Different MicroRNAs expression in Mycobacterium tuberculosis and correlation with prognosis of the disease
Karthikeyan Sundaram, Sridhar Rathinam

TL;DR
This paper reviews how different microRNAs are expressed in tuberculosis and how they may be linked to disease prognosis.
Contribution
The paper highlights specific microRNAs associated with tuberculosis progression and drug resistance.
Findings
miRNA-146 is more highly expressed in TB patients' serum exosomes compared to healthy individuals.
Drug-resistant tuberculosis is linked to miR-548m, miR-631, and other microRNAs in lesion tissue and blood.
miR-155, miR-29, circ-miRNA, and lncRNAs regulate gene expression in TB prognosis.
Abstract
Tuberculosis, caused by Mycobacterium tuberculosis, is an infectious disease linked to high mortality and can stay in the host cell longer when inactive. Multiple factors are linked to disease prognosis, including microRNAs. It is a diminutive single-stranded RNA that regulates the expression of its target mRNAs. It consists of a brief nucleotide sequence, often 19–25 nucleotides in length, of non-coding RNA. It is also essential for early embryonic development, invasion, cell migration, apoptosis, and cell death. The review aims to analyse the transcriptome characteristics of various miRNAs in the tuberculosis prognosis. However, miR-155, miR-29, circ-miRNA, and lncRNAs regulate gene expression. In TB patients’ serum exosomes, miRNA-146 expression was noticeably higher than in healthy individuals. Drug-resistant tuberculosis was related to miR-548 m, miR-631, miR-328-3p, and…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
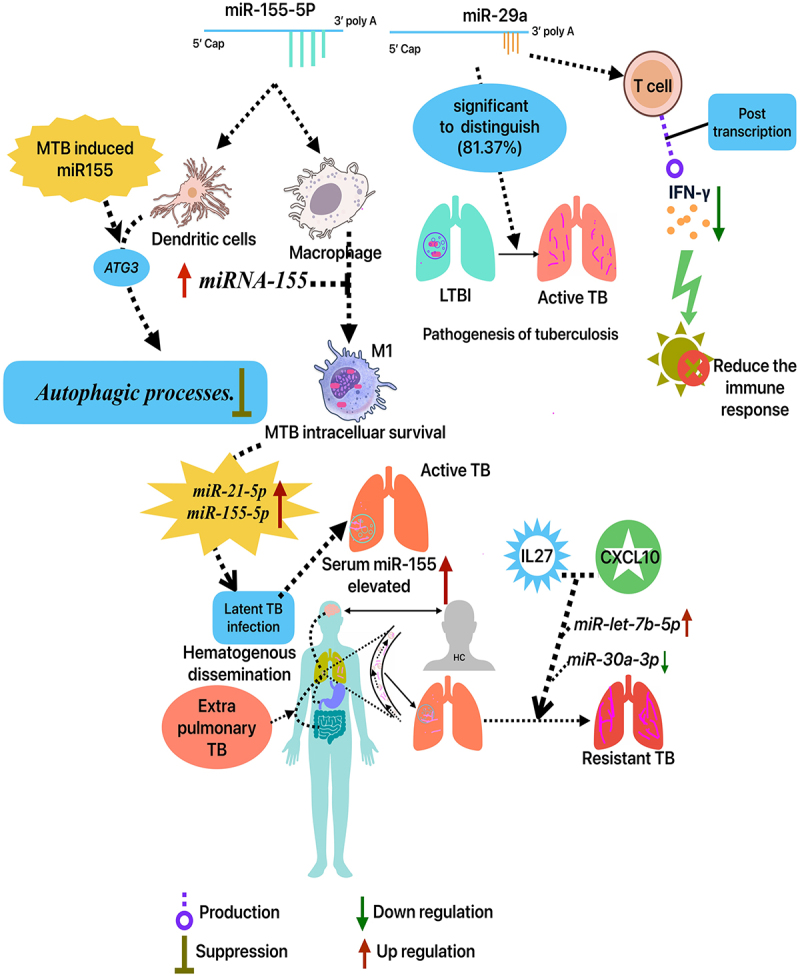 Figure 1
Figure 1 Figure 2
Figure 2Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMicroRNA in disease regulation · Tuberculosis Research and Epidemiology · Infectious Diseases and Tuberculosis
Introduction
Tuberculosis (TB) is an infectious disease caused by Mycobacterium tuberculosis (MTB). MTB can stay in the host cell longer while it is dormant, even though it is a preventable and treatable infection. The WHO Global Tuberculosis Report 2024 revealed that 10.8 million individuals (95% UI: 10.1–11.7 million) became ill with TB globally in 2023 [1]. The progression of TB from latent to the active state depends on the host’s primary defence mechanism. Although, the MTB gene’s function is capable of adapting within the alveolar macrophage, and enhances their self-defence mechanism against autophagosome-lysosome fusion action [2]. Differently expressed (DE) circRNA, miRNA, and mRNA control transcription and immunological responses in TB. They usually affect transcription, protein binding, and immunological responses positively and negatively. A recent study identified a core ceRNA regulatory network with five circRNAs (hsa-circ_0001844, hsa-circ_0000566, hsa-circ_0007587, hsa-circ_0086710, and hsa-circ_0005408) and one miRNA (hsa-miR-607) that compete to regulate IFNG, three mRNAs, including GBP2, impact IFNG simultaneously. More research is needed to address microbe-host interaction gaps. Database for Annotation, Visualization, and Integrated Discovery (DAVID) analyzes DE genes. CircRNAs are ribonuclease and exonuclease-resistant due to their covalently closed-ring structure and lack of a 5′ terminal cap and 3′ terminal poly(A) tail. CircRNAs’ long half-life and stability make them promising biomarkers [3–5]. Notably, miRNAs affect a number of important processes, such as apoptosis, differentiation, and cell proliferation. Most significantly, abnormal patterns of miRNA expression have been linked to the pathophysiology of numerous diseases. These patterns control the generation of numerous cytokines and chemokines that modify the host immune response throughout different disease states. Importantly, transcriptome patterns in whole blood can detect TB, infection, and illness using biomarkers including IFN-γ, IP-10, LAM, and IL-2 [6]. Also, the limitation behind the conventional diagnosis of TB including insensitivity of microscopy and the time required for culture make regular TB testing challenging. Molecular accelerators like GeneXpert are expensive and uncommon in rural regions. The molecular testing or culture of smear-negative pulmonary and extra-pulmonary TB needs invasive techniques. Therefore, this review aims to examine the different types of miRNAs, and other regulatory genes involved along with mRNA to the expression of genes in modulating the MTB survival as dormant in macrophages as well as progressing their ability to the active stage.
Role of miRNAs
MiRNAs, single-stranded, non-coding RNA with a 19–25 nucleotide sequence, govern the expression of their target mRNAs. They are essential for invasion, cell migration, apoptosis, cell death, and early embryonic development. Also, persistent miRNAs are discovered in sputum, and several diseases are significantly linked to their differential expression. In severe asthma, sputum miR-629-3p, miR-223-3p, and miR-142-3p levels are higher, indicating neutrophilic airway inflammation. Sputum miRNAs contribute to asthma’s inflammatory phenotype. NSCLC sputum had higher miR-223 levels than healthy controls [7–9].
Role of miRNA-155 in TB pathogenesis
2.1.
The miRNA-155 is a master regulator for numerous diseases particularly this miR-155 is linked to pathogenesis of TB infection. Through the regulation of Ras Homologue Enriched in Brain (RHEB) gene expression, it was found that miRNA-155 may promote cellular autophagy and facilitate the elimination of MTB from the host. While Akt, AMP-activated protein kinase, and glycogen synthase kinase-3β affect RHEB activity, the mechanisms governing RHEB expression modulation are not well understood. Autophagy results from the post-transcriptional downregulation of RHEB expression by miR-155. Furthermore, study demonstrated that miRNA-155 regulates the production of BCG-induced reactive oxygen species via influencing SHIP-1. Research indicates that MTB-infected RAW264.7 cells had significantly increased miRNA-155 expression, which was closely associated with MTB intracellular survival. However, the miR-155 gene promotes iNOS production and classical activation (M1 polarization) in alternatively activated (M2) macrophages by targeting CEBPβ. The study demonstrated that miR-155 stimulates LC3 processing and puncta formation in BCG-challenged and unchallenged RAW264.7 cells. Macrophage autophagy and intracellular microorganism reduction are caused by miR-155. Moreover, Atg7 knockdown or 3-MA autophagy suppression lowered miR-155-mediated autophagy and bacterial mortality. 3-MA also significantly boosted BCG survival after 6 h in control mimic-treated RAW264.7 cells, despite low LC3-II decrease. In addition, 3-MA inhibits nitric oxide generation, boosting mycobacteria survival. Targeting ATG3 in human dendritic cells allows MTB-induced miRNA-155 to suppress autophagic processes. Serum miRNA-155 concentrations in patients with active tuberculosis were markedly elevated compared to those in healthy individuals, suggesting that miRNA-155 serves as a biomarker for the diagnosis of active tuberculosis. These results illustrate that miRNA-155 is a crucial regulator of tuberculosis infection by altering host gene expression, either directly or indirectly [10–13]. The human macrophage cell line THP-1, when exposed to the MTB strain H37Rv, exhibited an elevation of miR-155. In addition to macrophages, CD14+ cells or peripheral blood mononuclear cells (PBMC) isolated from persons with active tuberculosis exhibited an upregulation of miR-155. Concerning the mechanism of miR-155 immunomodulatory action, previous studies indicated that MTB may target the ATG3 gene to alter miR-155 production, thereby inhibiting autophagy in human dendritic cells for its survival (Figure 1) [7,14]. Importantly, the role of miRNAs-155 is crucial in MTB infection, since one important modulator of innate immunity is miR-155-5p. Early on in the infection process, innate immunity is suppressed due to the increased expression of miR-155-5p caused by the MTB infection. In this context, the pure protein derivative 10 stimulated PBMCs from active TB patients, increasing the production of miR-155 and miR-155×. Thus, the miR-155 was thoroughly examined. Though the innate immunological responses brought on by MTB infection and the macrophage apoptosis brought on by Bacillus Calmette–Guérin (BCG) infection were both suppressed by ESAT6-dependent miR-155. However, miR-155 targets immune-related proteins like C/EBP and SHIP1, which negatively regulate the IL-6 signalling pathway, along with SOCS1. ESAT-6 may control SHIP-1, C/EBP β, or both via miR-155 to affect immunological response [15,16]. Figure 1.Different miRNAs associated with the progression of TB.
Role of miR-29 in TB pathogenesis
2.2.
The recent meta-analysis study observed that miR-29a is effective in discriminating latent pulmonary TB from active TB (AUC = 84.35%) and recognizing active TB from healthy patients (AUC = 81.37%). Blood-derived miR-29a-3p can identify active TB patients from healthy controls, suggesting it could be a useful biomarker. This miRNA was found in plasma samples, unlike sputum, suggesting its potential use in diagnosing TB, especially in children and extra-pulmonary cases. Also, the results supported earlier opinions. miR-29a has 82% sensitivity and 81% specificity for adult tuberculosis diagnosis and 74% and 89% for kid diagnosis [17,18]. Also, multiple studies have shown that the gene expression patterns of macrophages and NK cells differ among active TB patients, latent patients, and healthy controls (HC). In systemic infections produced by Mycobacterium bovis or Listeria monocytogenes, NK cells downregulate miR-29, an mRNA repressor in viral infections targeting the HIV-1 3′UTR region, reducing IFN- γ. The host’s Listeria resistance is improved by miR-29a. Compared to HCs, active pulmonary TB patients had higher serum and sputum miR-29a levels. Mycobacterium avium-infected human macrophages had increased miR-29a and miR-let-7e, which targeted caspases 7 and 3. Since Mycobacteria infections reduce apoptosis, miRNAs regulate it. MiRNAs regulate gene expression and cellular composition in TB patients. Many miRNAs regulate T cell development and function. It is also known that miRNAs regulate dendritic, NK, macrophage, and innate immune cell function [19]. Hence, patients with active and latent TB had markedly elevated levels of blood miR-29a-3p in comparison to healthy controls. Consequently, miR-29a-3p along with miR-155 play a significant role in prognosis of the disease, might serve as a possible biomarker for pulmonary TB diagnosis (Table 1) [20].Table 1.Functional analysis of miRNA-155 and miR29 linked to tuberculosis.StudyTypes of miRNAsRoleClinical outcomesTechnical aspects: in-vivo/in-vitroZheng, M. L et al., 2016miR-155controlling the anti-mycobacterial response through autophagygood performance between PTB and control groups.Between PTB and control groups, miR-155 with high AUC of 0.976. miR-449a, miR-212, and miR-132 with AUC values of 0.947, 0.931, and 0.930.Ying H et al., 2020miR-155controlling the anti-mycobacterial response through autophagyThe sensitivity and specificity of miR-155 identification were 94.1% and 87.7%.68 PTB and 122 non-PTB patients.Wang, L. H et al., 2025miR-155controlling the anti-mycobacterial response through autophagyThe serum miR-155 AUC-0.968, threshold of 1.50. 91.80% sensitivity. 89.00% specificity. sputum miR-155 AUC for TB diagnosis was 0.977 with a threshold of 1.70, resulting in 96.40% sensitivity and 90.0% specificity.PTB-110 cases, and HC-50. In this active TB-50/latent TB-50.miR-146aAffecting expression of TNF receptor associated factor-6 (TRAF-6)High AUC predicts better TB diagnosis miR-146a/155. Serum miR-146a‘s AUC −0.925 cut-off of 1.25, leading in 85.50%-sensitivity 87.00%-specificity. sputum miR-146a‘s AUC-0.940, threshold of 1.35, 80.90% sensitivity. 84.55% specificity.Ying, H et al., 2020miR-155controlling the anti-mycobacterial response through autophagyThe miR-155 detection sensitivity-94.1%. specificity-87.7%. It exceeded anti-TB antibodies and sputum smears.68 PTB patients and 122 non-PTBNdzi, E. N et al., 2019hsa-miR-29a-3p**miRNA-29a-3p reduces immunological response by downregulating IFN-γ production in T cells post-transcriptionally.miR-29a-3p was good in distinguishing ATB from LTB and HEC (81.37% and 84.35%, respectively).A total of 162 cases, active TB-84, latent-35, HC-43Angria N et al., 2022hsa-miR-29a-3p**miR-29a-3p attenuates the immune response by post-transcriptionally downregulating interferon (IFN)-γ production in T cells.For latent TB, the miR-29a-3p ROC curve – AUC of 0.808 (95% CI: 0.698–0.919), 70% specificity, and 84.8% sensitivity. Neither in patients with active or latent pulmonary TB (R = 0.005; p = 0.62) nor in healthy individuals (R = 0.060; p = 0.19), did the expression of miR-29a-3p show any association with INF-γ levels.50 active TB, 33 household contacts with positive IFN-γ release, and 30 healthy controls were included in this study.
Circulatory miRNAs roles in diagnosis of TB
2.3.
Circulating miRNA influences gene expression and immunological and non-immune cell activity. Endogenous regulators include many miRNAs. For several diseases and pathologies, stable circulating miRNAs have been reported, miR-let-7a, −221, −26a, −191, −320a, and -93-5p. The most often utilized reference genes for normalizing circulating miRNA expression are RNU6B and miR-16. After geometric ranking analysis, miR-22-3p was the most stable at 1.19 and 1.73, followed by miR-93-5p. miR-16-5p was least steady at 2.83. Total stability: let-7i-5p > let-7a-5p > miR-16-5p > miR-22-3p > miR-93-5p. Thus, host, environmental, or both variables may affect circulating miRNA stability and gene expression [21]. Also, comparing the circulatory-miRNA (c-miRNA) signature to the RNA-CoR (RNA based Correlate of Risk) signature may help c-miRNAs regulate the interferon response to TB. RNA-CoR genes showed a strong correlation with mycobacteria-induced miR-21, a marker of immune cell activation. RNA-CoR genes, including FCGR1B, were negatively associated with miR-26a, which suppresses the macrophage response to IFN-γ, and miR-30b, which suppresses the expression of Fc receptors and the generation of pro-inflammatory cytokines [22]. However, c-miRNA profiles have associated miR-21 with exosomes, which are small vesicles derived from cells that transport stable short RNAs, including miRNA, suggesting its role as a secreted biomarker of disease. Exosomes serve as conventional immunomodulators by facilitating cell-to-cell communication. The presence of miR-21 in various exosomes, including those derived from tumours and immune cells, indicates its important role in the regulation of immunological functions [23]. Most notably, persistent bacterial or viral infections increase the expression of miR-21 in immune-compromised conditions include asthma, psoriasis, cancer, and the development of regulatory immune cells such as Th2, regulatory T-cells, or M2 macrophages, as well as the reprogramming of pro-inflammatory Th1-cells and M1 macrophages, are associated with a number of these disorders. Therefore, damage-associated molecular pattern molecules (DAMPs) or pathogen-associated molecular pattern molecules (PAMPs) dysregulating miR-21 may create an immunosuppressive, anti-inflammatory milieu that causes illness [24]. Besides that, mycobacterial species survive and thrive in macrophages and create miR-21 via interfering with host responses. Also, miR-21 activates inflammatory mediators in non-haematopoietic cells, leading to neoplastic transformation [25].
Hierarchical miRs were found in control and childhood TB groups. Peripheral white blood cells were clustered utilizing 29 miRs expressed differentially in TB children and healthy controls (HC). It is intriguing that 14 candidate miRNAs were chosen for validation from 25 tuberculosis-affected children and 21 HCs. Compared to HCs, TB patients exhibited increased miR-29 but decreased miR-155, miR-31, miR-1, miR-146a, miR-125b, miR-150, and miR-10a. Thus, any of them might be a potential initial diagnostic biomarker for diagnosis of TB in children [26]. Furthermore, recent research found that exosomal miR-130b-3p and miR-423-5p, which boost p65 and Cyclin D1 expression, promote lung cancer (LC) development. The study found high levels of these miRNAs in pleural effusion exosomes but not blood plasma [27,28].
Different miRNAs association with drug-resistant TB
2.4.
The miRNAs are associated with various function in TB significantly, meanwhile drug-resistant TB needs to be studied. However, a recent study found the substantial results that miRNAs roles in drug-resistant (DR) and drug-susceptible (DS) – TB compared to HC. DR-TB alone showed a substantial decrease in miR-548 m and miR-631, two of the down-regulated miRNAs. Recent study analysed that DR-TB showed a substantial decrease in miR-548 m compared to DS-TB, LTB, and HC (fold change: −1.62, p = 0.04, −2.11, p < 0.05, −1.94 for HC). The miR-631 decrease was statistically significant compared to DS-TB (FC: −1.64, p = 0.01) and HC (FC: −1.62, p = 0.008) but not LTB [29]. Additionally, miRNA assessment for DR-TB treatment monitoring: rifampicin-resistant (RR), multi-drug resistant (MDR), or extensively drug resistant (XDR) strains may have formed if miR-let-7e-5p is down-regulated and miR-328-3p is increased at sputum conversion T (3–5). The study also found that miR-20a-3p up-regulation is connected to MDR resistance. Alongside other clinical and microbiological data, the expression of miR-let-7e-5p and miR-328-3p may facilitate the early monitoring of DR-TB patients after therapy [30]. The miRNA signature model for DR-TB (miR-let-7e-5p, miR-223-3p, and miR197-3p) demonstrated 100% diagnostic sensitivity and 75% specificity [31]. Similarly, evidence suggests a relationship between miRNAs and DR-TB, as patients with MDR-TB, DS-TB, and healthy controls showed significant variations in miR-4433b-5p, CD44, miR-424-5p, F11, miR-199b-5p, and KNG1; p < Blood coagulation requires KNG1 and human platelet miRNA miR-199b-5p [32].
Various types of miRNAs are involved in pathogenesis of TB
2.5.
Recent studies have found that differently expressed genes (Table 2) were associated with up and down regulation miRNAs are links to pathogenesis of TB. Particularly, the PTB patients exhibited considerably lower miR-454-3p, miR-15a-5p, miR-590-5p, miR-381, and miR-449a expressions than healthy controls. EPTB patients’ miRNA expression was unaltered. Study also evaluates MTC genotype miRNA expression. miR-590b-5p, which was upregulated in zoonotic strains and LAM7TUR, was the sole miRNA with pretreatment and post treatment changes. A miRNA signature investigation of active TB patients discovered that miR-590-5p was considerably different from healthy patients. However, miR-590-5p plays a clinically crucial in individuals infected with TB. In contrast, the LAM7TUR and Beijing strains had higher miR-15b-5p and miR-21-5p levels than the control and pretreatment patient groups [33]. This study also found that untreated TB patients infected with Beijing strains had considerably higher miR-21-5p expression levels than the control group, contradicting to a recent study that reported decreased expression in cured ones due to anti-TB medications may lower miR-21-5p by downregulating host defence. Due to improved inflammation and antimicrobial status, untreated TB patients should express higher miR-21-5p. The immune system and inflammation may be compromised by anti-TB medications that suppress miR-21-5p expression [33–35]. Besides that, in a preliminary study observed that MTB infection and pulmonary TB pathogenesis depend on miRNA-146a. Numerous studies have shown that the C allele in miRNA-146a’s rs2910164 may decrease mature miRNA-146a synthesis, altering the inflammatory process. Significantly, miR-146a has lower expression in TB than control group, especially Males with TB exhibited higher levels of miR-146a expression (median 0.46; range 0.01–11.59) compared to females (median 0.24; range 0.01–1.3), while the difference was not statistically significant (p = 0.061). The miRNA-146ars2910164 C > G single-nucleotide polymorphism (SNP) may influence the immune response to MTB infection, which could increase the risk of pulmonary tuberculosis. Systemic down-regulation of miRNA-146a, a negative regulator of the immune response, may exacerbate TB patients’ inflammatory response. Unlike the mature miRNA-146a sequence, SNP rs2910164 C > G, encoded on chromosome 5q33, is present in the precursor stem region, + 60 relatives to the first nucleotide [36,37].Table 2.Significant outcomes of various miRNAs in diagnosis of TB.StudyTypes of miRNAsClinical outcomesTechnical aspects: in-vivo/in-vitroKorma W et al., 2020miR-16-5p, −22-3p, −93-5pBestkeeper models for miRNA expression data assessment show acceptable standard deviations (below 1) for miR-93-5p, miR-22-3p, and miR-16-5p. miR-22-3p and miR-16-5p exhibited 0.90 and 0.94 standard deviations, whereas miR-93-5p was the most stable at 0.82.68 trial participants (26 HC, 23 LTBI, and 19 TB) were included in this studyZhou M et al., 2016miR-146a, miR-10a, miR-125b, miR-150, miR-29, miR-1, miR-155, −31Eight miR’s demonstrated excellent diagnostic value with AUCs of 0.996 (95% CI, 0.914–1.0). Both sensitivity and specificity are 95.8% and 100%. miR-1, miR-155, miR-31, miR-146a, miR-10a, miR-125b, miR-150, and miR-29 suggested as early diagnostic biomarker.Total of 129 childrens were enrolled. 28 TB and 24 HCAshirbekov Y et al., 2024miR-376a-3p, miR-15b-5pCompared to controls, TB patients had much greater and lower miR-376a-3p and miR-15b-5p levels.38 pulmonary TB patients, 41 healthy controls, and 68 LC patientsKang, H J et al., 2024Tuberculous exosomes, miR-130b-3p and miR-423-5pPCR sequencing in cell studies and nude mice trials showed that tuberculous exosome-treated A549 cells had much greater miR-130b-3p and miR-423-5p levels.Male BALB/c mice, 12 mice got a subcutaneous transplant of 2 × 10^6^ A549 cells in their flanks.Buha I et al., 2022miRNA-146aCompared to controls, TB patients showed significantly lower miRNA-146a expression (median 0.30, 0.01–1.72). p < 0.001 vs. median 1.000 (0.06–11.59).TB patients − 44, HC-17.Sun X et al., 2022miR-378ATB patients express more miR-378 than LTBI ones. serum miR-378 > 1.490 was sensitive and specific for TB diagnosis. In TB patients, IFN-γ, TNF-α, and IL-12 levels were low, whereas IL-4, IL-6, and IL-1β levels were increased. In the active group, serum miR-378 levels were favourably connected with IL-4, IL-6, and IL-1β, while IFN-γ, TNF-α, and IL-12 concentrations were adversely correlatedA total of 126 TB patients (63 active and 63 latent) and 62 HC were selected.Liu J et al., 2023let-7d-5p, miR-140-5pPanel ROCs for biomarkers. (a) Training ROC curve AUC was 0.93; (b) Test was 0.923. LOOCV’s ROC results showed that our let-7d-5p and miR-140-5p-trained model could differentiate LTBI from ATB or HC with an AUC > 0.9.GSE25435 found 174 DEMs between ATB and LTBI, including 39 downregulated and 51 upregulated miR’s. The GSE29190 dataset comprised 24 DEMsSampath P et al., 2023miR-548 m, miR-631DR-TB alone decreased miR-548 m and miR-631 more than any other group. The decrease in miR-548 m in DR-TB was significantly different from DS-TB, LTB, and HC (FC: −1.94, p < 0.05 vs. FC: −1.62, p = 0.04). However, miR-631 declined significantly compared to DS-TB (FC: −1.64, p = 0.01), HC (FC: −1.62, p = 0.008), but not LTB.(DS-TB) (n = 6), latent TB (LTB) (n = 6), and drug-resistant TB (n = 6), and HC-6 were included in this study.Carranza C et al., 2021let-7e-5p, −197-3p, and −223-3pThe DR-TB (AUC = 0.96, 95%, CI = 0.907–1) and MDR-TB (AUC = 0.95, 95%, CI = 0.89–1) groups showed greater sensitivity in separating HC from those with TB when miR-let-7e-5p, −197-3p, and -223-3p were used.Extracellular Vesicles were recovered from serum of 26 DR-TB patients and 16 HC.Cui X et al., 2025exo-miR-766-3pExosomal miR-766–3p, miR-376c-3p, miR-1283, and miR-125a-5p had AUCs of 0.8963, 0.8313, 0.8125, and 0.8050, respectively. Notably, Plasma-derived exo-miR-766-3p lowered NRAMP1 expression in A549 cells, while miR-766-3p inhibition enhanced it.The study included 32 TB patients and 32 HC.
A recent study analysed serum samples from patients with active and latent tuberculosis, revealing elevated levels of miR-378 compared to healthy individuals, with the active group exhibiting the highest expression levels. In TB patients, miR-378 was always raised. Also, this study found that miR-378 had an AUC of 0.767, a cut-off value of 2.130, 92.060% sensitivity, and 52.380% specificity for distinguishing active and latent groups. Thus, this study suggests miR-378 may be a TB biomarker [38]. On the other hand, the roles of let-7d-5p and miR-140-5p in the pathophysiology of TB remain unclear, and a recent study found that their expression was down-regulated in patient serum when compared to healthy controls. In this context, the biological importance of their target genes was concentrated in kinase activity regulation, vascular development, and growth factor response. However, miRTarBase and miRDB generated 86,781 paired miRNA-target gene interactions with 2459 miRNAs and 8849 target genes. By blocking translation in target genes’ 3’ untranslated regions (3’UTRs), microRNAs downregulate them. After eliminating miRNAs that upregulate mRNAs, ATB and LTBI had 2156 target genes regulated by 89 miRNAs while LTBI and HC had 2139 by 81 miRNAs. Training set AUC for the biomarker panel was 0.930 (sensitivity = 100%, specificity = 88.5%) and test set AUC was 0.923 (sensitivity = 100%, specificity = 92.3%). The receiver operating characteristics (ROC) of LOOCV data showed that let-7d-5p and miR-140-5p-trained models could distinguish LTBI from ATB or HC with an AUC greater than 0.9. Thus, let-7d-5p and miR-140-5p are essential TB indicators. In addition, Dong et al. found that miR-140-5p reduced lung fibrosis via blocking the Wnt1/β-catenin pathway [39,40].
Recent study on validation of Solexa sequencing research found significant differences in 181 serum miRNAs between 10 TB patients and 10 healthy controls (fold change > 2, p< 0.05). A total of 88 miRNAs were down-regulated (<0.50-fold, p < 0.05) and 93 were up-regulated (>2.0-fold, p < 0.05). Solexa found 19 autophagosome maturation miRNAs with above 10 copies for qRT-PCR validation. Study also investigated 11 miRNAs with fold changes (TB patients/HCs) ≥1.2 or ≤0.8 and p < 0.05 from the training set. Sixty TB patients and 41 healthy controls had 10 miRNAs. Patients with TB showed substantially higher levels of miR-17-5p, −20b-5p, −378a-3p, and -423-5p (p < 0.001). In addition to TB patients’ serum, PBMCs the primary source of macrophages that remove MTB, also showed increased miR-423-5p [41]. Also, in the preliminary screening study examined miR-181b-5 connection. Using qRT-PCR, plasma from eight HC and eight TB patients was quantified for hsa-miR-181b-5p. In TB patients’ plasma samples, miR-181b-5p expression increased significantly. Also, in monocytes from whole blood samples, infection with MTB with RAW 264.7 and BMDM in mouse model entails the regulation of SAMD9L expression by TLR2 and HIF-1. The analysis revealed that miR-181b-5p and SAMD9L do not interact, despite being a valuable TB marker [42]. Also, multiple studies have shown that in mycobacteria-infected macrophages, miR-17-5p acts as an immunomodulatory regulator of autophagy. Hsa-miR-17-5p regulates autophagy in MTB-infected macrophages via targeting Mcl-1 and STAT3 [43]. The preferred pathways were systematically categorised and organised. The inhibition of FOXO1 has been demonstrated to inhibit apoptosis and the inflammatory response. Interleukin-6 demonstrates pro-inflammatory characteristics. The suppression of the Egr1/TGF-β/Smad pathway by miRNA-181a-5p is associated with the regulation of cell growth [44–46].
Various differentially expressed genes (DEGs) play a vital role in disease pathogenesis. Likely, abnormal levels of miR-191, miR-146a, miR-20, miR-22, and miR-320 are among the patterns of miRNA expression in body fluids that are linked to the severity of the disease and may be signs of active tuberculosis rather than latent infection [47]. The miR-15/16 family member miR-15a-5p modulates signalling pathways involved in apoptosis, cell cycle regulation, and inflammatory cytokines. Also, miR-15a-5p regulates pro- and anti-inflammatory responses by targeting genes such BCL2, IKKα, and NF-κB signalling cascade components. miR-15a-5p can regulate inflammation and macrophage-mediated bacterial clearance, making it a potential TB biomarker and treatment target [48–51]. Notably, silencing endogenous miRNA-31 can inhibit PD-1/PD-L1 expression on lung CD4+/CD8+T cells, increase MTB burdens, and worsen lung pathology, suggesting a novel TB mechanism [52]. Notably, targeting the 3′ UTR of the TNF mRNA, the miR-125b was up-regulated in MTB-stimulated macrophages, reducing TNF production. Additionally, another study indicated that following TB infection, miR-125b-5p is significantly expressed in a variety of immune cells, mostly monocytes-macrophages. In particular, miR-125b decreases inflammatory responses via TNF-α [24,53,54].
A recent study used qPCR to assess the plasma biomarker potential of 10 miRNAs and 9 proteins. The AUC for all 19 analytes upon diagnosis was 1.000, indicating 100% sensitivity and 100% specificity in TB patients. The panel was reduced to five miRNAs and proteins (IP-10, miR-29a, miR-146a, miR-99b, and miR-221), yielding an AUC of 0.995, sensitivity of 96%, and specificity of 97%. IP-10 was the most effective TB biomarker among 19 plasma miRNAs and proteins. Results reveal that a specific mix of miRNA and protein biomarkers may enhance protein or miRNA biomarker candidates to achieve WHO’s criteria for elevated sensitivity and specificity need several biomarkers. In serum exosomes of TB patients, miRNA-146 expression was significantly elevated compared to the healthy cohort. Additionally, significant increases were noted in the expression levels of miR-425, miR-484, and miR-96. Subsequently, exosomes are crucial for cellular communication, and diseases cause abnormalities in the exosomal process. Exosomal miRNAs have also been suggested by numerous researchers to be a marker of the illness progression, exosomes show promise as therapeutic agents and biomarkers for diagnosis [55–59]. Downregulation of miR-146a-5p following MTB infection enhanced the expression of UBE2L6, which functions as a ubiquitin and ISG15 E2 enzyme. UBE2L6 and miR-146a-5p appear to serve as biomarkers for tuberculosis. The role of UBE2L6 in tuberculosis is significant [60–62].
Additionally, the network’s miRNA hsa-miR-185-5p was 3.65 times more abundant in the cured TB group than in the untreated TB group. In addition, in a case-control study of HIV-negative TB patients, miR-215 was significantly higher after 2 months of medication compared to untreated patients. This study confirmed that the downregulated hsa-miR-215-5p likely affects the TGF-α signalling pathway, as previously reported [29,63,64]. Interestingly, nicotine decreases macrophage phagocytosis. However, miR-296-3p targets SIRPα and reduces its mRNA and protein levels in macrophages by binding to two locations in the 3′-UTR. It is interesting that miR-296-3p inhibits nicotine from boosting SIRPα mRNA and protein levels, indicating its importance in managing the interaction between SIRPα and nicotine in macrophages [65,66].
Various differentially expressed genes association with prognosis of TB
2.6.
The regulation of gene expression is additionally affected by other principal categories of long noncoding RNAs (lncRNAs), small noncoding RNAs (sncRNA), such as small nucleolar RNAs (snoRNA), small nuclear RNAs (snRNA), and PIWI-interacting RNAs (piRNA) [67]. In this context, recent study used CNC to analyse mRNAs and lncRNAs. In TB, differentially expressed lncRNAs target IL6ST, IL18, BCL2, IL5, TLR6, and NOD2. Toll-like receptors (TLRs) identify MTB and its cells, activating the macrophage system and triggering acquired and innate immunity. Aberrant lncRNA expression affects both the pathogenic process of TB and lncRNA polymorphisms linked to susceptibility, demonstrating that lncRNAs play a major role in its pathogenesis. Besides that, in the therapy of TB, lncRNAs are essential. Because the gut microbiota controls lncRNAs to have immune-protective effects against TB, research suggests that the gut-lung axis may be a novel therapeutic target for pulmonary disorders [68–71]. In addition, circRNA functions are still mostly unknown. CircRNAs modify the expression of parental genes and can be produced from exonic or intronic sequences. Four circRNAs of interest were sequenced in the current investigation after microarray screening. Among these, active TB plasma showed up-regulation of hsa_circ_091692, hsa_circ_102295, and hsa_circ_102295 [72,73]. Notably, a preliminary investigation of c-miRNAs in active PTB patients showed 33 downregulated and 59 elevated signatures compared to non-TB controls. This study examined EPTB’s most upregulated and downregulated miRNAs, hsa-miR-425-5p and hsa-miR-4523 [74,75].
Conclusion
This review analysed the diverse functions of miRNAs in MTB and their role in the pathogenesis of the disease. However, miRNAs, also known as epigenetic modulators, regulate post-transcriptional gene expression in many physiologic and pathological processes [76]. Also, the relationship between let-7b-5p and miR-30a-3p and IL-27 and CXCL9/10/11, as well as their expression in TB patients’ lesion tissue and peripheral blood [77], provides a foundation and new ideas for further research in DS and DR-TB.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Global tuberculosis report 2024. Geneva: World Health Organization; 2024. Licence: CC BY-NC-SA 3.0 IGO.
- 2Sundaram K, Vajravelu LK. Functional analysis of genes in Mycobacterium tuberculosis action against autophagosome-lysosome fusion. Indian J Microbiol. 2024;64(2):367–375. doi: 10.1007/s 12088-024-01227-439011011 PMC 11246336 · doi ↗ · pubmed ↗
- 3Pu X, Sheng S, Fu Y, et al. Construction of circ RNA-mi RNA-m RNA ce RNA regulatory network and screening of diagnostic targets for tuberculosis. Ann Med. 2024;56(1):2416604. doi: 10.1080/07853890.2024.241660439435612 PMC 11497567 · doi ↗ · pubmed ↗
- 4Chen LL. The expanding regulatory mechanisms and cellular functions of circular RN As. Nat Rev Mol Cell Biol. 2020;21(8):475–490. doi: 10.1038/s 41580-020-0243-y 32366901 · doi ↗ · pubmed ↗
- 5Zheng ML, Zhou NK, Luo CH. Mi RNA-155 and mi RNA-132 as potential diagnostic biomarkers for pulmonary tuberculosis: a preliminary study. Microb Pathog. 2016;100:78–83. doi: 10.1016/j.micpath.2016.09.00527616444 · doi ↗ · pubmed ↗
- 6Daniel EA, Sathiyamani B, Thiruvengadam K, et al. Micro RN As as diagnostic biomarkers for tuberculosis: a systematic review and meta-analysis. Front Immunol. 2022;13:954396. doi: 10.3389/fimmu.2022.95439636238288 PMC 9551313 · doi ↗ · pubmed ↗
- 7Ying H, Feng Ying S, Yan Hong W, et al. Micro RNA-155 from sputum as noninvasive biomarker for diagnosis of active pulmonary tuberculosis. Iran J Basic Med Sci. 2020;23(11):1419–1425. doi: 10.22038/ijbms.2020.44029.1032433235699 PMC 7671418 · doi ↗ · pubmed ↗
- 8Li J, Zou X. Mi R-652 serves as a prognostic biomarker in gastric cancer and promotes tumor proliferation, migration, and invasion via targeting RORA. Cancer Biomark. 2019;26(3):323–331. doi: 10.3233/CBM-19036131524147 PMC 12826413 · doi ↗ · pubmed ↗