Genome-Wide Identification and Analysis of bZIP Transcription Factors in Coptis chinensis Reveals Their Regulatory Roles in Stress Responses
Wuke Wei, Zijian Le, Lianan Guo, Rangyu Mo, Yu Wang, Yuan Pan

TL;DR
This study identifies and analyzes bZIP transcription factors in Coptis chinensis, revealing their roles in stress responses and medicinal compound production.
Contribution
The first genome-wide analysis of bZIP transcription factors in Coptis chinensis, revealing lineage-specific evolution and stress-related gene expression patterns.
Findings
Fifty-five CcbZIP genes were identified and classified into 12 subfamilies, with subfamilies L and O absent.
Expression of CcbZIP genes was found to respond dynamically to temperature and light stresses in a tissue-specific manner.
The study provides insights into the regulatory roles of CcbZIPs in stress adaptation and secondary metabolism.
Abstract
The basic leucine zipper (bZIP) transcription factors play crucial roles in plant growth and stress adaptation. However, a comprehensive genome-wide analysis of this family has been lacking in the medicinal plant Coptis chinensis. In this study, we identified 55 bZIP genes (CcbZIPs) from the C. chinensis genome and systematically classified them into 12 subfamilies through phylogenetic analysis with Arabidopsis thaliana. Notably, subfamilies L and O were absent, while two orphan genes were discovered, indicating lineage-specific evolution. Expression profiling revealed that numerous CcbZIPs respond dynamically to temperature and light stresses in a tissue-specific manner. These findings provide a foundation for understanding the regulatory roles of CcbZIPs in environmental adaptation and secondary metabolism, offering potential genetic targets for future breeding aimed at improving…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
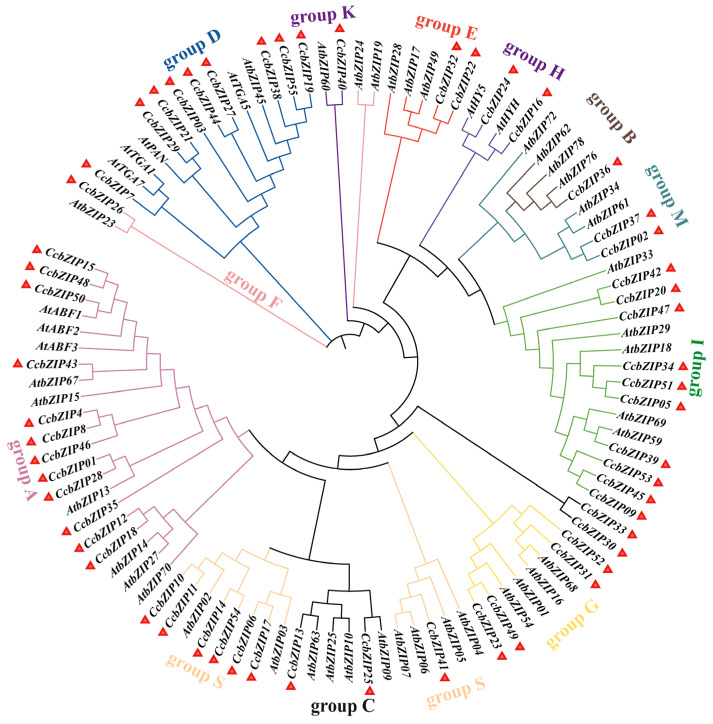 Figure 1
Figure 1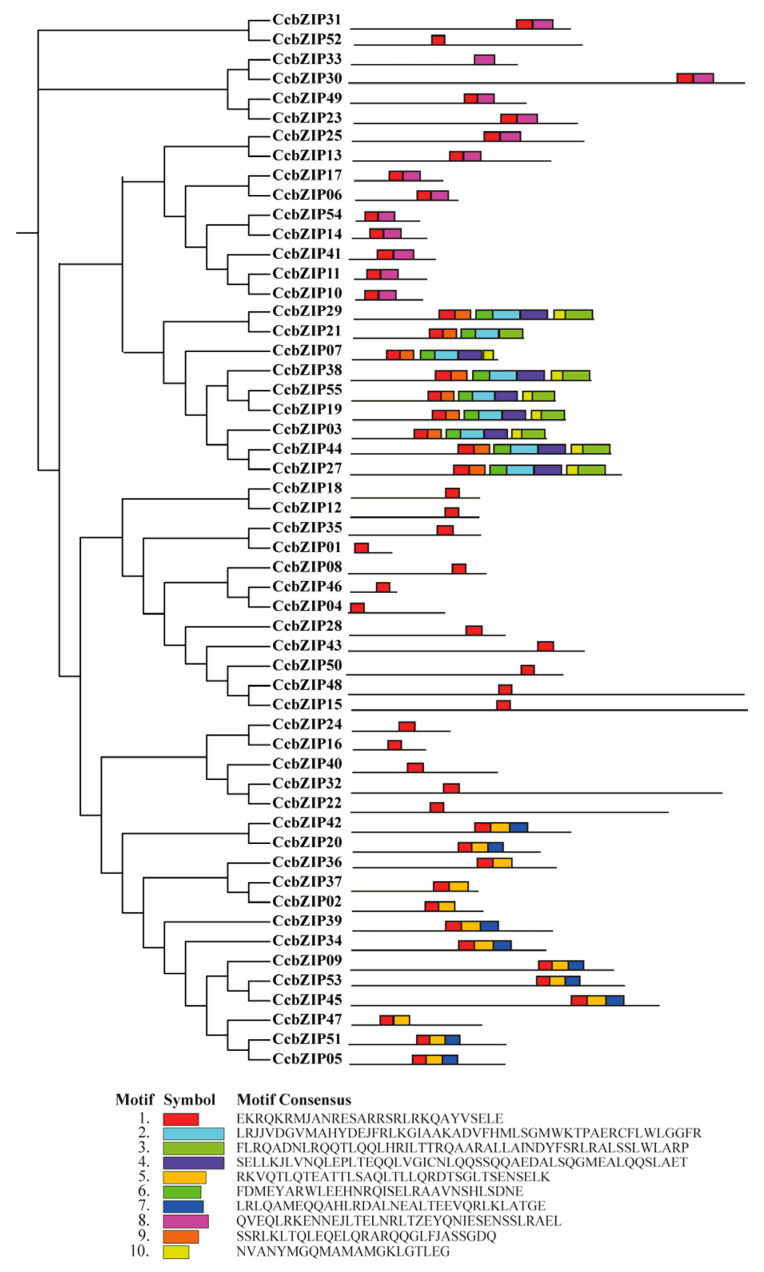 Figure 2
Figure 2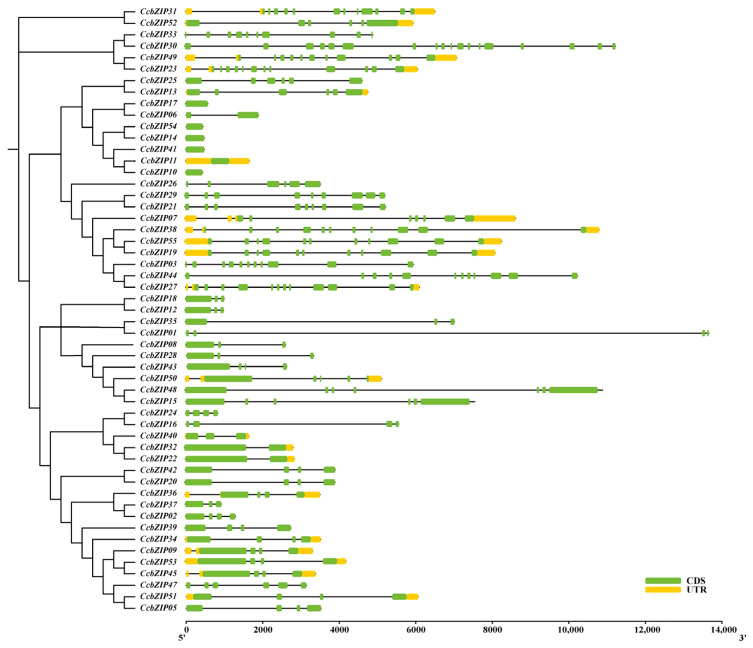 Figure 3
Figure 3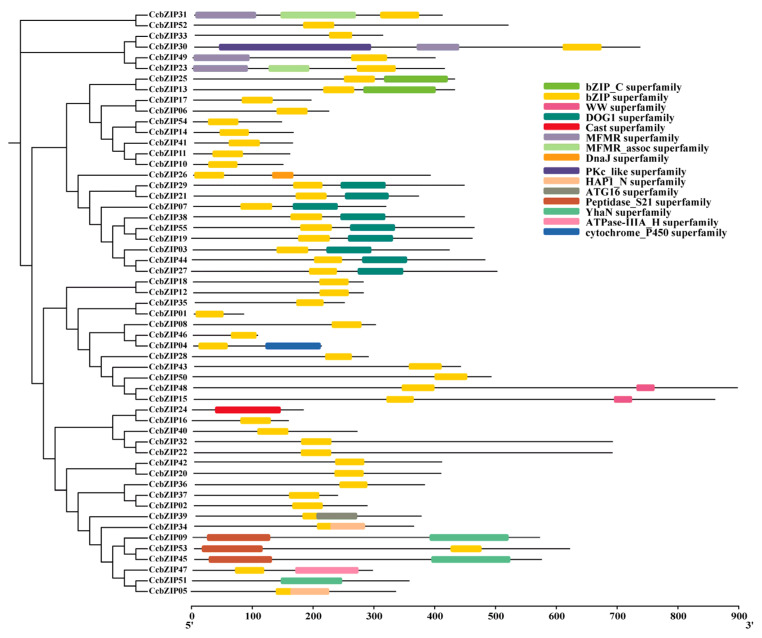 Figure 4
Figure 4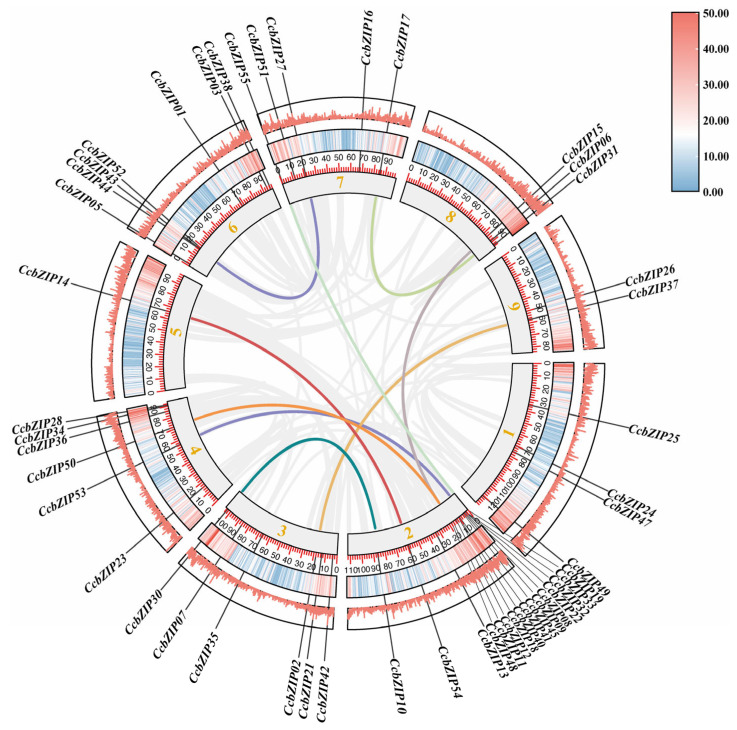 Figure 5
Figure 5 Figure 6
Figure 6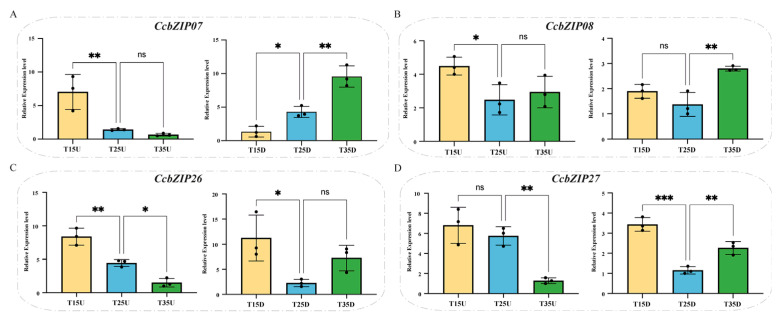 Figure 7
Figure 7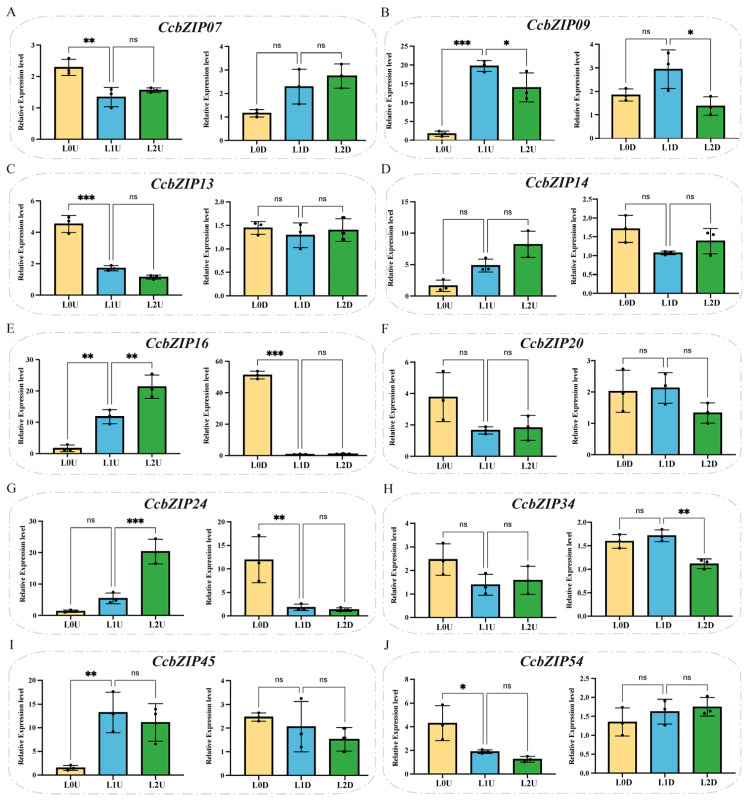 Figure 8
Figure 8- —the National Key Research and Development Program of China
- —Chongqing Special Program for Technological Innovation and Application Development
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Gene Expression Analysis · Plant Molecular Biology Research · Plant Stress Responses and Tolerance
1. Introduction
Transcription factors (TFs) are central regulators of gene expression, governing diverse processes in plant growth, development, and adaptation to environmental stresses [1,2]. Among various TF families, the basic leucine zipper (bZIP) family represents one of the largest and most evolutionarily conserved groups in eukaryotes [3]. Typical bZIP proteins are characterized by two functional domains: an N-terminal basic region that facilitates sequence-specific DNA binding, and a C-terminal leucine zipper domain responsible for dimerization [4]. This structure enables bZIP TFs to recognize a variety of DNA motifs—such as G-box (CACGTG), A-box (TACGTA), and C-box (GACGTC)—thereby regulating distinct transcriptional programs involved in numerous signaling pathways [1,5]. Functionally, bZIPs participate in a wide spectrum of physiological events, including embryogenesis, photomorphogenesis, flowering time control, and responses to biotic and abiotic stresses, underscoring their broad regulatory significance [6,7].
Genome-wide studies have systematically identified bZIP members across numerous plant species, revealing both conserved and lineage-specific evolutionary patterns [8]. In the model plant Arabidopsis thaliana, 78 bZIP genes have been classified into 13 subfamilies, with many members implicated in abscisic acid (ABA) signaling, pathogen defense, and energy homeostasis [1]. Similarly, in horticultural and medicinal plants such as litchi [9], wheat [10], rice [11], maize [12], tomato [13], potato [14], banana [15], cucumber [16], sesame [17], pomegranate [18], etc. bZIP TFs have been linked to fruit senescence, stress tolerance, and the biosynthesis of specialized metabolites like terpenoids [19]. Notably, bZIPs are increasingly recognized as key regulators of secondary metabolism in medicinal plants. For instance, AabZIP1, AabZIP9, and AaTGA6 in Artemisia annua positively regulate the biosynthesis of the antimalarial compound artemisinin [20,21,22], while bZIPs in Carthamus tinctorius modulate flavonoid biosynthesis [23]. These findings highlight the potential of engineering bZIP TFs to enhance the production of valuable phytochemicals [24].
As integrators of environmental signals, bZIP TFs also play pivotal roles in light and temperature response pathways. They participate in photomorphogenesis through HY5-mediated signaling and in temperature stress responses via both ABA-dependent and independent mechanisms [25]. Examples include MdHY5, which regulates light-induced anthocyanin biosynthesis in apple, and OsbZIP52, which enhances cold tolerance in rice [19,26]. These studies position bZIP TFs as critical hubs within stress-responsive networks, linking environmental perception to metabolic reprogramming [27].
Coptis chinensis (Chinese goldthread), a perennial herb of the Ranunculaceae family, is a renowned medicinal plant whose rhizomes accumulate bioactive isoquinoline alkaloids—such as berberine, palmatine, and coptisine—which exhibit antimicrobial, anti-inflammatory, and anticancer properties [28]. Environmental factors, particularly light and temperature, are known to influence both growth and alkaloid accumulation in medicinal plants [29,30]. However, despite its medicinal significance, the molecular mechanisms governing the biosynthesis and regulation of these valuable compounds remain poorly characterized. Moreover, how abiotic signals are perceived and transduced into transcriptional reprogramming—specifically through bZIP TFs—has not been systematically investigated in C. chinensis.
Therefore, to bridge this knowledge gap, we performed a genome-wide identification and analysis of the bZIP family in C. chinensis. This study aims to characterize the phylogenetic relationships, gene structures, conserved motifs, and chromosomal distributions of CcbZIP genes. Furthermore, we analyzed their expression patterns under different temperature and light conditions. Our findings provide a foundational resource for understanding the regulatory roles of CcbZIP TFs in the environmental adaptation and secondary metabolism of this medically important species.
2. Results
2.1. Genome-Wide Identification, Classification and Phylogenetic Analysis of CcbZIP Genes
Through a combination of BLASTP and HMMER searches against the Coptis chinensis genome, we identified 55 non-redundant proteins containing the characteristic bZIP domain. These genes were systematically renamed from CcbZIP1 to CcbZIP55 based on their ascending physical locations on the chromosomes (Table 1). The encoded proteins exhibited considerable diversity in their physicochemical properties. Their lengths ranged from 81 to 893 amino acids, corresponding to molecular weights from approximately 9.56 to 93.52 kDa. The theoretical isoelectric points (pI) varied from 4.89 to 11.44, with 21 proteins (38.2%) being acidic (pI < 7) and the remainder basic. All proteins were predicted to be hydrophilic, with grand average of hydropathy (GRAVY) values between −0.44 and 0.023. Subcellular localization predictions strongly indicated that the vast majority are nuclear proteins, consistent with their putative function as transcription factors [31].
To elucidate the evolutionary relationships within the bZIP family, a phylogenetic tree was constructed using the full-length amino acid sequences of the 55 CcbZIPs and 50 well-characterized AtbZIPs from Arabidopsis thaliana (Figure 1). The CcbZIP proteins were classified into subfamilies based on their clustering with established A. thaliana orthologs. This analysis classified the 55 members into 12 subfamilies: A, B, C, D, E, F, G, H, I, K, M, and S. Notably, subfamilies L and O, which are present in A. thaliana, were absent in C. chinensis. Subfamilies A and I were the most expanded, containing 12 and 10 members, respectively, followed by D (9), S (7), G (4), C (2), M (2), H (2), E (2), while B, F, and K each contained a single member. Two genes, CcbZIP30 and CcbZIP33, did not cluster with any defined subfamily and were designated as orphan genes, suggesting potential lineage-specific evolution [32].
2.2. Gene Structure and Conserved Domain Analysis
To gain deeper insights into the structural characteristics and potential functional diversification of the CcbZIP family, we systematically analyzed their conserved motifs, gene structures, and domain architectures. As shown in Figure 2, conserved motif analysis identified 10 distinct motifs (Motif 1–10) distributed across the CcbZIP proteins. The composition and sequential order of these motifs were highly conserved within members of the same phylogenetic subfamily but exhibited clear variation between different subfamilies. For instance, Subfamily A members commonly shared Motifs 1, 2, 3, and 4, whereas Subfamily I was characterized by a distinct set including Motifs 1, 5, and 6. This specific motif architecture strongly supports the reliability of our phylogenetic classification and suggests potential functional specialization among subfamilies. Analysis of the exon–intron structures further revealed substantial structural diversity (Figure 3). Genes clustered within the same subfamily generally possessed similar exon–intron organizations, reinforcing their evolutionary relationships. Notably, members of the expanded subfamilies A and I typically contained multiple introns, indicating structural complexity. In contrast, several genes from other subfamilies exhibited fewer introns or intron-less structures. This structural variation provides genomic evidence for the divergent evolution of the CcbZIP family.
Furthermore, examination of the conserved domain architecture confirmed the presence of the characteristic bZIP domain in all 55 CcbZIP proteins (Figure 4). Beyond this core domain, numerous members harbored additional functional domains, such as regulatory or protein–protein interaction domains, which were often specific to certain subfamilies. The diversity in domain composition underscores the functional versatility within this transcription factor family and supports the notion of subfamily-specific regulatory roles acquired during evolution. Collectively, the integrated analysis of motifs, gene structures, and domains reveals a strong correlation between phylogenetic grouping and structural features, providing a comprehensive genomic foundation for understanding the functional evolution of the bZIP family in C. chinensis.
2.3. Chromosomal Distribution, Gene Duplications and Synteny Analysis of CcbZIPs
The genomic locations of the 55 CcbZIP genes were mapped onto the nine chromosomes of C. chinensis (Figure 5). The distribution was uneven, with chromosomes 1, 2, 3, 4, 6, 7, and 8 harboring multiple genes, while chromosome 5 contained only a single gene (CcbZIP14). To understand the evolutionary mechanisms underlying the expansion of the CcbZIP family, we performed synteny analysis. Several conserved syntenic blocks were identified within the C. chinensis genome, indicative of segmental duplication events. Significant collinearity was observed between specific chromosome pairs, including Chr1-Chr4, Chr2-Chr4, Chr2-Chr7, and Chr6-Chr7 (highlighted by lines in Figure 5), suggesting these events have been crucial in the evolution of this gene family. Tandem duplication events were also detected, with relevant genes highlighted in Figure 5. These findings suggest that both segmental and tandem duplications have contributed to the expansion of the bZIP family in C. chinensis, a pattern consistent with the evolution of gene families in other plant species [33].
2.4. Expression Patterns of CcbZIP Genes Under Different Temperature and Light Conditions
To elucidate the transcriptional response of CcbZIP genes to environmental stimuli, we analyzed their expression profiles under varying temperature and light conditions using transcriptome data (Figure 6). FPKM-based heatmaps revealed that the expression of CcbZIP07, CcbZIP08, CcbZIP26, and CcbZIP27 was notably influenced by temperature, whereas CcbZIP07, CcbZIP09, CcbZIP13, CcbZIP14, CcbZIP16, CcbZIP20, CcbZIP24, CcbZIP34, CcbZIP45, and CcbZIP54 responded to different light intensities.
We further validated these patterns using qRT-PCR under temperature stress (Figure 7). CcbZIP07 expression was up-regulated in aerial tissues (U) at 15 °C and in subterranean tissues (D) at 35 °C. CcbZIP08 showed no significant changes across temperature treatments. CcbZIP26 was up-regulated throughout the plant under low temperature (15 °C), suggesting a systemic cold response. CcbZIP27 was down-regulated in U at 35 °C but up-regulated in D under both 15 °C and 35 °C, with a more pronounced induction under cold conditions.
Under light stress, tissue-specific expression patterns were also evident (Figure 8). CcbZIP07 was induced in U under L0 intensity but unaffected in D. CcbZIP09 was down-regulated in all tissues under L2. CcbZIP13 was up-regulated in U under L0, while CcbZIP14 remained unchanged. CcbZIP16 decreased in U under L0 but increased under L2; in D, it increased under L0 and was stable under L2. CcbZIP20 showed no significant response. CcbZIP24 was up-regulated in U under L2 and in D under L0. CcbZIP34 was suppressed in D under L2. CcbZIP45 decreased in U under L0, and CcbZIP54 increased in U under the same condition; neither gene showed significant changes in D. Collectively, these results demonstrate that CcbZIP genes exhibit distinct and tissue-specific expression patterns under temperature and light stresses, suggesting their potential roles in mediating environmental adaptation in Coptis chinensis.
3. Discussion
Our genome-wide analysis successfully identified and characterized 55 bZIP transcription factors (TFs) in the non-model medicinal plant Coptis chinensis. Phylogenetic classification revealed that the CcbZIP family is organized into 12 subfamilies, exhibiting notable divergence from the model plant A. thaliana. Two key observations underscore the unique evolutionary trajectory of this gene family in C. chinensis: the absence of the L and O subfamilies, and the presence of two orphan genes, CcbZIP30 and CcbZIP33, which did not cluster with any established subfamily. The absence of entire subfamilies suggests significant lineage-specific adaptation of the bZIP functional landscape in C. chinensis. In Arabidopsis and other plants, the L and O subfamilies are often implicated in specific stress responses and developmental processes. Their loss in C. chinensis may indicate a rewiring of regulatory networks, potentially driven by selective pressures to optimize the production of specialized metabolites, such as the isoquinoline alkaloid berberine. Conversely, the marked expansion of subfamilies A and I—known to be involved in stress signaling and plant development—suggests that these functions are of critical importance in C. chinensis.
A particularly intriguing finding is the identification of CcbZIP30 and CcbZIP33. As orphan genes, they may represent rapidly evolving, species-specific TFs that have acquired novel functions [34]. Given the rich and unique alkaloid profile of C. chinensis, it is tempting to speculate that these genes could be involved in the regulation of its specialized secondary metabolism, a compelling hypothesis that warrants further functional investigation.
Supporting their potential roles in environmental adaptation, our expression analysis revealed that numerous CcbZIP genes respond dynamically to temperature and light stresses in a tissue-specific manner. For instance, CcbZIP26 was systemically up-regulated under cold stress (15 °C), while CcbZIP27 exhibited contrasting regulation between aerial and subterranean tissues, suggesting complex, tissue-specific roles in temperature response. Similarly, several genes, including CcbZIP07, CcbZIP16, and CcbZIP24, showed distinct expression patterns under different light intensities. These tissue-specific expression profiles strongly imply that CcbZIP TFs are integral components of the signaling networks that mediate environmental perception and adaptive responses in C. chinensis.
In conclusion, this study provides the first systematic characterization of the bZIP family in C. chinensis. Our results not only elucidate the evolutionary dynamics of this important gene family in a medicinal plant but also pinpoint key candidate genes—particularly the expanded A and I subfamilies and the orphan genes CcbZIP30 and CcbZIP33—for future research. Functional validation of these candidates will be crucial for understanding their specific roles in C. chinensis growth, stress adaptation, and, most importantly, the regulation of its valuable medicinal compounds.
4. Materials and Methods
4.1. Plant Materials and Stress Treatments
The Coptis chinensis cv. “Wei Lian” used in this study was employed. Two-year-old seedlings were collected from the Huangshui Town C. chinensis cultivation base in Shizhu County, Chongqing Municipality [35]. The seedlings were transplanted into soil in a growth chamber under controlled conditions: temperature of 26 °C (light)/20 °C (dark), relative humidity of approximately 60%, and a photoperiod of 16 h light/8 h darkness. After three weeks of acclimatization, when the seedlings had established and showed vigorous growth, uniformly growing seedlings were selected.
After acclimatization, uniformly growing seedlings were subjected to abiotic stress treatments for 48 h. For temperature treatments, plants were transferred to chambers set at constant temperatures of 15 °C (low temperature), 25 °C (control), or 35 °C (high temperature) during both light and dark periods, while maintaining the same light intensity and photoperiod as the acclimatization stage. For light stress treatments, plants were exposed to different light intensities (measured at the canopy level) using white LED lights: 476lx (low light, L0), 2060lx (normal light, L1), or 8340lx (high light, L2), while maintaining the temperature at 25 °C and the original photoperiod. Following treatment, aerial parts (U) and subterranean parts (D) were harvested separately, immediately frozen in liquid nitrogen, and stored at −80 °C for subsequent RNA extraction. For each treatment, three biological replicates were collected, each consisting of a pool of three individual plants.
4.2. Identification and Sequence Analysis of CcbZIP Genes
The protein sequences of 50 Arabidopsis thaliana bZIP transcription factors were used as queries to perform a BLASTP (version 2.17.0+) search (E-value ≤ 1 × 10^−5^) against the C. chinensis protein database (BioProject: SAMN15658057) using TBtools software (version 2.303). Simultaneously, the Hidden Markov Model (HMM) profiles for the bZIP domain (PF00170 and PF07716) were retrieved from the PFAM database and used to search the same protein database using HMMER 3.0 (E-value ≤ 1 × 10^−5^). All candidate sequences obtained from both searches were merged, and redundant sequences were removed. The presence of the conserved bZIP domain in each candidate was further verified using the NCBI Conserved Domain Database (CDD) and SMART database. The identified non-redundant genes were systematically renamed from CcbZIP1 to CcbZIP55 based on their ascending physical positions from chromosome 1 to 9. The physicochemical properties of the deduced CcbZIP proteins, including molecular weight and theoretical isoelectric point (pI), were predicted using the ExPASy ProtParam tool (https://web.expasy.org/protparam/, accessed on 17 December 2025). Subcellular localization was predicted using CELLO v2.5 [36].
4.3. Phylogenetic, Gene Structure, Conserved Motif Analysis, Chromosomal Distribution and Synteny Analysis
Multiple sequence alignment of the full-length amino acid sequences of the 55 identified CcbZIPs and 50 AtbZIPs was performed using ClustalW (version 2.1) with default parameters (Gap opening penalty = 10, Gap extension penalty = 0.2; and the alignment results were manually checked to correct obvious misalignments). A phylogenetic tree was constructed using the Maximum Likelihood (ML) method in MEGA7.0 software with the JTT substitution model and 1000 bootstrap replicates. The amino acid sequence of all genes was list in Table S1. The CcbZIP proteins were classified into subfamilies based on the established classification of their A. thaliana orthologs. The exon–intron structures of the CcbZIP genes were visualized based on the genome annotation file using the Gene Structure Display Server (GSDS). Conserved protein motifs were identified using the online MEME suite (version 5.5.8) with the following parameters: maximum number of motifs, 10; optimum motif width, 6 to 50 amino acids [37]. The identified motifs were annotated by searching against the NCBI-CDD database and visualized using TBtools [38]. The physical positions of the CcbZIP genes on the chromosomes were mapped using TBtools software. Gene duplication events were analyzed using the Multiple Collinearity Scan Toolkit (MCScanX, version 1.0) with default parameters [39].
4.4. RNA Extraction, Transcriptome Sequencing, and qRT-PCR Analysis
Total RNA was extracted from approximately 100 mg of ground tissue using TRIzol reagent (Invitrogen, Waltham, MA, USA) according to the manufacturer’s instructions. Genomic DNA was removed by treatment with DNase I (Takara, Kyoto, Japan). RNA integrity was verified using 1.5% agarose gel electrophoresis. For transcriptome sequencing, cDNA library construction and Illumina sequencing were performed by a commercial service (e.g., Novogene/BGI). Fastp (version 0.18.0) software was used to control the quality of raw RNA-seq data and remove the adapter [40]. The resulting clean reads were mapped to the C. chinensis reference genome, and gene expression levels were calculated as Fragments Per Kilobase of transcript per Million mapped reads (FPKM) using StringTie v1.3.1 [41,42].
For qRT-PCR validation, first-strand cDNA was synthesized from 1 μg of total RNA using the PrimeScript™ RT reagent kit with gDNA Eraser (Takara, Japan). Gene-specific primers were designed using Primer-BLAST (https://www.ncbi.nlm.nih.gov/tools/primer-blast/, accessed on 17 December 2025) to span exon-exon junctions, and primer specificity was verified by both 1.5% agarose gel electrophoresis (single clear band) and melt curve analysis (single peak) (Supplementary Table S2 for all primer sequences). The C. chinensis actin gene was used as an internal control [35]. qRT-PCR was performed in a 20 μL reaction volume containing 10 μL of 2× qPCR MasterMix (Applied Biological Materials, Richmond, BC, Canada), 0.8 μL of each primer (10 μM), 2 μL of 1:10 diluted cDNA, and 6.4 μL of nuclease-free water. The reactions were run on a CFX96 Touch Real-Time PCR Detection System (Bio-Rad, Hercules, CA, USA) with the following thermal cycling protocol: initial denaturation at 95 °C for 30 s, followed by 40 cycles of 95 °C for 5 s and 60 °C for 30 s. A melt curve analysis (65 °C to 95 °C with increments of 0.5 °C) was performed to confirm amplification specificity. All reactions were performed with three biological and three technical replicates. Relative gene expression levels were calculated using the 2^−ΔΔCt^ method [43]. Statistical significance was determined by one-way ANOVA using GraphPad Prism version 10.0 (* p < 0.05; ** p < 0.005; *** p < 0.0005) [44].
5. Conclusions
This study presents the first genome-wide identification and analysis of the bZIP transcription factor family in the medicinal plant Coptis chinensis. We identified 55 CcbZIP genes and classified them into 12 subfamilies based on phylogenetic relationships with A. thaliana. The absence of the L and O subfamilies, common in model plants, and the discovery of two putative orphan genes (CcbZIP30 and CcbZIP33) highlight the unique evolutionary path of the bZIP family in this species. Furthermore, the distinct distribution of CcbZIP members, with significant expansions in stress-associated subfamilies A and I, suggests specific functional adaptations. Expression profiling confirmed that many CcbZIPs are responsive to temperature and light stresses in a tissue-specific manner. Collectively, these findings establish a crucial foundation for future functional studies aimed at elucidating the roles of CcbZIPs in the environmental adaptation and, potentially, the regulation of valuable benzylisoquinoline alkaloid biosynthesis in C. chinensis.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Dröge-Laser W. Snoek B.L. Snel B. Weiste C. The Arabidopsis b ZIP transcription factor family—An update Curr. Opin. Plant Biol.201845364910.1016/j.pbi.2018.05.00129860175 · doi ↗ · pubmed ↗
- 2Weidemüller P. Kholmatov M. Petsalaki E. Zaugg J.B. Transcription factors: Bridge between cell signaling and gene regulation Proteomics 202121232410.1002/pmic.20200003434314098 · doi ↗ · pubmed ↗
- 3Jeung D. Li X. Cho Y. New Roles of b ZIP-Containing Membrane-Bound Transcription Factors in Chromatin Tethering and Karyoptosis Int. J. Mol. Sci.2025261089610.3390/ijms 26221089641303380 PMC 12652670 · doi ↗ · pubmed ↗
- 4Liu Q. Fang H. Zhou H. Wang X. Hou Z. Identification and Characterization of b ZIP Gene Family Combined Transcriptome Analysis Revealed Their Functional Roles on Abiotic Stress and Anthocyanin Biosynthesis in Mulberry (Morus alba)Horticulturae 20251169410.3390/horticulturae 11060694 · doi ↗
- 5Pérez-Rodríguez P. Riaño-Pachón D.M. Corrêa L.G.G. Rensing S.A. Kersten B. Mueller-Roeber B. Pln TFDB: Updated content and new features of the plant transcription factor database Nucleic Acids Res.201038 D 822D 82710.1093/nar/gkp 80519858103 PMC 2808933 · doi ↗ · pubmed ↗
- 6Guo Z. Dzinyela R. Yang L. Hwarari D. b ZIP Transcription Factors: Structure, Modification, Abiotic Stress Responses and Application in Plant Improvement Plants 202413205810.3390/plants 1315205839124175 PMC 11313983 · doi ↗ · pubmed ↗
- 7Liu M. Wen Y. Sun W. Ma Z. Huang L. Wu Q. Tang Z. Bu T. Li C. Chen H. Genome-wide identification, phylogeny, evolutionary expansion and expression analyses of b ZIP transcription factor family in tartaty buckwheat BMC Genom.20192048310.1186/s 12864-019-5882-z PMC 656085831185893 · doi ↗ · pubmed ↗
- 8Li H. Li L. Shang Guan G. Jia C. Deng S. Noman M. Liu Y. Guo Y. Han L. Zhang X. Genome-wide identification and expression analysis of b ZIP gene family in Carthamus tinctorius L.Sci. Rep.2020101552110.1038/s 41598-020-72390-z 32968100 PMC 7511407 · doi ↗ · pubmed ↗