The secretome of the environmental bacterium Pseudomonas protegens PBL3 has broad-spectrum antimicrobial activity against plant and human pathogenic bacteria
Biwesh Ojha, Kiyara Grosz, Clemencia M. Rojas

TL;DR
A soil bacterium called Pseudomonas protegens PBL3 produces a mix of antimicrobial molecules that can fight both plant and human pathogens, offering a new approach to combat antibiotic resistance.
Contribution
The study identifies the secretome of Pseudomonas protegens PBL3 as a novel, broad-spectrum antimicrobial source effective against multiple plant and human pathogens.
Findings
The secretome of Pseudomonas protegens PBL3 inhibits the growth of plant pathogen Burkholderia sp. and human pathogens like Acinetobacter baumannii and Staphylococcus aureus.
Growth inhibition levels ranged from 25% to 95% across tested pathogens.
The secretome's diverse chemical composition may reduce resistance development in bacteria.
Abstract
Antimicrobials are widely used to manage bacterial diseases in humans, pets, and livestock. Unfortunately, their extensive use has contributed to the emergence of antimicrobial resistance (AMR), a threat to global health. In contrast to the widespread antimicrobial use in medicine and veterinary, the use of antimicrobials to control bacterial diseases in plants is limited, posing a threat to global food security. These two threats highlight the urgent need to discover novel antimicrobial compounds with new modes of action. Previously, we demonstrated that the environmental bacterium Pseudomonas protegens PBL3 inhibits the growth of the plant pathogenic bacterium Burkholderia glumae, indicating that this strain produces and secretes potent antimicrobial compounds. In this study, we showed that the P. protegens PBL3 secretome also shows a broad spectrum of activity against plant…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
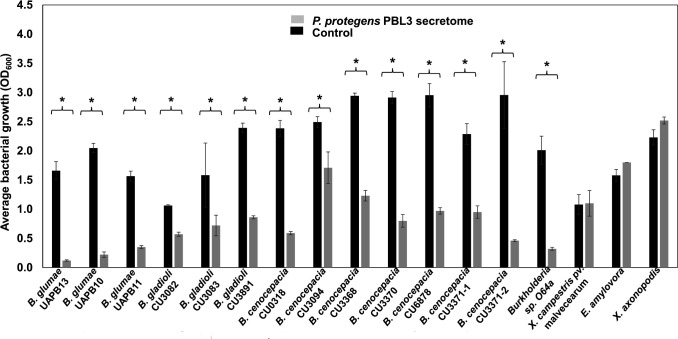 Fig 1
Fig 1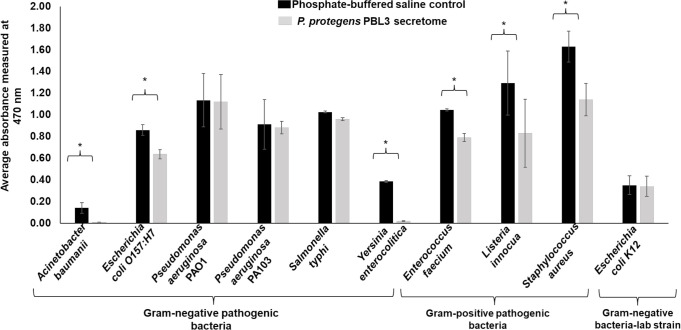 Fig 2
Fig 2| Bacterial strains | Reference/source |
|---|---|
|
| Martin Conda-Sheridan, University of Nebraska Medical Center |
| ( | |
| ( | |
| ( | |
| ( | |
| ( | |
| ( | |
| ( | |
| ( | |
| ( | |
| ( | |
| Yeshi Wamishe, University of Arkansas, Fayetteville | |
| Yeshi Wamishe, University of Arkansas, Fayetteville | |
| Yeshi Wamishe, University of Arkansas, Fayetteville | |
| Steven Beer, Cornell University | |
|
| Food Processing Center, University of Nebraska-Lincoln |
|
| Elena Garcia, University of Arkansas, Fayetteville |
| ATCC 10798 | |
| U.S. Department of Agriculture-Agriculture Research Service | |
|
| ATCC 33090 |
|
| Jim Alfano, University of Nebraska-Lincoln |
|
| Jim Alfano, University of Nebraska-Lincoln |
| ( | |
|
| Food Processing Center, University of Nebraska-Lincoln |
|
| ATCC 25923 |
|
| Craig Rothrock, University of Arkansas, Fayetteville |
|
| Jim Alfano, University of Nebraska-Lincoln |
- —Nebraska Research InitiativeUniversity of Nebraska-Lincoln
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsInfections and bacterial resistance · Plant Pathogenic Bacteria Studies · Plant-Microbe Interactions and Immunity
INTRODUCTION
Antimicrobials to control bacterial diseases have long been used to manage bacterial infections in humans, pets, livestock, and aquatic animals as therapeutic or prophylactic agents (1–4). However, this widespread use of antimicrobials has significantly contributed to the emergence of antimicrobial resistance (AMR) (4). The AMR threat is becoming a matter of global concern that has not been mitigated, as most of the recently approved antimicrobials are derivatives of previously known classes (5, 6), and bacteria quickly evolve mechanisms to render them ineffective. In contrast to the widespread use of antimicrobials in the medical and veterinary fields, the use of antimicrobials to combat bacterial diseases in crops has been limited, as effective antimicrobials for those pathogens have not been identified, have proven ineffective, or have only been approved for agricultural use for a limited number of bacterial pathogens (7, 8). Thus, both the threat of antimicrobial resistance in the medical and veterinary fields and the absence of effective antimicrobials in agriculture necessitate the discovery of new molecules with unique chemistries and modes of action.
Historically, naturally occurring microorganisms have been a prolific source of antimicrobial compounds since the discovery of penicillin from the fungus Penicillium notatum that marked the beginning of the antimicrobial era and demonstrated the therapeutic potential of microbial metabolites (9). Since then, various fungal species have yielded diverse classes of antimicrobials. Beyond fungi, bacteria, particularly actinomycetes, have played a dominant role in antimicrobial discovery. Of the estimated 22,500 biologically active microbial compounds, more than 64% of all known antimicrobial classes are derived from the genus Streptomyces (10, 11). Additional contributors of antimicrobials are bacteria from the genera Bacillus, Paenibacillus, and Pseudomonas (11). Although natural products from microorganisms have yielded the majority of clinically used antimicrobials, a vast reservoir of untapped bioactive compounds still exists in microbial genomes.
Previously, the environmental bacterium Pseudomonas protegens PBL3 was identified as an antagonistic bacterium to the plant pathogenic bacterium Burkholderia glumae, the causal agent of the rice disease Bacterial Panicle Blight (12, 13). The antagonistic activity of P. protegens PBL3 against B. glumae was associated with the P. protegens PBL3 secreted fraction (secretome), indicating that P. protegens PBL3 produces and secretes antimicrobials (13). Those antimicrobials are likely associated with at least 14 biosynthetic clusters in the P. protegens PBL3 genome predicted to encode secondary metabolites, such as 2 Pyoverdines, Pyrrolnitrin, Pyoluteorin, 2,4-diacetylphloroglucinol, Orfamide C, Arylpolyene, 2-Hydroxyphenylthiazoline/Thiazostatin/Watasemycin B/Pyochelin, Fengycin, Lipopeptide 8D1-1, 2 Bacteriocins, Cyclodipeptides, and N-acetylglutamine amide (13). Among those, eight were commercially available for testing, but only Pyoverdine, Pyoluteorin, and 2,4-Diacetylphloroglucinol inhibited the growth of B. glumae at high concentrations, yet did not fully explain the antimicrobial activity of P. protegens PBL3 secretome (14). Moreover, other studies demonstrated the antifungal, but not antibacterial activity of Thiazostatin, Orfamide, and Fengycin (15–18); and only Watasemycin B has demonstrated weak antibacterial activity (17).
Although the complete chemical composition of the P. protegens PBL3 secretome is still unknown, the predicted complexity based on its coding capacity led us to hypothesize that the P. protegens PBL3 secretome possesses broad-spectrum antimicrobial activity against other bacterial pathogens. In this study, we investigated the spectrum of activity of the P. protegens PBL3 secretome against plant and human pathogenic bacteria. Our results showed that the secretome inhibited the growth of plant pathogenic bacteria of the genera Burkholderia, as well as selected human pathogens, such as Acinetobacter baumannii, Yersinia enterocolitica, Escherichia coli O157:H7, Listeria innocua, Enterococcus faecium, and Staphylococcus aureus. These findings suggest that the P. protegens PBL3 secretome represents a source of antimicrobials of broad spectrum and possibly novel mechanisms of action for the management of both plant and human bacterial pathogens.
MATERIALS AND METHODS
Bacterial strains
Bacterial strains in this study are listed in Table 1. P. protegens PBL3 was grown on LB agar, plant pathogens were grown on King’s B (KB) agar, and human pathogens were grown on Tryptic Soy agar. Bacterial strains were streaked from the −80°C freezer on agar plates and incubated at 28°C for plant pathogenic bacteria and 37°C for human pathogenic bacteria.
Antimicrobial assay against plant pathogenic bacteria
A single colony of P. protegens PBL3 was grown in M9 Minimal media (1× M9 Minimal Salts [Sigma-Aldrich, St. Louis, MO, U.S.A.] 2 mM MgSO_4_·7H_2_O, 56 mM myo-inositol, and 0.2 mM CaCl_2_) for 36–48 h until the optical density measured at 600 nm (OD_600_) reached 0.5. The secreted fraction (secretome) was separated from whole bacterial cells by centrifugation and filter sterilized as previously described (14).
Plant pathogenic bacterial strains were scraped from the agar plates and resuspended in sterile water to a final concentration of OD_600_ = 0.2. The bacterial suspension was mixed with either P. protegens PBL3 secretome or water (control) and incubated on a shaker for 18 h. Bacterial growth was evaluated by measuring the OD_600_. Each treatment had three replications and was repeated three times.
Antimicrobial assay against human pathogenic bacteria
Human pathogenic bacterial strains were scraped from the agar plates and resuspended in 1× phosphate-buffered saline (PBS) to an OD_600_ of 0.2 (1.5 × 10⁸ colony-forming units [CFU]/mL) and further diluted in a growing medium to a final bacterial concentration of 1.5 × 10⁶ CFU/mL. The antimicrobial activity of the P. protegens PBL3 secretome was assessed in a 96-well microtiter plate. Each well contained a total volume of 100 µL, consisting of growth media, P. protegens PBL3 secretome (or PBS control), and the bacterial suspension. A fixed volume of 10 µL of the bacterial suspension to be evaluated was added to both treatment and control wells. The antimicrobial potency of P. protegens PBL3 was evaluated using a microdilution assay, where varying volumes (10, 20, and 30 µL) of the PBL3 secretome were tested, adjusting the final volume to 100 µL with growth media. Each treatment was conducted in six replicates. For the controls, PBS was added in volumes equivalent to those of the secretome. Growth media alone (100 µL) was used as a negative control, while 10 µL of bacterial suspension in 90 µL of growth media was used as a positive control. Microtiter plates were incubated at 37°C for 18 h. Visual confirmation of bacterial growth was done by adding 0.1% of 2,3,5-triphenyl-tetrazolium chloride (TTC) solution to each well, followed by incubation at 37°C for 30 min to quantify the conversion of colorless TTC into formazan that occurs through the metabolic activities of live bacteria. The formation of formazan was measured by reading absorbance at 470 nm (A470nm) using a plate reader (Synergy 2 Multi-mode microplate reader, Biotek Instruments, Inc.).
Statistical analysis
Data analysis was conducted using R Studio version 4.2.1, calculating the mean and standard deviation of bacterial growth measured for both P. protegens PBL3 treatment and control groups in plant and human pathogens. An unpaired two-way Student’s t-test was performed to determine differences between the control and P. protegens PBL3 secretome, with the significance level set at P = 0.05.
RESULTS
The P. protegens PBL3 secretome has a broad-spectrum activity against plant pathogens
The finding that the P. protegens PBL3 secretome exhibited antimicrobial activity against the rice pathogen B. glumae (13, 14) prompted us to evaluate its activity against several plant pathogenic bacteria, such as other Burkholderia species, as well as Xanthomonas campestris pv. malvacearum, Erwinia amylovora, and Xanthomonas axonopodis. The antimicrobial assays revealed that the P. protegens PBL3 secretome significantly inhibited the growth of all the strains belonging to the genus Burkholderia, but it was ineffective against X. campestris pv malvacearum, E. amylovora, and X. axonopodis (Fig. 1; Table S1). The levels of inhibition among Burkholderia spp. ranged from 31% to 93% (P < 0.05). B. glumae strains grown with the P. protegens PBL3 secretome showed growth reductions ranging from 78% to 93% compared to their growth without the P. protegens PBL3 secretome (P < 0.05). The inhibitory effect of P. protegens PBL3 secretome against Burkholderia gladioli was moderate, with a 46% growth reduction in B. gladioli CU3082 (P < 0.05), a 54% growth reduction in B. gladioli CU3083, and a 64% growth reduction in B. gladioli CU3891 (P < 0.05). Strains of Burkholderia cenocepacia exhibited variable susceptibility against the P. protegens PBL3 secretome, with 85% reduction in B. cenocepacia CU3371-2 (P < 0.05), a reduction of 75% in B. cenocepacia 0318 (P < 0.05), 67% reduction in B. cenocepacia CU6878 (P < 0.05), a reduction of 59% in B. cenocepacia CU3371-1 (P < 0.05) and B. cenocepacia 3368 (P < 0.05), and a minimal reduction of 31% in B. cenocepacia CU3094 (P < 0.05). The Burkholdera spp. O64a showed a growth reduction of 84% compared with the controls (P < 0.05) (Fig. 1). Overall, the P. protegens PBL3 secretome displayed broad yet selective antimicrobial activity, showing strong inhibition of B. glumae strains and variable suppression of other Burkholderia species but no measurable effect on Xanthomonas or Erwinia strains.
The P. protegens PBL3 secretome inhibits the growth of a broad range of plant pathogenic bacteria. Bacterial strains OD600 = 0.2 were grown on King’s B (KB) broth (black bars) or KB amended with the P. protegens PBL3 secretome (gray bars) and incubated at 28°C with constant agitation. Bacterial growth (OD600) was measured after 18 h. Bars represent the average bacterial growth, and the line graph above the bar represents the standard deviation of bacterial growth for three replications. Students’ t-test was used to evaluate statistical significance when comparing bacterial growth in non-amended versus amended media. The asterisk above comparing bars indicates statistically significant differences with a P value = 0.05.
The P. protegens PBL3 secretome demonstrated an inhibitory effect on human pathogens
To evaluate the antimicrobial activity of the P. protegens PBL3 secretome against human bacterial pathogens, we selected seven Gram-negative bacteria, including the human pathogens A. baumannii, E. coli O157:H7, Pseudomonas aeruginosa PAO1, P. aeruginosa PA 103, Salmonella typhi, Y. enterocolitica, as well as the non-pathogenic laboratory strain E. coli K12. We also selected three Gram-positive pathogenic bacteria: E. faecium, L. innocua, and S. aureus. The results showed that the P. protegens PBL3 had differential activity against Gram-negative bacteria, inhibiting the growth of A. baumannii, E. coli O157:H7, and Y. enterocolitica (Fig. 2; Table S2). In A. baumannii, the P. protegens PBL3 secretome at 20% and 30% (vol/vol) caused a 93% reduction in growth (P < 0.05) when compared against the control (Fig. 2; Fig. S1A). Similarly, E. coli O157:H7 showed an average reduction of 40% (P < 0.05) when growth media was amended with P. protegens PBL3 secretome at 20% and 30% (vol/vol) in comparison with growth in non-amended media (Fig. 2; Fig. S1B). Similarly, P. protegens PBL3 secretome caused a 96% reduction in growth (P < 0.05) in Y. enterocolitica when the P. protegens PBL3 were added at 20% and 30% (vol/vol) (Fig. 2; Fig. S1F). The P. protegens PBL3 secretome did not have any effect against the pathogens P. aeruginosa PAO1 and PA13, and S. typhi or the laboratory strain E. coli K12 (Fig. 2).
The P. protegens PBL3 secretome inhibits the growth of a broad range of human pathogens. Pathogenic Gram-negative and Gram-positive bacteria and the laboratory strain E. coli K12 were grown in agar plates for 20 h. Bacteria were scraped from a plate and diluted in PBS buffer to an optical density 600 (OD600) of 0.2 and further diluted in growing media to a final concentration of 1.5 × 106 CFU/mL. Twenty microliters of the P. protegens PBL3 secretome or PBS were mixed with different volumes of growing media to a final volume of 100 µL and grown for 18 h at 37°C with constant agitation. After 18 h, 0.1% 2,3,5-triphenyl-tetrazolium chloride was added to the cultures. Absorbance at 470 nm was read in a plate reader. The bar represents the average absorbance of bacteria minus the absorbance of media alone, and the error bar represents the standard deviation of six replicates. Asterisk suggests a difference between the PBS and PBL3 secretome, with a level of significance set at P = 0.05.
We also found that the P. protegens PBL3 secretome caused a growth reduction in E. faecium, L. innocua, and S. aureus (Fig. 2; Table S2). In E. faecium, the P. protegens PBL3 secretome at 20% and 30% (vol/vol) caused a growth reduction of 25% and 55%, respectively (P < 0.05), when compared with the controls (Fig. 2; Fig. S2A), whereas in L. innocua amended with P. protegens PBL3 secretome, a growth reduction of 33% was only observed at 30% (vol/vol) (P < 0.05) (Fig. S2B). Similarly, S. aureus showed an average reduction of 30% (P < 0.05) when the P. protegens PBL3 secretome was added at 20% and 30% (vol/vol) (Fig. 2; Fig. S2C). Altogether, these results show that the P. protegens PBL3 secretome has a differential level of activity against the Gram-negative pathogens tested, showing higher inhibition of A. baumannii and Y. enterocolitica, and measurable inhibitory effects on E. coli O157:H7, S. aureus, L. innocua, and E. faecium, while having no detectable effect on E. coli K12, P. aeruginosa PAO1, P. aeruginosa PA13, or S. typhi.
DISCUSSION
Our previous findings that P. protegens PBL3 has antagonistic activity against B. glumae and that such activity was associated with the P. protegens PBL3 secretome created an opportunity to harness the P. protegens PBL3 secretome to develop effective biopesticides against B. glumae. In the past, B. glumae infections in rice were treated with kasugamycin and oxolinic acid, but they proved ineffective with the emergence of resistance (20, 21), prompting research into alternative strategies to control this pathogen (22–24).
In this work, we demonstrated that in addition to B. glumae, the P. protegens PBL3 secretome has a broad yet selective spectrum of activity inhibiting the growth of other plant pathogenic Burkholderia strains, including B. gladioli, which also causes BPB, and B. cenocepacia, causing sour skin disease in onion (19). While our study focused on phytopathogenic strains of Burkholderia, it is important to note that other species within this genus are significant human pathogens. The Burkholderia cepacia complex (BCC), which includes at least 24 species, is known to cause opportunistic infections, such as cystic fibrosis-related lung disease, bloodstream infections, and respiratory tract infections (25, 26). Although B. gladioli is not part of the BCC, it has also been isolated from patients with cystic fibrosis and is considered clinically relevant (27). The BCC group is notoriously difficult to treat due to both intrinsic and acquired resistance to multiple classes of antimicrobials, leaving few effective treatment options (25). While we did not test any BCC-associated human pathogens in this study, the demonstrated antimicrobial activity of the P. protegens PBL3 secretome against B. gladioli and other Burkholderia spp. suggests its potential relevance beyond plant disease management and warrants further exploration in the context of human health.
In that regard, we found that the P. protegens PBL3 secretome was also effective against human pathogens, such as A. baumannii, E. coli O157:H7, Y. enterocolitica, S. aureus, and E. faecium. The activity of the P. protegens PBL3 secretome against the human pathogens tested is significant in the context of AMR. A. baumannii, listed by the World Health Organization in 2017 as a top-priority (critical) pathogen for the development of new antimicrobials, is often multidrug-resistant and is associated with mortality rates of up to 60% in hospital-acquired pneumonia and bloodstream infections (28–30). The A. baumannii strain used in our study is an antimicrobial-resistant clinical isolate recently obtained from a patient.
Similarly, the WHO Tier 2 high-priority list includes other clinically significant bacteria, such as E. faecium and S. aureus, which are also increasingly resistant to current antibiotics (30). While our study did not specifically evaluate antibiotic-resistant strains of these pathogens, we observed measurable inhibition of their growth when exposed to the PBL3 secretome. These findings suggest the presence of compounds with activity against a broad range of pathogens and support the potential of this secretome as a source for future antimicrobial discovery.
Our previous results, validating genomic predictions by evaluating the antimicrobial activity against B. glumae of eight purified secondary metabolites: Orfamide A, Orfamide B, Fengycin, Pyrroniltrin, Pyochelin, Pyoverdine, 2,4 DAPG, and Pyoluteroin revealed that only Pyoverdine, 2,4 DAPG, and Pyoluteroin had antimicrobial activity at high concentrations. However, those concentrations were not equivalent to their respective endogenous concentrations in the P. protegens PBL3 secretome, implying that the overall antimicrobial activity of the P. protegens PBL3 secretome is associated with novel molecules or a combination of molecules (14).
Although the specific molecules associated with the antimicrobial activity in the P. protegens PBL3 secretome remain to be identified, some of the P. protegens PBL3 genome-predicted secondary metabolites have been previously studied. The exposure of bacterial cells to 2,4-DAPG leads to cell membrane damage and cell content disintegration (31, 32). Pyoverdine is a high-affinity siderophore whose antimicrobial properties are associated with ferric iron (Fe³^+^) sequestration that deprives pathogen cells of iron (33, 34). Pyoluteorin damages the bacterial cell membrane, causing cytoplasmic leakage, as well as inducing the accumulation of reactive oxygen species, leading to lethal oxidative damage (35). These multiple modes of action associated with molecules expected to be in the P. protegens PBL3 secretome suggest that the P. protegens PBL3 secretome could have a multifaceted effect, simultaneously targeting several bacterial processes.
Despite the broad spectrum of activity of the P. protegens PBL3 secretome, our results also showed differential activity among strains, suggesting that bioactive molecule(s) in the P. protegens PBL3 secretome have unique target(s) that are present in P. protegens PBL3-sensitive bacteria and absent in the P. protegens PBL3-insensitive bacteria. Hypothetically, bacteria that are insensitive to the P. protegens PBL3 secretome employ mechanisms known to drive AMR, such as detoxification, efflux pumps, or specific adaptive strategies to prevent internalization of bioactive molecules (36). Our results also revealed that P. aeruginosa and S. typhi were insensitive to the P. protegens PBL3 secretome. P. aeruginosa has intrinsic resistance to antimicrobials through several mechanisms, including (i) unique outer membrane composition that prevents antibiotic uptake, (ii) downregulation of membrane porins required for antimicrobial uptake, (iii) presence of multiple efflux pump systems which actively expel a wide range of structurally unrelated antibiotics, (iv) constitutive production of antibiotic hydrolyzing enzymes, and (v) biofilm formation that provides a protective shield restricting antibiotic penetration and promotes nutrient and oxygen gradients (37, 38). The resistance mechanisms of S. typhi against frontline antibiotics, particularly fluoroquinolones and third-generation cephalosporins, are complex and often plasmid-mediated. S. typhi exhibits antibiotic resistance through multiple mechanisms, including (i) chromosomal mutations which alter the drug target, (ii) plasmid-mediated mechanisms that protect target enzymes and decrease intracellular drug accumulation, (iii) production of antibiotic hydrolyzing enzymes, (iv) overexpression of efflux pumps that actively expel antibiotics from the cell, and (v) decreased outer membrane permeability resulting from porin modifications (39, 40).
The broad-spectrum antimicrobial profile of the P. protegens PBL3 secretome is significant, given that the affected pathogens are usually sensitive to antibiotics representing multiple mechanistic classes. For instance, β-lactam antibiotics, which act by binding to penicillin-binding proteins involved in peptidoglycan biosynthesis, are commonly used to treat infections caused by A. baumannii, S. aureus, and L. innocua (28, 41). Likewise, Y. enterocolitica is susceptible to aminoglycosides, which exert their effects by irreversibly binding to the 30S ribosomal subunit and inhibiting protein synthesis, while fluoroquinolones inhibit bacterial DNA gyrase and topoisomerase IV, preventing DNA replication, used for the treatment of S. typhi infections (39). The inhibition of multiple bacterial genera by the P. protegens PBL3 secretome, each typically sensitive to distinct classes of antibiotics, highlights its potential to contain a combination of antimicrobial compounds that act through different or overlapping molecular targets.
In clinical settings, numerous antibiotic classes are available for the treatment of human bacterial infections; however, in agriculture, only three antibiotics, streptomycin, kasugamycin, and oxytetracycline, are currently registered in the United States for the management of bacterial plant diseases (7). Consequently, the broad-spectrum antimicrobial activity exhibited by the P. protegens PBL3 secretome against multiple plant pathogens highlights its potential as a novel biological alternative for sustainable disease management. This is particularly relevant in agricultural systems where the limited availability of effective antimicrobial agents, coupled with the growing concern over resistance development, highlights the urgent need for safe and environmentally compatible biocontrol solutions.
Overall, our study shows that the P. protegens PBL3 secretome contains bioactive molecules with activity against diverse plant and human bacterial pathogens that could be used as a source of antimicrobial compounds for the development of novel therapeutics. Although the specific compounds remain to be identified, ongoing work aims to identify and validate them.
Supplementary Material
Reviewer comments
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Prescott JF. 2017. History and current use of antimicrobial drugs in veterinary medicine. Microbiol Spectr 5. doi:10.1128/microbiolspec.arba-0002-2017 PMC 1168755329271338 · doi ↗ · pubmed ↗
- 2Schar D, Klein EY, Laxminarayan R, Gilbert M, Van Boeckel TP. 2020. Global trends in antimicrobial use in aquaculture. Sci Rep 10:21878. doi:10.1038/s 41598-020-78849-333318576 PMC 7736322 · doi ↗ · pubmed ↗
- 3Van Boeckel TP, Glennon EE, Chen D, Gilbert M, Robinson TP, Grenfell BT, Levin SA, Bonhoeffer S, Laxminarayan R. 2017. Reducing antimicrobial use in food animals. Science 357:1350–1352. doi:10.1126/science.aao 149528963240 PMC 6510296 · doi ↗ · pubmed ↗
- 4Velazquez-Meza ME, Galarde-López M, Carrillo-Quiróz B, Alpuche-Aranda CM. 2022. Antimicrobial resistance: one health approach. Vet World 15:743–749. doi:10.14202/vetworld.2022.743-74935497962 PMC 9047147 · doi ↗ · pubmed ↗
- 5Miethke M, Pieroni M, Weber T, Brönstrup M, Hammann P, Halby L, Arimondo PB, Glaser P, Aigle B, Bode HB, et al.. 2021. Towards the sustainable discovery and development of new antibiotics. Nat Rev Chem 5:726–749. doi:10.1038/s 41570-021-00313-1PMC 837442534426795 · doi ↗ · pubmed ↗
- 6WHO. 2023. Antimicrobial resistance. World Health Organization. Available from: https://www.who.int/news-room/fact-sheets/detail/antimicrobial-resistance. Retrieved 21 Jul 2025.
- 7Sundin GW, Wang N. 2018. Antibiotic resistance in plant-pathogenic bacteria. Annu Rev Phytopathol 56:161–180. doi:10.1146/annurev-phyto-080417-04594629856934 · doi ↗ · pubmed ↗
- 8Verhaegen M, Bergot T, Liebana E, Stancanelli G, Streissl F, Mingeot-Leclercq MP, Mahillon J, Bragard C. 2023. On the use of antibiotics to control plant pathogenic bacteria: a genetic and genomic perspective. Front Microbiol 14:1221478. doi:10.3389/fmicb.2023.122147837440885 PMC 10333595 · doi ↗ · pubmed ↗