Gut commensal Bifidobacterium longum confers resistance to Salmonella Typhimurium and Shigella flexneri in a Caenorhabditis elegans model
Phurt Harnvoravongchai, Samara Paula Mattiello, Achuthan Amabat, Jusail C. P., Syed M. Faisal, Radhey S. Kaushik, Joy Scaria

TL;DR
Bifidobacterium longum, a gut bacterium, protects against Salmonella and Shigella infections in a worm model, suggesting it could be a non-antibiotic treatment.
Contribution
Identifies Bifidobacterium longum as a gut commensal that inhibits Salmonella and Shigella through distinct mechanisms in a C. elegans model.
Findings
B. longum reduces pathogen burden and improves survival in C. elegans infected with Salmonella and Shigella.
B. longum inhibits Salmonella via acidification and Shigella via protein or heat-stable metabolites.
B. longum modulates host immune pathways like p38 MAPK and insulin/IGF-1 signaling.
Abstract
Salmonellosis and shigellosis remain major global health concerns, with Salmonella Typhimurium and Shigella flexneri classified as high-priority antibiotic-resistant pathogens by the World Health Organization. The development of new antibiotics is slow and challenging, underscoring the need for alternative therapeutic strategies. One promising approach involves leveraging gut microbiota-derived bacteria that confer colonization resistance against enteric pathogens. In this study, we screened a human gut microbiota culture collection and identified Bifidobacterium longum as the most effective species in inhibiting S. Typhimurium and S. flexneri in vitro. To evaluate its protective potential in vivo, we utilized Caenorhabditis elegans as a model system. Our findings demonstrate that B. longum significantly reduced pathogen burden and enhanced host survival following infection. Mechanistic…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
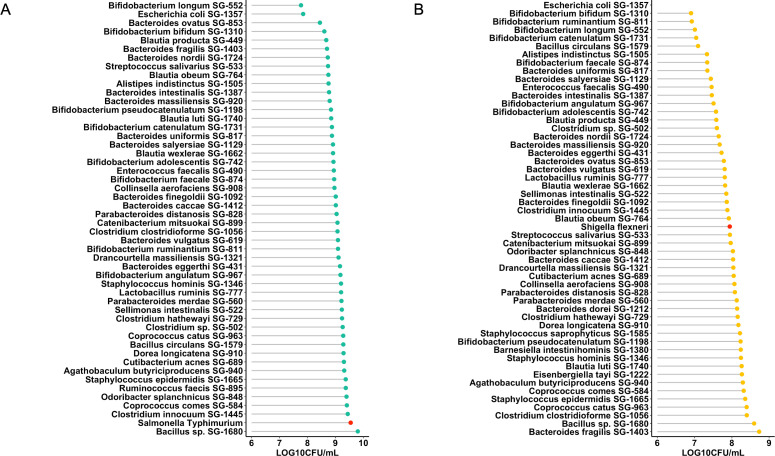 Fig 1
Fig 1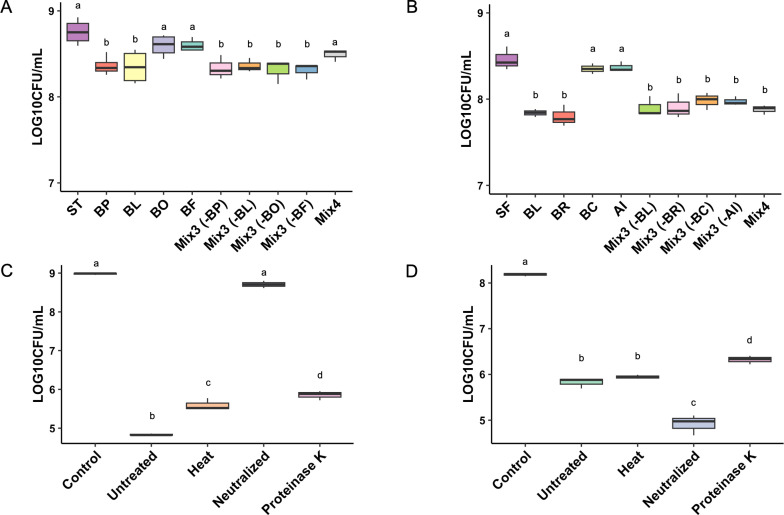 Fig 2
Fig 2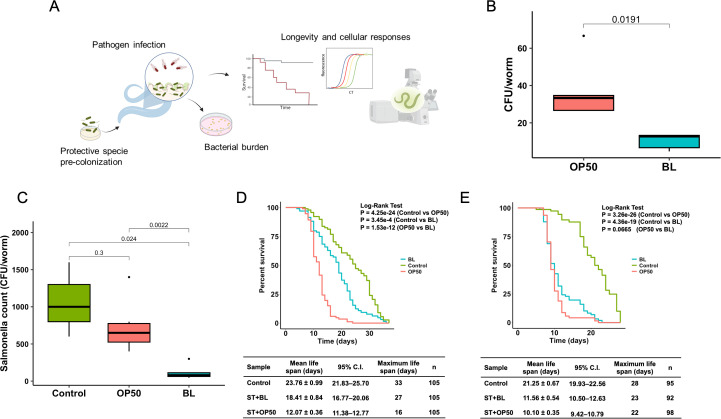 Fig 3
Fig 3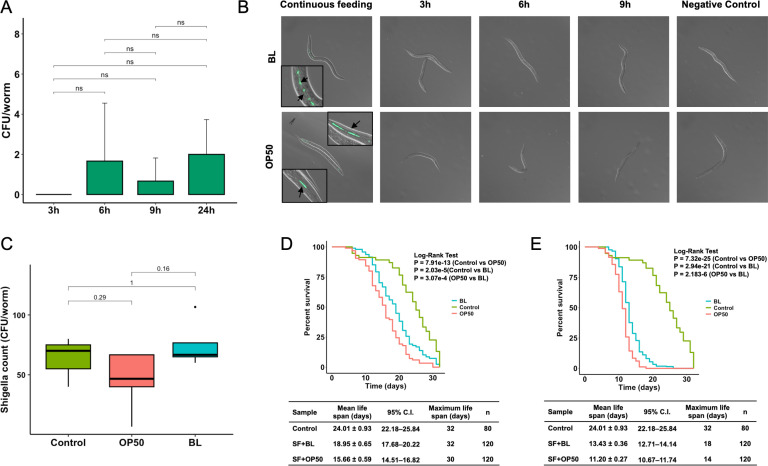 Fig 4
Fig 4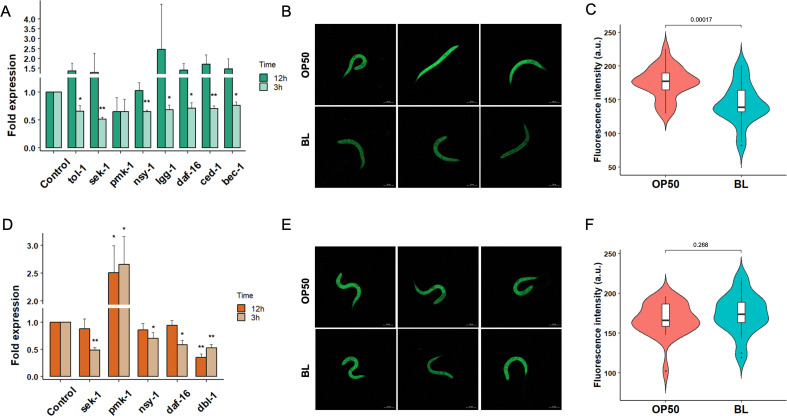 Fig 5
Fig 5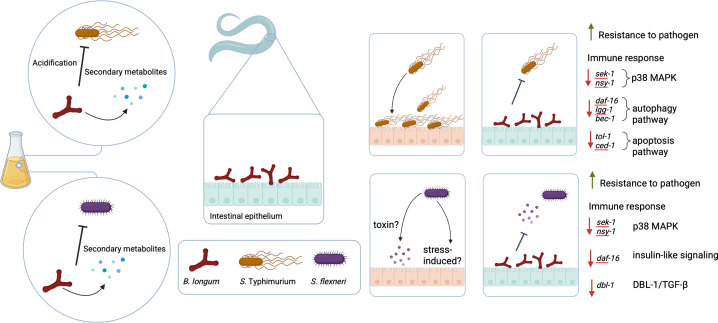 Fig 6
Fig 6Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsEscherichia coli research studies · Probiotics and Fermented Foods · Listeria monocytogenes in Food Safety
INTRODUCTION
The rise in antibiotic resistance among human bacterial pathogens presents a significant global challenge. The World Health Organization’s (WHO) 2024 bacterial priority pathogen list includes 24 pathogens across 15 families of antibiotic-resistant bacteria, underscoring the escalating concern (1). Several pathogens, particularly those classified as critical and high priority, exhibit multidrug resistance. This issue is exacerbated by the lengthy development and approval process for new-generation antibiotics, which typically requires 10–15 years from discovery to market authorization. Moreover, the success rate for novel classes of antibiotics remains low, with only one in 30 preclinical trials progressing to approval (2).
As a result, there is an urgent need to develop non-antibiotic alternatives to control multidrug-resistant bacteria. In this regard, bacteria from healthy gut microbiota constitute a rich source of pathogen-suppressing species, primarily through a mechanism known as colonization resistance (3–5). They outcompete pathogens for nutrients and attachment (6, 7), produce antimicrobial compounds such as bacteriocins and short-chain fatty acids, enhance gut barrier function by regulating tight junction proteins (8), and modulate the immune responses to promote host defense while preventing excessive inflammation (9–11). The colonization resistance effect on the gut microbiota has been leveraged in fecal microbiota transplantation (FMT), where fecal material from healthy patients is transferred to patients with antibiotic-refractory diarrhea. While FMT is highly effective as a mainstream treatment for diarrheal infections, challenges in large-scale preparation could limit its application, particularly to severe cases (12, 13).
Identifying specific bacterial species responsible for colonization resistance is challenging due to the gut microbiota’s complexity and the interconnected role of its members (14, 15). Multiple bacterial species interact in intricate ways, making it difficult to isolate individual effects (16, 17). Some bacteria indirectly enhance immune defenses, further complicating the distinction between direct inhibition and immune-mediated resistance. Additionally, functional overlap among closely related commensals adds another layer of complexity. However, recent efforts have led to the development of gut microbiota culture collections to address these challenges (17–20). In this study, we utilized a culture collection (20) to screen and identify candidate species capable of inhibiting Salmonella Typhimurium and Shigella flexneri. These two pathogens were selected for screening because both are listed as high-priority antibiotic-resistant pathogens by the WHO (1). Non-typhoidal Salmonella causes approximately 93.8 million infections annually, representing a major global health and economic burden (21, 22). S. Typhimurium has a broad host range that extends beyond humans to include livestock such as chickens and pigs, which play a significant role in its transmission and the occurrence of widespread outbreaks globally (23, 24). While effective vaccines exist for typhoid fever, few options are available for non-typhoidal salmonellosis, despite its broad host range and impact on livestock. Shigella similarly causes over 80 million infections each year, with more than 69% of cases occurring in children under 5 years of age (25, 26). For our screen, we employed both in vitro assays and an in vivo model. We chose Caenorhabditis elegans as the in vivo model due to its simplicity and natural ability to consume bacteria. Numerous human gut pathogens, including gram-negative Diarrheagenic Escherichia coli, Salmonella enterica, Vibrio cholerae, and gram-positive L. monocytogenes, have been examined using this model (27–31). Alterations in worm behavior, motility, and innate immune signaling pathways are commonly employed to study bacterial pathogenesis and host responses (31, 32). This combined in vitro/in vivo approach enabled a rapid platform to screen microbiota strains for pathogen inhibition. Our screen identified Bifidobacterium longum as the most effective species against both Salmonella and Shigella. This strain also showed significant protection in the C. elegans model through innate immune modulation and enhanced host defense responses.
RESULTS
Screening and identification of bacterial species capable of inhibiting Salmonella and Shigella
Our group previously developed a microbiota culture collection from healthy human donors that represented 70% of the functional capacity of the gut microbiota (20). We used this culture collection to identify species capable of inhibiting S. Typhimurium and S. flexneri growth. Since growth rate is a critical factor in competitive exclusion and fast-growing bacteria have strong potential for colonization resistance due to their efficiency in competing for nutrients and occupying available niche spaces, approximately 52 fast-growing species from this collection were chosen for this screen using co-culture assays. Most of the strains tested inhibited Salmonella to varying degrees, except for Bacillus sp. SG-1680, which promoted Salmonella growth (Fig. 1A). B. longum SG-552 and E. coli SG-1357 exhibited the most pronounced inhibitory effects with 1.77 and 1.70 log reductions, respectively. In the case of Shigella, inhibition was limited to 29 strains, whereas 23 species enhanced Shigella growth (Fig. 1B). Bacteroides fragilis SG-1403 exhibited the highest enhancement of Shigella growth, by nearly 1 log. In contrast, this species was the most effective at inhibiting Salmonella. The bacterial species exhibiting the most significant inhibitory effects on the pathogens: B. longum SG-552, E. coli SG-1357, Bacteroides ovatus SG-853, Bifidobacterium bifidum SG-1310, Blautia producta SG-449, Bacteroides fragilis SG-1403, Bifidobacterium ruminantium SG-811, Bifidobacterium catenulatum SG-1731, Bacillus circulans SG-1579, and Alistipes indistinctus SG-1505 were selected for further investigation. To assess the safety profile of these species, a cell invasion assay using human epithelial Caco-2 cells was performed. The results showed that B. bifidum, B. circulans, and E. coli actively invaded the Caco-2 cells. No invasion of Caco-2 cells by A. indistinctus, B. catenulatum, B. fragilis, B. longum, B. ovatus, and B. producta was observed, while relatively low invasion was observed for B. ruminantium (Fig. S1). Therefore, species that demonstrated substantial inhibitory effects with minimal cell invasion were chosen as candidates for subsequent experiments.
Screening of bacteria from human culture collection to identify species capable of inhibiting enteric pathogens. (A) Inhibitory effects of 49 species on Salmonella growth. (B) Inhibitory range of 52 species against Shigella. Co-cultures were incubated anaerobically for 24 h at a 9:1 ratio (bacteria:pathogen). The growth of the pathogen alone is indicated by a red dot in the lollipop plot. Values represent the mean from three independent experiments.
Having identified species with relatively low cell invasiveness that could inhibit both pathogens, we next tested whether additive inhibition could be obtained by a pool of all these species. Based on the pathogen inhibition rank, we selected the best four species and constructed a synthetic consortium. Single-species dropout experiments were also conducted by omitting one to demonstrate the importance of individual species in the consortium. When compared to individual species inhibition capacity, no mix improved the inhibition of Salmonella or Shigella (Fig. 2A and B), clearly showing that the inhibitory capacity is not additive. Notably, B. longum alone significantly reduced the growth of both Salmonella and Shigella in the co-culture assay, with inhibitory effects indistinguishable from those of bacterial mixes.
In vitro inhibition of pathogens by a synthetic bacterial consortium. Inhibitory effects of bacterial mixes (Mix3 and Mix4) on (A) Salmonella (ST) and (B) Shigella (SF). Mixes were composed of selected strains (BP, BL, BO, BF, BR, BC, and AI). BP, B. producta; BL, B. longum; BO, B. ovatus; BF, B. fragilis; BR, B. ruminantium; BC, B. circulans; AI, A. indistinctus. Inhibition of (C) Salmonella and (D) Shigella by spent media from BL cultured for 48 h under various treatment conditions. Control: pathogen cultures in fresh modified brain heart infusion (mBHI) medium. The experiment was performed in triplicate, and statistical analysis was conducted using ANOVA and Tukey HSD test. Different letters indicate statistically significant differences (P < 0.05).
Pathogen suppression by commensal bacteria may be attributed to multiple mechanisms, including direct nutritional competition, production of bacteriocins and secondary metabolites, and pH alteration (33–35). To elucidate which of these mechanisms is responsible for suppression, we compared the inhibitory effects of spent media on both Salmonella and Shigella under various treatment conditions. Spent media of B. longum significantly inhibited Salmonella growth, resulting in a four-log reduction. The treatment of the spent media with heat or Proteinase K slightly decreased this inhibition, suggesting the involvement of proteins and heat-labile components produced by B. longum (Fig. 2C). Organic acid production by probiotics, primarily lactic and acetic acid, creates an acidic environment that is well known to inhibit pathogen growth. To determine whether acidification is responsible for the observed antimicrobial activity, pH adjustment is commonly performed. As expected, neutralizing the pH substantially reduced the inhibitory effect, emphasizing the importance of acidic conditions produced by B. longum in inhibiting Salmonella growth. In contrast, Shigella growth was inhibited by approximately two logs in untreated spent media, with heat treatment resulting in no significant effect on inhibition (Fig. 2D). A slight reduction in the level of inhibition was observed after Proteinase K treatment. Notably, the neutralized spent media exhibited the highest degree of inhibition of Shigella, suggesting that a pH-independent component is responsible for the suppression of Shigella by B. longum spent media. In summary, the spent media results indicate that the mechanism of Salmonella inhibition by B. longum is pH reduction, whereas Shigella inhibition occurs through a protein-mediated mechanism or a heat-stable secondary metabolite.
B. longum colonization reduces Salmonella load and enhances pathogen resistance in a C. elegans model
To determine whether pathogen suppression by B. longum observed in vitro could be replicated in vivo, an animal experiment was conducted using C. elegans. Worms were pre-colonized with B. longum and subsequently infected with Salmonella to assess bacterial load and the effects on worm lifespan (Fig. 3A). B. longum successfully colonized and persisted in C. elegans following a 2-day pre-colonization period (Fig. 3B). However, the quantity of B. longum was significantly lower than that of E. coli OP50 (OP50), which is conventionally used as feed for C. elegans. As demonstrated in Fig. 3C, a 3 h exposure period was sufficient for Salmonella to successfully colonize and induce acute infection in C. elegans. Pre-colonization of C. elegans with B. longum resulted in significantly enhanced Salmonella clearance, achieving a one-log reduction compared with worms fed OP50 and the control (dead OP50). The survival of worms infected with Salmonella was evaluated under two distinct conditions. Continuous feeding of the worms with Salmonella was conducted to replicate chronic exposure, a standard procedure for assessing the pathogen’s ability to kill the worms, and acute exposure by introducing C. elegans to the pathogen for 3 h before transferring them to a pathogen-free environment to simulate the normal course of infection. Both continuous and brief exposure to Salmonella significantly affected the lifespan of worms (Fig. 3D and E). Colonization with B. longum results in substantial protection against Salmonella during acute infection. The percent survival of C. elegans pre-colonized with B. longum was significantly higher (P < 0.001) with a mean life span of 18.41 ± 0.84 days, compared to 12.07 ± 0.36 days in the worms fed with OP50 (Fig. 3D). However, no protective effect of B. longum was observed when C. elegans was continuously exposed to Salmonella. In conclusion, B. longum had a protective effect against acute Salmonella infection in a C. elegans model, and this protective mechanism appears to be independent of niche competition. Furthermore, the inhibitory effect of B. longum could potentially be extended beyond non-typhoidal strains to include typhoidal strains, thereby broadening its therapeutic applications.
B. longum confers protection against Salmonella infection in a C. elegans model. (A) Schematic of the experimental design. Image was created with Biorender. (B) Colonization efficiency of B. longum (BL) compared to E. coli OP50 (OP50) in C. elegans 48 h post-feeding. (C) Salmonella burden in worms precolonized with BL or OP50, measured 48 h post-infection. C. elegans pre-colonized with dead OP50 was used as the Control. Survival of precolonized worms following (D) acute or (E) chronic Salmonella (ST) exposure. Lifespan assessed by Kaplan–Meier analysis (log-rank test). The mean lifespan represents the mean value ± standard deviation of worms from different treatments. Control; no ST infection worms.
Shigella reduces worm life span, and B. longum increases longevity
To determine the effect of Shigella on C. elegans and the potential protective role of B. longum, analogous to the experimental model employed for Salmonella, both acute and chronic exposure of C. elegans to Shigella were investigated. Initially, we were unable to quantify Shigella loads in C. elegans transiently exposed to the pathogen, in contrast to those that were continuously exposed. This observation led us to postulate that Shigella does not persist in the intestinal tract of C. elegans. To test this hypothesis, we examined Shigella colonization in C. elegans by varying the exposure duration from 3 h to 24 h before transferring the worms to a pathogen-free environment. Colony-forming units (CFU) were assessed once the worms reached the young-adult stage. As anticipated, we were unable to observe any CFU of Shigella from C. elegans, irrespective of the exposure period (Fig. 4A). The inability of the pathogen to persist in C. elegans was further corroborated by visualization of GFP-tagged bacteria. No GFP signal was detected after temporary feeding of C. elegans with bacteria for 3 h, 6 h, or 9 h, supporting our findings from the previous set of experiments (Fig. 4B). Consequently, Shigella burden in C. elegans was assessed under chronic exposure conditions. In contrast to B. longum’s protection against Salmonella, the Shigella load in the worms was comparable across all treatments (Fig. 4C), suggesting that colonization by B. longum is insufficient to reduce the Shigella burden in C. elegans. Despite the inability of Shigella to persist in C. elegans, worm longevity was significantly reduced following both acute and chronic exposure to the pathogen. This indicates that stable colonization by Shigella is not necessary to induce worm mortality. The observed effect may be driven by colonization-independent mechanisms, such as toxin-mediated damage (e.g., Shiga toxin or enterotoxins) or stress-induced responses to pathogen exposure. Further studies are needed to validate these possibilities and elucidate the underlying mechanisms of host mortality. The survival assay of C. elegans under acute bacterial infection demonstrated a statistically significant difference in worm life span (P < 0.001), with the mean life span extending from 15.66 ± 0.56 days in OP50-fed worms to 18.95 ± 0.65 days in worms pre-colonized with B. longum (Fig. 4D). The capacity of B. longum to enhance pathogen suppression in C. elegans was also demonstrated under chronic exposure conditions, as evidenced by the increased mean lifespan of 13.43 ± 0.36 days in C. elegans pre-colonized with B. longum compared to those fed with OP50 (11.20 ± 0.27 days). These results suggest that exposure to Shigella significantly affects worm longevity independently of pathogen colonization, whereas B. longum colonization enhances worm longevity.
B. longum enhances C. elegans survival during Shigella infection. (A) Shigella load in worms exposed for 3 h, 6 h, 9 h, or 24 h, quantified 48 h post-infection (ANOVA, Tukey HSD; P < 0.05). (B) Confocal imaging of GFP-labeled Shigella in worms precolonized with B. longum (BL) or E. coli OP50 (OP50). (C) Shigella burden in worms pre-colonized with BL or OP50, assessed 48 h after continuous exposure. C. elegans pre-colonized with dead OP50 served as the Control. Survival of precolonized worms following (D) acute or (E) chronic Shigella (SF) exposure. Lifespan assessed by Kaplan–Meier analysis (log-rank test). The mean lifespan represents the mean value ± standard deviation of worms from different treatments. Control; no SF infection worms.
B. longum causes suppression of genes and pathways associated with the host defense during Salmonella and Shigella infections
Previously, we reported that the colonization of C. elegans with B. longum resulted in the upregulation of innate immune and age-associated genes (36). To investigate differential host responses to Salmonella and Shigella infections, the genes and pathways associated with host defense were analyzed in C. elegans using qRT-PCR. Three hours post-infection, downregulation of the p38 MAPK pathway was observed in C. elegans pre-colonized with B. longum, as evidenced by the reduced levels of SAPK/ERK kinase-1 (sek-1) and neuronal symmetry (nsy-1) compared with those in worms fed OP50. Relative to OP50 feeding, colonization with B. longum resulted in the suppression of forkhead box protein O (daf-16), LC3, GABARAP and GATE-16 family (lgg-1), and beclin homolog 1 (bec-1), leading to the attenuation of autophagy in C. elegans in response to Salmonella infection. Additionally, a decrease in the expression of toll-like receptor (tol-1) and cell death abnormality protein 1 (ced-1), which are related to apoptosis in C. elegans, was observed (Fig. 5A). No significant changes in gene expression were observed 12 h post-infection. The C. elegans transgenic mutant CF1139 expressing DAF16::GFP was used to confirm the expression level of DAF16 using confocal microscopy. A relatively lower GFP signal was detected in CF1139 worms pre-colonized with B. longum during Salmonella infection than in OP50 feeding (Fig. 5B and C). Therefore, it appears that the protective effect of B. longum against Salmonella infection in C. elegans occurs through host immune modulation, which could be indicated by the downregulation of host defense genes related to pathogen infection. We also analyzed the differences in gene expression related to the host response in C. elegans pre-colonized with B. longum compared with OP50 during Shigella infection (Fig. 5D). Significant reductions in the expression of genes and pathways contributing to the immune response, including sek-1, nsy-1, daf-16, and Dpp and BMP-like protein 1 (dbl-1), were observed 3 h post-infection. While most responses were restored 12 h post-infection, the repression of dbl-1 remained evident. Our findings demonstrate that B. longum attenuates Shigella infection by suppressing genes involved in the innate immune response of C. elegans. Despite changes in mRNA levels, no difference in DAF16::GFP expression was observed in CF1139 worms (Fig. 5E and F ).
*Suppression of innate immunity indicates the protective effects of B. longum against pathogenic infections in C. elegans. Differential gene expression in C. elegans upon exposure to (A) Salmonella and (D) Shigella at 3 and 12 h post-infection. Gene expression was quantified by qRT-PCR and normalized to that of the housekeeping gene pmp-3. Expression of DAF16::GFP in CF1139 transgenic C. elegans mutants in response to (B and C) Salmonella or (E and F) Shigella at 3 h post-infection, compared between worms fed B. longum (BL) to those fed E. coli OP50 (OP50). Worms (N = 25) from three biological replicates were used for the quantitative analysis of fluorescence intensity using FIJI software. Statistical differences between groups were analyzed using Student t-test (*P < 0.05 and *P < 0.01).
DISCUSSION
Using a previously established healthy human microbiota culture collection (20), we conducted a high-throughput screen to identify strains capable of inhibiting S. Typhimurium and S. flexneri. In vitro co-culture assays revealed pathogen-inhibiting, inhibition-neutral, and pathogen growth-enhancing strains. Most strains tested against Salmonella showed mild inhibition or had no effect on pathogen growth. Notably, the inhibiting strains primarily belonged to the genus Bifidobacterium, while numerous moderately inhibitory strains were from the genus Bacteroides. Similar trends were observed with Shigella. However, a key difference was that many strains markedly increased the abundance of Shigella, several of which were members of Bacteroidetes. This unanticipated finding aligns with the known trends in gut ecology. Pathogens typically exist in low numbers in the mammalian because the diverse microbial community prevents their overgrowth. However, gut dysbiosis can disrupt this balance, facilitating both pathogens and many co-colonizing species to proliferate (37). Bacteroides thetaiotaomicron, a known gut symbiont that aids dietary sugar degradation and improves gut maturation (38), can also worsen infections by enterohemorrhagic E. coli (39). It should be noted that the outcome of inhibition assays may be influenced by the ratio of the bacterial strain to the pathogen. Further investigation into dose-dependent effects would provide additional justification for these interactions.
B. longum and E. coli showed strong inhibition against both Salmonella and Shigella. Due to the invasiveness of the E. coli strain, only B. longum was utilized for subsequent mechanistic investigations. Synthetic mixtures comprising other inhibiting strains, with and without B. longum, did not enhance the inhibition when compared to that of B. longum alone. This outcome was unexpected, as additive inhibitory effects have been observed in multiple species combinations against enteric pathogens like Clostridium difficile (20, 40, 41). The lack of additive or enhanced inhibition observed in synthetic mixtures, compared to individual strains, may suggest potential competition among community members. This could result from antagonistic interactions, such as interference from strain-specific metabolites, or niche saturation that limits the overall carrying capacity and population growth. These factors may reduce the collective inhibitory effect, highlighting the complexity of designing effective multi-strain consortia. The ability of a single gut commensal to inhibit multiple enteric pathogens has been reported only to a limited extent. Our results demonstrate that B. longum possesses dual-pathogen inhibition capacity, highlighting its potential as a broad-spectrum probiotic candidate. To better understand the factors responsible for inhibition, we analyzed the effects of the spent supernatant. Salmonella inhibition appeared to be driven by acidification, as buffered supernatants lost most of their inhibitory capacity. In contrast, Shigella inhibition was reduced by heat and proteinase K treatment, indicating involvement of heat-stable metabolites or proteinaceous components. Bacteria within the Bifidobacterium genus are known to produce a variety of antimicrobial metabolites, including short-chain fatty acids and bacteriocins. Therefore, the B. longum strain likely employs a combination of these mechanisms to inhibit pathogenic bacteria (42, 43). Further characterization of these components using proteomic and metabolomic analyses is necessary.
Although the in vitro screen provided key initial insights, it did not account for host-related factors such as immune responses and other host defense mechanisms. To ensure the safety of the probiotic candidate, we confirmed that the B. longum strain, obtained from a healthy human fecal sample, was non-invasive in Caco-2 cell assays. In addition, B. longum has been extensively studied and is generally recognized as safe in mammalian models (44, 45). Following this, C. elegans was used in an in vivo model to further assess host-microbe interactions and probiotic effects. This worm is widely used in microbiome research due to its short life cycle, ease of handling, and well-characterized immune responses (46, 47). Salmonella infections, both acute and chronic, significantly reduced worm lifespan. However, B. longum extended survival in the acute infection model, suggesting a protective effect. No protection was observed in the chronic model. This may be due to a differing ratio of B. longum to the pathogen (48). During acute infection, limited exposure (3 h) may have allowed pre-colonized B. longum to effectively inhibit Salmonella proliferation. In continuous exposure, increased pathogen burden may have overwhelmed B. longum’s protective effect. Pathogen count and fluorescence images revealed that Shigella was unable to colonize and establish a persistent infection in C. elegans. The worm grinder likely disrupts ingested Shigella (49, 50). The virulence plasmid has been reported to be essential for S. flexneri accumulation in C. elegans (51), and the strain used in our study may have lacked this component, which could explain its inability to colonize the worm intestine. S. flexneri was selected in this study due to its well-characterized pathogenesis, making it a suitable representative for investigating bacterial infection and potential intervention strategies. Although B. longum did not reduce S. flexneri burden in C. elegans model, it appeared to enhance host survival following infection. Future studies evaluating the protective effect of B. longum against other clinically relevant Shigella species would provide more insight into the protective capabilities of this strain.
Analysis of C. elegans host defense and innate immune genes revealed significant modulation of these genes during B. longum colonization and pathogen infection. Salmonella infection is known to trigger p38 MAPK signaling pathway in C. elegans (52). Our RT-qPCR results confirmed that B. longum colonization suppressed expression of p38 MAPK-associated genes after 3 h of Salmonella infection, compared to worms fed with OP50. Reduced mRNA levels of lgg-1, bec-1, and daf-16 suggest altered regulation of genes involved in the autophagic response, a key mechanism for C. elegans to defend against various pathogens, including Salmonella (53). This is further supported by the results of fluorescent-labeled proteins, as the process is activated under the regulation of DAF-16. Furthermore, the downregulation of tol-1, a gene predicted to be involved in pathogen recognition (54), was also observed, supporting the finding that B. longum enhances pathogen clearance.
Compared to Salmonella, Shigella has been relatively less studied in C. elegans. Therefore, we analyzed genes involved in p38 MAPK, DBL-1/TGF-β, and the insulin-like signaling pathway, which are key regulators of intestinal defense (55). As hypothesized, the expression of immune-related genes was predominantly downregulated in B. longum-colonized worms compared to those fed OP50 following Shigella infection. Conversely, we observed an increase in pmk-1 expression levels at 3 h and 12 h post-infection, potential crosstalk between p38 MAPK and other pathways through PMK-1 regulation (56, 57). Our recent study reported that feeding C. elegans with B. longum extends worm longevity and alters the expression of genes associated with autophagy and lysosomal functions (36). This supports the idea that autophagy may contribute to B. longum-mediated colonization resistance. It should be noted that changes in mRNA levels do not always correlate with protein function; therefore, future experiments focusing on protein-level analyses, such as proteomic studies or reporter-based assays, are necessary to validate these findings. While C. elegans is a valuable model for studying probiotics and host-microbe interactions, its lack of an adaptive immune system and simplified circulatory system may limit its ability to fully demonstrate the complex immune responses seen in higher organisms. This absence restricts the model’s translational relevance, particularly for understanding interactions that involve adaptive immunity mechanisms. Therefore, further validation in mammalian models, such as studies of colonization resistance in gnotobiotic systems, is essential to provide a more comprehensive assessment of probiotic effects and host responses.
Our findings reveal that B. longum, isolated from the human gut, can inhibit the growth of Salmonella and Shigella (Fig. 6). The acidic environment created by B. longum significantly impairs Salmonella growth, whereas suppression of Shigella likely involved a secreted protein or heat-stable metabolite. Additionally, B. longum colonization modulated innate immune pathways during pathogen exposure, indicating its potential to attenuate the severity of infection. While B. longum has potential as a non-antibiotic probiotic candidate, additional studies in mammalian models are required to validate these findings and evaluate their clinical relevance.
Summary of the pathogen-inhibitory effect of B. longum and its contribution to colonization resistance in C. elegans, potentially through modulation of immune pathways. The image was created with Biorender.
MATERIALS AND METHODS
Bacterial strains and culture conditions
The bacterial species used in this study were selected from a human gut culture library previously constructed in our laboratory (20). All species were cultured and maintained in modified brain heart infusion (mBHI) medium at 37°C under anaerobic conditions with 85% CO_2_, 10% H_2_, and 5% N_2_. Salmonella Typhimurium 4,[5],12:i:, isolated from swine samples (58), and S. flexneri ATCC 12022 were cultured in Luria-Bertani medium. Xylose Lysine Tergitol 4 (XLT4) agar (BD Difco, Houston, TX, USA) was used for the selective identification of Salmonella and Shigella.
High-throughput screening and co-culture assays
Screening of bacterial strains capable of inhibiting S. Typhimurium and S. flexneri was performed using a coculture assay. Overnight cultures of bacteria in mBHI broth were adjusted to an OD_600_ of 0.5, and co-cultivation with the pathogen was performed at a ratio of 9:1 in a final volume of 1 mL mBHI. After 24 h of incubation at 37°C in an anaerobic environment, the bacterial suspensions were serially diluted 10-fold in phosphate-buffered saline (PBS), and pathogen counts were enumerated using the drop plate method on XLT4 agar. Cultures containing only S. Typhimurium or S. flexneri were used as untreated controls. The degree of pathogen inhibition was assessed by comparing colony counts to the control after 24 h. For combination mixtures, individual strains adjusted to 0.5 OD_600_ were mixed to create Mix3 or Mix4 and tested in the co-culture assay with a consistent ratio of 9:1.
Caco-2 cell invasion assay
Human colorectal adenocarcinoma (Caco-2) cells were used to evaluate the invasive potential of the bacterial isolates. Overnight cultures of each bacterial strain were used to infect Caco-2 cells at a multiplicity of infection of 1:100 and incubated for 1 h at 37°C. Following infection, the bacterial suspensions were removed, and the cells were washed twice with PBS. A Dulbecco's modified eagle medium (DMEM) solution supplemented with 100 µg/mL gentamicin was added to the cells and incubated for an additional hour to ensure the elimination of extracellular bacteria. Subsequently, the cells were lysed with 1% Triton X-100 under anaerobic conditions, and intracellular bacteria were quantified by CFU counts on mBHI agar.
Spent media assay
To prepare the spent medium, B. longum was cultured in mBHI broth for 48 h to reach the stationary phase. The culture was then collected by centrifugation and filtered through a 0.22 µm filter to remove residual bacterial cells. The resulting filtrate was divided into four portions, each subjected to different treatment conditions. Heat treatment involved heating the filtrate to 90°C for 1 h to preserve heat-stable components while inactivating heat-labile compounds. For protein removal, the filtrate was incubated with Proteinase K (1 mg/mL) for 2 h. To obtain neutralized spent media, the pH of the filtrate was adjusted to ~7.0 with NaOH, followed by a second filtration through a 0.22 µm filter. The inhibition efficacy of the treated spent media was evaluated by incubating 0.5 OD_600_ of Salmonella or Shigella with 500 µL of the treated media under anaerobic conditions at 37°C for 24 h. Pathogen growth was assessed by CFU on XLT4 agar plates. A negative control was performed using fresh mBHI broth.
Bacterial colonization and pathogen infection in C. elegans
Wild-type N2-Bristol C. elegans were maintained on nematode growth medium (NGM) plates (RPI, Mt Prospect, IL) seeded with E. coli OP50 at 20°C. Synchronization was achieved via bleach treatment, allowing eggs to hatch for 14–16 h to obtain synchronized L1 stage worms. Bacterial colonization was performed as described previously (36). Briefly, synchronized L1 worms were exposed to a concentrated lawn of E. coli OP50 or B. longum in an anaerobic chamber for 3 h. Following exposure, worms were transferred to a biosafety cabinet for aeration for an additional hour. Worms were then washed three times with M9 buffer and transferred to an NGM plate seeded with dead OP50 prepared as previously described (59). After a 2-day colonization period, 20–30 worms were picked and surface-sterilized using a 1:1,000 bleach solution in M9 buffer. The worms were washed three times and resuspended in anaerobic PBS before being lysed with a motorized pestle in the anaerobic chamber. The number of B. longum colonies in C. elegans was determined by CFU counts on mBHI agar. To investigate the protective effect against pathogens, C. elegans pre-colonized with B. longum or OP50 for 24 h were briefly exposed to a concentrated pathogen for 3 h. Following exposure, worms were surface-sterilized and washed three times before being transferred to NGM plates seeded with a lawn of dead OP50. Pathogen load was determined by counting CFU on XLT-4 agar plates.
C. elegans survival assay
For short-term exposure to pathogens, 30–40 pre-colonized L3/L4 worms were fed a concentrated lawn of Salmonella or Shigella for 3 h. The worms were then surface sterilized and washed three times before being transferred to fresh NGM plates seeded with dead OP50. To evaluate the protective effect under continuous pathogen exposure, worms were constantly fed pathogens on NGM plates. The number of dead worms was recorded daily, and worms that crawled off the plates were excluded from the analysis.
RNA isolation and qRT-PCR
Total RNA was extracted using the TRIzol reagent (Thermo Fisher Scientific, Waltham, MA, USA). Worms were snap frozen in liquid nitrogen and subjected to multiple freeze-thaw cycles to ensure complete lysis. Reverse transcription was performed using ProtoScript II Reverse Transcriptase (New England BioLabs), following the manufacturer’s instructions. The resulting cDNA was used for qRT-PCR using the Power SYBR Green PCR Master Mix (Applied Biosystems, Waltham, MA, USA). The peroxisomal membrane protein-related gene (pmp-3) was used as the reference gene, and fold changes in gene expression were calculated using the 2^−ΔΔCt^ method.
C. elegans fluorescence imaging and quantification
To evaluate Shigella burden in C. elegans, L3/L4 worms were temporarily fed *Shigella-*GFP for 3 h, 6 h, or 9 h, then transferred to NGM plates seeded with a lawn of dead OP50. Worms at the young adult stage were surface sterilized, washed three times, and visualized under a confocal microscope (Zeiss LSM 980). The DAF16::GFP-expressing worm strain CF1139 was used to assess gene expression. Worms expressing L3/L4 GFP were briefly exposed to pathogens for 3 h. Three hours post-infection, the worms were fixed with 4% paraformaldehyde in PBS, washed three times, and visualized using a confocal microscope. Quantification of the GFP signal was performed using the FIJI analysis tool (ImageJ).
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Sati H, Carrara E, Savoldi A, Hansen P, Garlasco J, Campagnaro E, Boccia S, Castillo-Polo JA, Magrini E, Garcia-Vello P, Wool E, Gigante V, Duffy E, Cassini A, Huttner B, Pardo PR, Naghavi M, Mirzayev F, Zignol M, Cameron A, Tacconelli E, WHO Bacterial Priority Pathogens List Advisory Group. 2025. The WHO Bacterial Priority Pathogens List 2024: a prioritisation study to guide research, development, and public health strategies against antimicrobial resistance. Lancet Infect Dis 25:1033–1043. doi · doi ↗ · pubmed ↗
- 2Organization WHO. 2022. 2021 antibacterial agents in clinical and preclinical development: an overview and analysis. World Health Organization.
- 3van der Waaij D, Berghuis-de Vries JM, Lekkerkerk Lekkerkerk-v. 1971. Colonization resistance of the digestive tract in conventional and antibiotic-treated mice. J Hyg (Lond) 69:405–411. doi:10.1017/s 00221724000216534999450 PMC 2130899 · doi ↗ · pubmed ↗
- 4Koopman JP, Kennis HM, Mullink JW, Prins RA, Stadhouders AM, De Boer H, Hectors MP. 1984. “Normalization” of germfree mice with anaerobically cultured caecal flora of “normal” mice. Lab Anim 18:188–194. doi:10.1258/0023677847808912536379286 · doi ↗ · pubmed ↗
- 5Pultz NJ, Stiefel U, Subramanyan S, Helfand MS, Donskey CJ. 2005. Mechanisms by which anaerobic microbiota inhibit the establishment in mice of intestinal colonization by vancomycin-resistant Enterococcus. J Infect Dis 191:949–956. doi:10.1086/42809015717271 · doi ↗ · pubmed ↗
- 6Wilson KH, Perini F. 1988. Role of competition for nutrients in suppression of Clostridium difficile by the colonic microflora. Infect Immun 56:2610–2614. doi:10.1128/iai.56.10.2610-2614.19883417352 PMC 259619 · doi ↗ · pubmed ↗
- 7Horrocks V, King OG, Yip AYG, Marques IM, Mc Donald JAK. 2023. Role of the gut microbiota in nutrient competition and protection against intestinal pathogen colonization. Microbiology (Reading) 169:001377. doi:10.1099/mic.0.00137737540126 PMC 10482380 · doi ↗ · pubmed ↗
- 8Gou H-Z, Zhang Y-L, Ren L-F, Li Z-J, Zhang L. 2022. How do intestinal probiotics restore the intestinal barrier? Front Microbiol 13:929346. doi:10.3389/fmicb.2022.92934635910620 PMC 9330398 · doi ↗ · pubmed ↗