441640, Quasipaa spinosa NC_013270 (Zhou et al. 2009), Quasipaa yei NC_024843 (Chen et al. 2015), Rana amurensis NC_030042, Rana chaochiaoensis NC_035803, Rana dabieshanensis NC_060306, Rana draytonii NC_028296 (Li, Lei, and Fu 2016), Rana dybowskii NC_023528, Rana hanluica NC_061371, Rana huanrensis NC_028521 (Dong et al. 2016), Odorrana ishikawae AB511282 (Kurabayashi et al. 2010), Rana jiemuxiensis PP797569, Rana johnsi NC_058599 (Chen et al. 2021), Rana kukunoris MN733918 (Wang et al. 2020), Rana kunyuensis NC_024548 (Li, Yin, et al. 2016), Rana longicrus NC_061370, Rana omeimontis NC_035805, Lithobates sylvaticus NC_027236 (Ni et al. 2016), Rana temporaria NC_042226 (J. J. Chen 2018), Rana uenoi NC_056272, Rana zhenhaiensis MN218687 (Huang et al. 2019), Nanorana yunnanensis KF199150.2 (Zhang et al. 2018).
Discussion and Conclusion
4
The assembly of the first complete mitogenome of N. quadranus provides critical insights into the evolutionary relationships of this high‐altitude endemic species and lays a foundation for future studies on high‐altitude adaptations, particularly involving mitochondrial genes related to energy metabolism. Notably, the close phylogenetic clustering of N. quadranus with N. taihangnica (BS = 100%) robustly supports its taxonomic placement within the Dicroglossidae family. Additionally, integrating nuclear genomic data, such as single‐copy orthologs or sex‐linked markers (Xiao et al. 2024), would refine phylogenetic hypotheses and elucidate whether adaptive traits (e.g., cold tolerance) originated via mito‐nuclear coevolution or independent nuclear mutations.
Given the close phylogenetic relationship between N. quadranus and N. taihangnica , we conducted additional analyses to clarify their taxonomic distinction. Specifically, we reconstructed a phylogenetic tree using the published cox1 sequences of both species together with the cox1 sequence generated in this study. The results robustly placed our specimen within N. quadranus and clearly differentiated N. quadranus from N. taihangnica . Furthermore, diagnostic morphological characters also support this conclusion: N. quadranus exhibits a square‐shaped cutaneous swelling surrounding the cloaca, a feature absent in N. taihangnica . This morphological distinction is consistent with the species description provided in the AmphibiaChina database (Che and Wang 2016), thereby reinforcing the recognition of N. quadranus as a species distinct from N. taihangnica . Our analysis shows that the new * N. quadranus cox1* differs from the published partial fragment by ~5%, while both differ from * N. taihangnica cox1* by ~18%, supporting them as distinct species.
Based on the phylogenetic analysis, N. yunnanensis clustered with N. quadranus and N. taihangnica , which further supports the previously proposed view that the genus Feirana is invalid (Zhang et al. 2018).
Regarding the loss of tRNA‐thr in our assembly, we carefully examined the annotation and compared it with closely related species and found that the loss of tRNA‐thr is not due to a software error but represents a branch‐specific gene loss. In the mitochondrial genome of N. taihangnica , a close relative of N. quadranus , tRNA‐thr was also not identified (Figure S3), suggesting an ancestral loss resulting from a certain evolutionary event. Combined with tRNAscan‐SE analysis of the complete mitochondrial genomes of both species (Tables S1 and S2), the results further support the conclusion of a lineage‐specific loss rather than an artifact of assembly or software error. Although the loss or duplication of tRNAs is relatively rare in vertebrates, previous studies have documented tRNA gene loss and rearrangements in the mitochondrial genomes of vertebrates such as Gekkonidae (Kumazawa et al. 2014), representing a branch‐specific evolutionary event.
Author Contributions
Jia Liu: data curation (equal), investigation (equal), methodology (equal), software (equal), writing – original draft (equal). Bin Zuo: data curation (equal), investigation (equal), methodology (equal), visualization (equal), writing – original draft (equal). Yan‐Bo Sun: conceptualization (lead), funding acquisition (lead), project administration (lead).
Funding
This work was supported by the National Key Research Development Program of China (2022YFF0802300) and the Yunnan Fundamental Research Projects (202401BC070011).
Disclosure
The authors have nothing to report.
Ethics Statement
This study involved research on Nanorana quadranus , a Near Threatened species classified by the International Union for Conservation of Nature (IUCN). All procedures strictly adhered to the IUCN Guidelines for Appropriate Uses of Red List Data. We further complied with the principles of the Convention on Biological Diversity (CBD) regarding access to genetic resources and benefit‐sharing, and confirmed that no activities related to international trade of specimens (as regulated by CITES) were conducted. Ethical approval for this research was granted by the Yunnan University Ethics Committee (YNU20230645), and all data collection protocols were designed to minimize disturbance to wild populations.
Conflicts of Interest
The authors declare no conflicts of interest.
Supporting information
Figure S1: The phylogenetic analysis further supports that the specimens in this study belong to N. quadranus , as the cox1 gene from our newly assembled mitochondrial genome clustered closely with published N. quadranus sequences, while N. taihangnica was positioned as an outgroup. Figure S2:. Analysis of sequencing depth in the D‐loop regions. The x‐axis represents the nucleotide positions within the D‐loop regions, and the y‐axis shows the sequencing depth on a log10 scale. Figure S3:. Complete mitochondrial genome of N. taihangnica (NCBI Reference Sequence: NC_024272.1) and its visualization. The result shows that the mitochondrial genome of N. taihangnica lacks the tRNA‐Thr. Table S1:. Results of tRNAscan‐SE analysis of the mitochondrial genome of N. quadranus . Table S2:. Results of tRNAscan‐SE analysis of the mitochondrial genome of N. taihangnica .
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Allio, R. , A. Schomaker‐Bastos , J. Romiguier , F. Prosdocimi , B. Nabholz , and F. Delsuc . 2020. “Mito Finder: Efficient Automated Large‐Scale Extraction of Mitogenomic Data in Target Enrichment Phylogenomics.” Molecular Ecology Resources 20: 892–905.32243090 10.1111/1755-0998.13160 PMC 7497042 · doi ↗ · pubmed ↗
- 2Bernt, M. , A. Donath , F. Jühling , et al. 2013. “MITOS: Improved Metazoan Mitochondrial Genome Annotation.” Molecular Phylogenetics and Evolution 69: 313–319.22982435 10.1016/j.ympev.2012.08.023 · doi ↗ · pubmed ↗
- 3Chan, P. P. , B. Y. Lin , A. J. Mak , and T. M. Lowe . 2021. “t RN Ascan‐SE 2.0: Improved Detection and Functional Classification of Transfer RNA Genes.” Nucleic Acids Research 49: 9077–9096.34417604 10.1093/nar/gkab 688PMC 8450103 · doi ↗ · pubmed ↗
- 4Che, J. , and K. Wang . 2016. “Amphibia China: An Online Database of Chinese Amphibians.” Dongwuxue Yanjiu 37: 57–59.26828034 10.13918/j.issn.2095-8137.2016.1.57PMC 4834736 · doi ↗ · pubmed ↗
- 5Chen, G. Y. , B. Wang , J. Y. Liu , F. Xie , and J. P. Jiang . 2011. “Complete Mitochondrial Genome of Nanorana pleskei (Amphibia: Anura: Dicroglossidae) and Evolutionary Characteristics.” Current Zoology 57: 785–805.
- 6Chen, J. J. 2018. “The Complete Mitochondrial Genome of Common Terrestrial Frog (Rana temporaria).” Mitochondrial DNA Part B Resources 3: 980–981.33490552 10.1080/23802359.2018.1507649 PMC 7800347 · doi ↗ · pubmed ↗
- 7Chen, Q. P. , L. Wu , S. S. Zhang , et al. 2021. “The Complete Mitochondrial Genome of Rana johnsi (Anura: Ranidae) and Its Phylogeny.” Mitochondrial DNA Part B Resources 6: 3493–3495.34869888 10.1080/23802359.2021.2002213 PMC 8635646 · doi ↗ · pubmed ↗
- 8Chen, S. F. 2023. “Ultrafast One‐Pass FASTQ Data Preprocessing, Quality Control, and Deduplication Using Fastp.” i Meta 2: e 107.38868435 10.1002/imt 2.107PMC 10989850 · doi ↗ · pubmed ↗