Membrane Proteins ClcB, PtsI, and YcaM Mediate the Bactericidal Effects of Colistin in Escherichia coli
Rhys Donafee, Mohammad Radi, Douglas Bruce Kell, Jesus Enrique Salcedo-Sora

TL;DR
This study identifies three membrane proteins in E. coli that play a role in the bactericidal effects of the antibiotic colistin.
Contribution
The first evidence that ClcB, PtsI, and YcaM are involved in the cytotoxic pathway of polymyxin action in Gram-negative bacteria.
Findings
Knockout of ClcB, PtsI, and YcaM affects E. coli susceptibility to colistin.
The proteins are part of the cytotoxic pathway following polymyxin membrane intercalation.
Findings were validated using growth assays, cytotoxic tests, and in vivo models.
Abstract
Bacteria can be killed very effectively by targeting their first line of protection. The cell membranes (outer and inner membranes) of Gram-negative bacteria are directly targeted by antibiotics, such as polymyxins, via electrostatic interactions with their lipopolysaccharide (LPS) fraction. The downstream effectspreceding the cell deathof the disruptions of the bacterial membranes upon the intercalation of these antibiotics are unknown. By screening a set of E. coli membrane protein knockouts, the absence of three membrane transporters was shown to influence the susceptibility to colistin. Their involvement was corroborated with growth assays on gain-of-function strains, cytotoxic assays, and in vivo infections in an invertebrate animal model. This is the first-time evidence that the disruption caused to membrane proteins, such as the chloride channel ClcB, the sugar transport system…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 1
1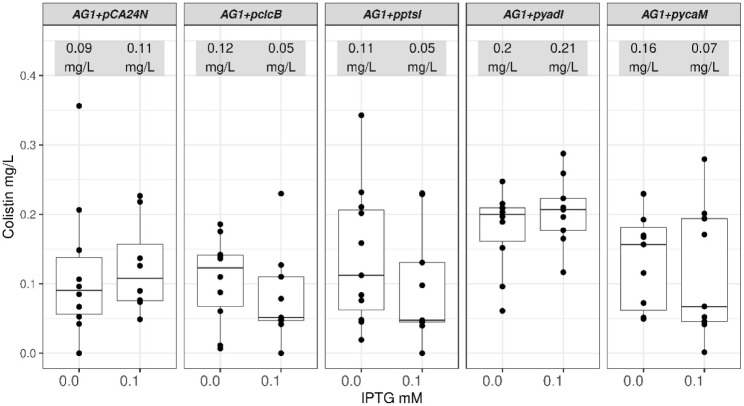 2
2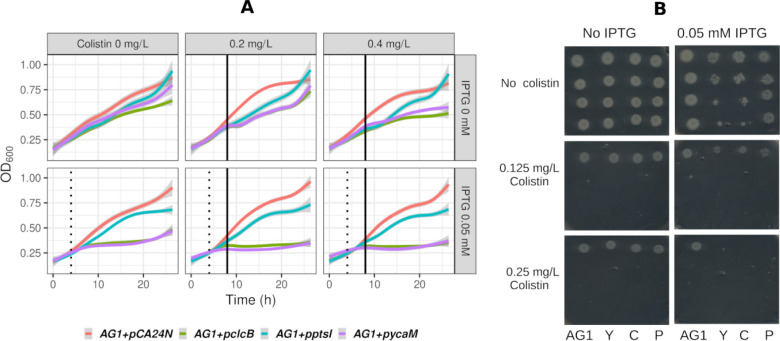 3
3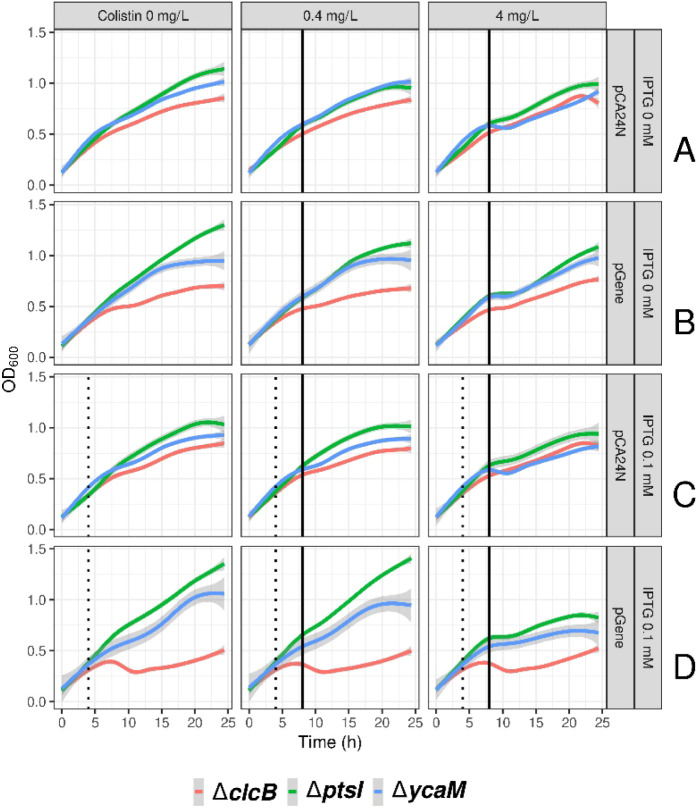 4
4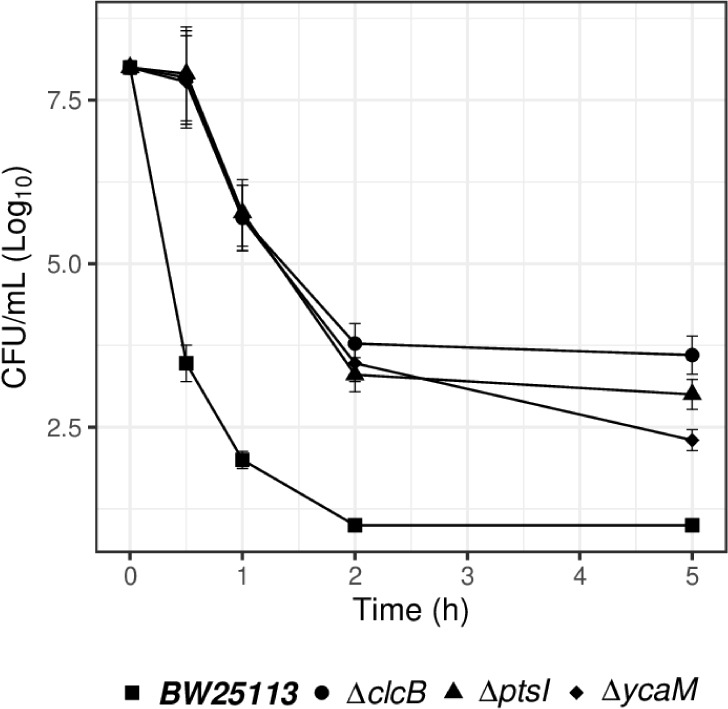 5
5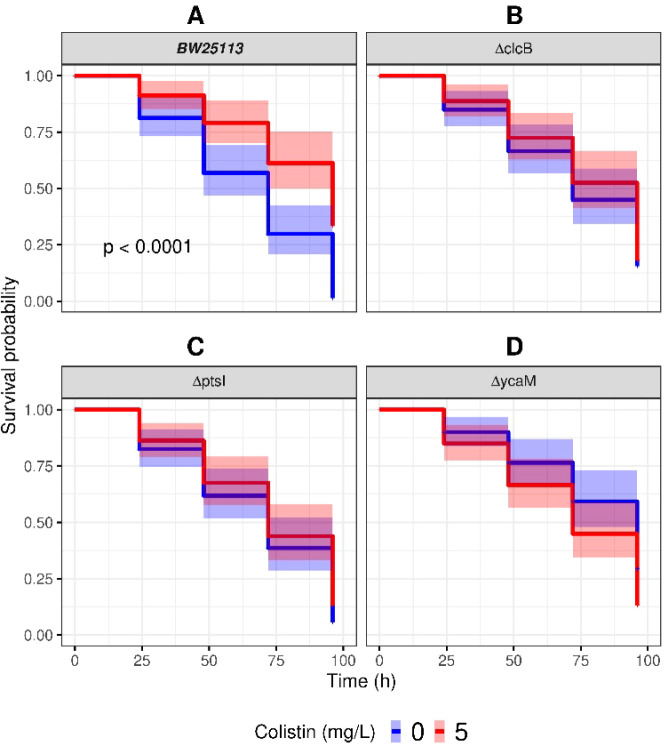 6
6- —Novo Nordisk Fonden10.13039/501100009708
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBacterial Genetics and Biotechnology · Antibiotic Resistance in Bacteria · Vibrio bacteria research studies
Colistin is currently used as a last resort antibiotic against infections by multidrug-resistant Gram-negative bacilli, such as carbapenemase-producing Enterobacteriales, Pseudomonas aeruginosa, and Acinetobacter baumanni.? The World Health Organization (WHO) has listed colistin as critically important because of the increasing use of colistin to treat serious infections in humans in many parts of the world, the prevalence of mcr (mobile colistin resistance) genes, and the spread of colistin-resistant bacteria via the food chain.? Polymyxins such as colistin, and other cell membrane-binding antibiotics, are also effective at targeting antibiotic persister bacteria (stochastic phenotypes that survive high concentrations of antibiotics). ?,?
Colistin (polymyxin E) is a cyclic lipopolypeptide produced using a nonribosomal synthase by Bacillus polymyxa var. colistinus (previously B. colistinus). ?,? Colistin is a polycation that binds lipid A in the lipopolysaccharide (LPS) fraction of the cell outer membrane (OM) of Gram-negative bacteria, displaces the divalent cations calcium (Ca^2+^) and magnesium (Mg^2+^), and impairs the structure of LPS while inserting its hydrophobic terminal acyl chain. This causes an expansion of the OM, permeabilization of the OM and a further movement of colistin beyond the OM. ?−? ? ? This seems to lead to an increase in cell permeability and cell barrier disruptions, including destabilization of membranes and loss of cytoplasmic content.?
Upon exposure to colistin bacteria show some visible cell membrane disruptions within one? and 2 h.? Other morphological postexposure observations include the formation of bleb-like projections from the cell wall of E. coli,? with cytoplasmic material detectable in the extracellular milieu after half an hour of exposure to polymyxins.? However, other reports show cell aggregation rather than cell membrane disruption.?
Modes of resistance to colistin have substantiated the cell membrane-binding properties of polymyxins. A reduction in the net negative charge of lipid A affects polymyxin’s affinity for the OM. Enzymes that mediate chemical modifications of lipid A and cause a loss of sensitivity to polymyxins include plasmid-encoded mcr gene products,? and two-component regulatory systems and sensor kinase systems such as pmrA/pmrB? and phoP/phoQ. ?,? Inhibiting the production of fatty acids destined for lipid A biosynthesis overcomes resistance to colistin altogether.?
This said, the mechanisms underlying the bactericidal effects of polymyxins upon their interactions with the outer, and the inner membranes? are not known. Furthermore, the cell permeabilisation and killing properties of polymyxins might not have a causal relationship. Modified versions of polymyxin B (polymyxin B nonapeptide) that still have cell membrane-binding properties and cause leakage of cytoplasmic contents, do not have antibacterial activity.?
Part of the answer could be in the role played by the proteins within the inner membrane (cell membrane) of Gram-negative bacteria.? The often generally overlooked integral membrane proteins and membrane-associated proteins are the most abundant component by mass of the cell membranes,? and responsible for the transport of any number of small molecules. ?−? ? It thus seems logical to investigate the involvement of membrane proteins in the bactericidal effects of a membrane-binding antibiotic such as colistin. We describe here that at least three membrane-associated proteins seem to mediate cell death caused by colistin. Specifically, the absence of these proteins caused tolerance while their presence sensitized E. coli to this antibiotic. The relevance of these findings to in vivo infections is illustrated in an invertebrate animal model.
Results
The Absence of Certain
Membrane Transporters in E. coli Gene Knockouts Reduced Their Sensitivity to Colistin
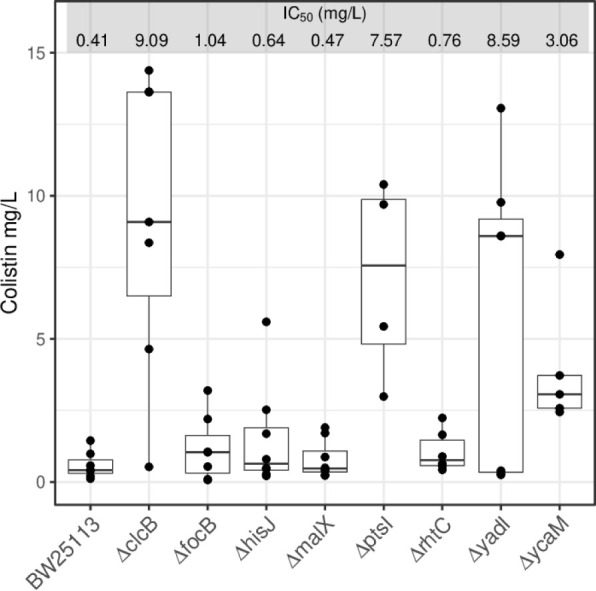
A set of 534 strains lacking membrane transporters and other membrane associated proteins from the E. coli Keio collection of gene knock outs (KOs)? was used to screen for tolerance to colistin. They were exposed to incremental concentrations of colistin up to 5 mg/L in complex media and ranked according to their growth in comparison to that of the reference strain for that collection, the E. coli BW25113 strain (Supporting Table S1). From that initial list, eight KO strains were selected for further growth inhibition assays (IC_50_): ΔclcB, ΔfocB, ΔhisJ, ΔmalX, ΔptsI, ΔrhtC, ΔyadI and ΔycaM. The IC_50_ data from four or more replicates for each of these strains showed that the KO strains ΔclcB, ΔptsI, ΔyadI and ΔycaM had IC_50_ median values six to 20-fold that of the reference strain BW25113 (Figure). A Welch-Anova analysis showed that there were significant differences in the IC_50_ values for colistin between all different strains (p = 2.2e-3). From the pairwise comparisons against the reference strain, the KOs strains ΔclcB, ΔptsI, ΔyadI and ΔycaM showed to have IC_50_ values significantly different (Figure).
E. coli KOs with increased tolerance to colistin. The median for the IC50 values of the E. coli reference strain BW25113 is presented against those of the eight KO strains initially observed to have different levels of higher tolerance to colistin: ΔclcB, ΔfocB, ΔhisJ, ΔmalX, ΔptsI, ΔrhtC, ΔyadI and ΔycaM. Data represent at least four biological replicates (black dots). The median values for each IC50 range are listed as mg/L. The data had a normal distribution (Shapiro-Wilk’s test p = 9.3e-4) with non-homogenous variances between strains (Levene’s test p = 4.2e-4). Thus, the pairwise comparisons between BW25113 and ΔclcB, ΔptsI, ΔyadI and ΔycaM were carried out with the Game-Howell test showing respectively p values of 5.5e-3, 2e-2, 3e-2 and 2e-2.
Expression of the Membrane Transporters clcB, ptsI and ycaM in the ASKA AG1 Strain Increased Their
Sensitivity to Colistin
The availability of the ASKA collection of E. coli transformed with plasmids expressing most of the protein-encoding genes for these bacteria allowed us to assess if the opposite (increased sensitivity to colistin) was observed in strains expressing the genes clcB, ptsI, yadI and ycaM. The ASKA collection uses the E. coli K-12 derivative strain AG1 carrying recombinant constructs in an IPTG-inducible lacO-T5 hybrid promoter expression system in the multicopy plasmid pCA24N (CmR, lacIq).? The E. coli reference strain AG1 as used here was a true control transformed with an empty version of the pCA24N plasmid.
Strains with the recombinant plasmids carrying clcB, ptsI, yadI or ycaM (denoted as AG1+pclcB, AG1+pptsI, AG1+ pyadI and AG1+pycaM, respectively) showed IC_50_ median values similar to that of the reference AG1 strain (AG1+pCA24N) (Figure): 0.09 – 0.2 mg/L. However, when 0.1 mM IPTG was added, IC_50_ values decreased to half or less for those strains carrying recombinant plasmids: 0.05 – 0.07 mg/L; except for the strains carrying the empty vector (AG1+pCA24N), and the strain expressing pyadI (AG1+pyadI) (Figure). The latter construct was not included in the next following parts of this work due to the lack of consistent with the expected trends of sensitivity to colistin upon addition of IPTG (Figures and ?).
E. coli strains expressing clcB, ptsI and ycaM are more sensitive to colistin. The median values for the IC50 of the E. coli reference strain AG1 carrying the empty pCA24N (AG1+pCA24N) plasmid and those strains containing this plasmid with either of four genes were calculated from samples incubated in the absence or the presence of 0.1 mM IPTG. Three of those strains (AG1+pclcB, AG1+pptsI and AG1+pycaM) showed a trend of increased sensitivity to colistin when the cognate membrane transporter was induced. Data illustrated originated from at least eight biological repeats (black dots). The median values for each IC50 range are listed as mg/L. The pairwise comparisons within each strain in the absence versus the presence of IPTG had the following p values: p > 5e-2 for AG1+pCA24N, 3.9e-2 for AG1+pclcB, 2.8e-2 for AG1+pptsI, p > 5e-2 for AG1+pyadI and 3.9e-2 for AG1+pycaM.
Given the differences noted above, the AG1+pclcB, AG1+pptsI and AG1+pycaM strains were selected for kinetic growth assays (Methods). In the kinetic assays (FiguresA and ?) cultures were allowed to grow before adding IPTG and colistin. A greater inoculum size allowed to have more cells expressing these membrane transporters upon addition of IPTG (0.05 mM) after 4 h of growth. After a window of IPTG-driven protein expression colistin was then added. At that time the membrane transporters were expected to have reached detectable protein levels induced by IPTG.? When colistin was added the cell mass of most cultures had reached OD_600_ values up to 0.5, which explains the subsequent growth observed in some of the strains, even in the presence of high colistin concentrations (FiguresA and ?). The attenuation of the bactericidal activity of colistin by inoculum size has been characterized.?
Growth inhibitory effects of colistin in E. coli expressing the membrane transporters clcB, ptsI and ycaM. (A) The abscissa represents time. The ordinate represents the culture media optical densities at 600 nm. The AG1 control and the recombinant samples are color-coded: AG1+pCA24N is red, AG1+pclcB is green, AG1+pptsI is blue and AG1+pycaM is magenta. Colistin is represented here in the 0.2 and 0.4 mg/L samples. The full range of colistin concentrations analyzed is presented in Table S2. The dotted vertical black line represents the addition of 0.05 mM IPTG at 4 h. The solid vertical black line represents the addition of colistin at 8 h. The data represented in this figure are tabulated in Supporting Table S2. (B).
Expression of ptsI and ycaM resensitizes E. coli KO strains to colistin. No-colistin or colistin samples at 0.4 and 4 mg/L are presented column-wise. Rows A and B represent growth (absorbance at OD600) in the absence of IPTG. Rows C and D represent growth in the presence of 0.1 mM IPTG. Strains transformed with the empty pCA24N plasmid are in rows A and C. Strains transformed with the recombinant plasmids expressing their cognate genes (denoted as pGene) are in rows B and D. Each KO strain carrying either the empty plasmid or the recombinant plasmid is color coded: ΔclcB is red, ΔptsI is green and ΔycaM is blue. Dashed lines represent the time point for the addition of IPTG while the solid line represents the addition of colistin.
In the kinetic assays all three strains had visible lower culture densities (absorbance at OD_600_) than the AG1+pCA24N control in the presence of colistin. Those differences widened in the samples upon induction with 0.05 mM IPTG (FigureA and Supporting Table S2), particularly for the AG1+pclcB and AG1+pycaM samples. The growth of the AG1+pptsI samples seemed to have had an intermediate sensitivity to the inhibitory effects of colistin (FigureA).
The Presence of clcB, ptsI, and ycaM Mediated the Growth Inhibition
Caused by Colistin in E. coli
AG1 is the control strain carrying the empty plasmid pCA24N. Y, C, and P represent the AG1 strains carrying the plasmids pycaM. pclcB or pptsI, respectively. Each strain is represented by four row-wise 1:10 dilutions of culture broth used for spotting. Two different colistin concentrations are shown, in the absence or the presence of IPTG (0.05 mM).
The overexpression of membrane transporters is often toxic to the bacterial host.? This was evident in the growth assays where AG1+pclcB and AG1+pycaM were visibly inhibited in the IPTG-induced samples in the absence of colistin (Figure). However, further assays based on growth in solid media substantiated, at two incremental doses, the observed sensitization to colistin caused by the expression of clcB, ptsI and ycaM additional to the baseline toxicity due to the expression of these proteins upon their induction with IPTG (Figure). Here, both colistin and IPTG were present in the solid media and therefore the inoculum for strain was exposed to colistin and IPTG from the beginning as opposed to the planktonic cultures where the initial cultures were allowed to grow beforehand (FiguresA and ?).
Gene Complementation Induced
Sensitivity to Colistin in Otherwise Tolerant E. coli BW25113 KO Strain
Each of the E. coli KOs ΔclcB, ΔptsI and ΔycaM from the Keio collection were transformed with either an empty pCA24N plasmid or with a recombinant plasmid carrying their cognate gene pclcB, pptsI or pycaM. The identity of the recombinant constructs in pCA24N plasmids as well as the empty plasmid were all confirmed by DNA sequencing (sequence data available upon request). Equally, whole genome sequencing for the ΔclcB, ΔptsI and ΔycaM strains confirmed their loci to have been replaced by the S. aureus aadD1 gene encoding for an aminoglycoside O-nucleotidyltransferase that confers the kanamycin resistance marker for the E. coli Keio KO collection.
The strains ΔclcB, ΔptsI and ΔycaM carrying the recombinant plasmids pclcB, pptsI and pycaM, respectively, had their growth affected in the absence of IPTG (FigureB) when compared against the empty plasmid controls (pCA24N, FigureA). We interpreted this to be the consequence of a leakier gene expression system in the E. coli BW25113 background strain in comparison to the E. coli AG1 strains (Figure). The BW25113-based strains seemed to have a higher basal lac operon activity, as is sometimes observed when it is assembled with the endogenous bacterial promoter T5,? as is the case of the pCA24N plasmid. Growth was further affected by the presence of 0.1 mM IPTG, particularly for the strain carrying pclcB which impeded to observe any further growth inhibition upon addition of colistin in this case. Nevertheless, it was still possible to observe the growth inhibitory effect of colistin in relation to the heterologous expression of ptsI and ycaM (FigureD).
E. coli KOs increased tolerance to high concentrations of colistin. Viable cell counts as Log10 of CFU/mL are presented for bacteria exposed up to 5 h to 5 mg/L of colistin. The inoculum size is 108 cfu/mL for all strains. The E. coli reference strain is BW25113. The three KO strains are ΔclcB, ΔptsI and ΔycaM. Data represent three biological replicates.
Time-Kill Assays Support the Mediation of clcB, ptsI, and ycaM in the Action
of Colistin
We measured the survival of the reference strain in comparison to that of ΔclcB, ΔptsI and ΔycaM upon exposure to 5 mg/L of colistin. A steep decline in viable cell counts (cfu/mL) indicated a bactericidal effect on BW25113 with a drop of 6 orders of magnitude in the first hour Figure). At the same time point the KO strains reduction in viable cells was approximately 3 3 orders of magnitude. By the second and fifth hour the cell viability was still 2 to 3 Log_10_ values higher in the KOs than in BW25113.
The Absence of clcB, ptsI, and ycaM Reduces the Efficacy of Colistin In Vivo
The larvae of the Greater wax moth (Galleria mellonella) provide an invertebrate animal model that is often used to study bacterial infections. ?−? ? ? A positive correlation between the virulence and immune responses between mammalian models and the innate immune response of G. mellonella has been established for several infections. ?,? We used larvae of G. mellonella to determine potential differences, in the absence as well as the presence of colistin, in the survival of the larvae infected with the E. coli reference BW25113 strain or the KOs ΔclcB, ΔptsI and ΔycaM. The data were represented as time-to-event analysis using Kaplan–Meier curves and using log rank tests to test for significance between strains. Each larva was injected with 50 ng of colistin. At an average weight of 250 mg, each larva received approximately 0.2 mg/kg of colistin.
In the absence of colistin, at day four the survival probability of larvae infected with E. coli BW25113 was 8.2% (Figure, Supporting Table S3). When the infected larvae were also injected with colistin, the survival trend increased to 47% for BW25113 at day four (Figure). Thus, the presence of colistin increased the survival of larvae infected with the reference strain by over 6-fold.
Galleria mellonella larvae infected E. coli KOs have reduced survival rates. Kaplan–Meier survival curves are shown for larvae first infected with the reference strain BW25113 (A) or either of the KOs ΔclcB (B), ΔptsI (C) and ΔycaM (D). Colistin control larvae (blue curves) were subsequently injected with PBS. Data of larvae subsequently injected with 5 mg/L of colistin are represented by the red curve. Data represents up to 80 larvae per group, from at least four different biological replicates. Shaded areas cover the 95% confidence intervals (Supporting Table S3).
The probabilities of survival for the larvae injected with the KOs were 14%, 10%, and 18% for ΔclcB, ΔptsI and ΔycaM, respectively, in the absence of colistin (FigureB,D, Supporting Table S3). In the presence of colistin, survival rates were 13%, 12%, and 8% for ΔclcB, ΔptsI and ΔycaM, respectively (FigureB,D, Supporting Table S3). Therefore, the presence of the antibiotic made no significant difference to those larvae infected with the KOs.
Discussion
Membrane lipid composition and integrity can modulate the structure and activity of transport proteins. ?,? The derangement of the bacterial cell membranes after its interactions with polymyxins such as colistin ?−? ? ?,?,? are thus expected to alter the integrity and activity of at least some membrane proteins. Colistin also reaches the cytoplasmic side of the inner membrane whereby it can disrupt the initial movement of LPS into the cell membrane. This would be expected to add to the irreversible changes in the integrity of the bacterial membrane.? This is corroborated by the reports of efflux inhibitors increasing the susceptibility of bacteria to colistin. ?,? Altogether, disruptions of cellular homeostasis are expected to be a primary cytotoxic effect caused by permeability disruption.? Our findings here point to specific membrane proteins whose malfunction can affect this disruption of cell homeostasis. Given their location and roles, the dysregulation of PtsI, ClcB and YcaM provides evidence for specific membrane protein dysgenesis as a mediator of the colistin’s lethality.
ClcB is one of the two CLC-type voltage-gated chloride channels found in bacteria, although the function of ClcB is largely unknown.? It is thought to act as an electrical shunt for an outwardly directed proton pump that is linked to amino acid decarboxylation, and is of importance in extreme acid resistance responses.? PtsI is enzyme I of the phosphoenolpyruvate:carbohydrate phosphotransferase system (PTS).? This system mediates the transport of sugars and their intracellular phosphorylation in bacteria. PtsI localizes to the inner surface of the cytoplasmic membrane where it interacts with sugar-specific inner-membrane permeases. PTS systems regulate carbon utilization in switching between the hierarchical (diauxic) and coutilization (nonspecific) use of sugars. YcaM has been classified by sequence similarity as a Glutamate:GABA antiporter, part of the amino acid-polyamine organocation superfamily.? YcaM function has not been characterized. It seems to be expressed when bacteria are in the presence of abundant nutrients in vitro.?
The Keio collection of E. coli gene KO used here has been previously profiled against a set of antibiotics that included colistin.? However, opposite to our approach, these authors assessed only the sensitivity of KOs to antibiotics (we screened for tolerance to colistin). This could explain why the membrane protein-encoding genes reported here do not appear in the list of genes reported in that work as conferring different levels of sensitivity to colistin.? A subsequent study, although polymyxins were not included, showed other membrane transporters mediating the growth inhibitory effects of a number of antibiotics in a subset of the Keio collection.?
The dysregulation of ClcB can affect the pH and ionic homeostasis of bacteria. Alkaline pH has a synergistic effect with colistin across several species of Gram-negative.? This observation was extended to intrinsically polymyxin-tolerant Gram-negative bacteria Burkholderia spp..? PtsI has been shown to be involved in ROS-mediated cellular responses to antimicrobials and other environmental stressors, where a lack of PtsI function allowed tolerance to different types of stressors. Such pan-tolerance in the absence of PtsI seems to be based on a reduced capacity for metabolic shifts between different central carbon metabolism pathways. This offers protection from stress-mediated ATP surges, with a concomitant reduced accumulation of ROS.? The potential mechanisms by which YcaM would mediate the toxicity of a membrane-acting antibiotic such as colistin will require further investigation, particularly since the function of this membrane protein is itself still to be characterized. Interestingly, while ClcB and PtsI have homologues in other Gram-negative bacteria, YcaM seems to be conserved only in E. coli and missing even in closely related species such as Salmonella sp.?
The evidence presented here opens new views regarding the mechanism of action of polymyxins. These antibiotics kill the bacterial cell via a series of discernible steps as opposed to a single event of detergent-like cell lysis. In contrast to detergents, the interactions of polymyxins with the cell membranes, mainly with the LPS fraction, are driven by electrostatic interactions (i.e., not hydrophobic), which cause membrane disorganization by antibiotic intercalation and self-promoted permeation.? It is evident that the ensuing cytotoxicity follows then a pathway that includes the disfunction of critical membrane transporters, represented here for E. coli by the chloride channel ClcB, the sugar transport system (at least PtsI) and the Glutamate:GABA antiporter YcaM. It is possible that these transporters are directly targeted by colistin. However, the genes encoding them do not seem to have been reported in gene mutation studies of colistin resistance.? We would therefore argue that their disruption is more likely to be secondary to the membrane damage caused by polymyxins.
Methods
Strains
The E. coli Keio collection of gene KO strains was provided by the National Institute of Genetics, Mishima, Shizuoka, Japan. ?,? A subset of 534 strains from the Keio Collection whose cognate genes are annotated as encoding for membrane associated proteins were selected for this study (Supporting Table S1). Individual knockout strains highlighted in this work are as follows: clcB (F-, Δ(araD-araB)567, ΔlacZ4787(::rrnB-3), λ-, ΔclcB740:kan, rph-1, Δ(rhaD-rhaB)568, hsdR514); ptsI (F-, Δ(araD-araB)567, ΔlacZ4787(::rrnB-3), λ-, ΔptsI745:kan, rph-1, Δ(rhaD-rhaB)568, hsdR514); ycaM (F-, Δ(araD-araB)567, ΔlacZ4787(::rrnB-3), λ-, rph-1, ΔycaM723:kan, Δ(rhaD-rhaB)568, hsdR514) ; ydaI (F-, Δ(araD-araB)567, ΔlacZ4787(::rrnB-3), λ-, ΔyadI746:kan, rph-1, Δ(rhaD-rhaB)568, hsdR514). Whole genome sequencing was used to verify the absence of the cognate genes in the clcB, ptsI and ycaM KO strains (Plasmidsaurus Inc.). We also used strain overexpressing clcB, ptsI and ycaM from the ASKA collection: E. coli K-12 derivative strain AG1 [recA1 endA1 gyrA96 thi-1 hsdR17 (r K– m K+) supE44 relA1] carrying recombinant constructs for those genes in the IPTG-inducible and multicopy plasmid pCA24N (CmR, lacIq).? The composition of those recombinants in pCA24N plasmids was also confirmed by whole plasmid DNA sequencing (Plasmidsaurus Inc.; sequencing data available upon request). The genes pCA24N vectors carry in the ASKA strains have an amino-terminal His-tag. No other further tags (e.g., GFP) are present in these constructs.
Inhibitory Concentration
Assays
Bacterial cultures were carried out routinely in complex media (lysogeny broth, Merck LB 110285).? Colistin was purchased from MP Biomedicals, cat number 194157, activity ≥ 15,000 U/mg. The screening of the Keio collection subset of 534 strains (Supporting Table S1) to incremental concentrations of colistin was carried out by replicating their glycerol stocks into 384-well plates with 50 μL of LB containing 30 μg/mL of kanamycin. These sealed plates were incubated overnight at 37 °C. The overnight cultures were diluted 1 in 100 in 50 μL of fresh LB without kanamycin in polystyrene 384-well plates with a transparent bottom (Sigma M6936–40EA). End-point growth inhibitory concentrations (IC_50_) were calculated from microtitration assays. Overnight cultures in LB were diluted 1 in 100 in fresh LB. Fifty μL of 2-fold dilutions of colistin were prepared in 96-well plates to which 50 μL of fresh bacterial culture were added. End point read outs of the media turbidity at 600 nm (OD_600_) were taken after 24 h at 37 °C.
Kinetic Assays
Continuous growth assays to follow the effect of protein expression and subsequent exposure to colistin were carried out with freshly 1 in 100 diluted cultures in 180 μL at 30 °C in 96 microwell plates and shaking at 200 rpm. After 4 h protein expression was induced with 0.05 mM IPTG. At 8 h colistin was added to final concentrations 0.05, 0.1, 0.2, 0.4, 0.8, 1.6, 3.2 mg/L. OD_600_ readouts were collected every 15 min in a BMG LabTech CLARIOstar Plus plate reader for 27 h.
Time-Kill Assays
The inoculum originated from overnight cultures diluted 1:500 in 5 mL of complex media and allowed to grow for 2h, 37 °C and shaking at 250 rpm. An inoculum of 10^8^ cfu/mL showed at least one hundred viable cells in the reference strain BW25113 throughout the 5 h of exposure to 5 mg/L of colistin. Freshly inoculated 5 mL cultures for BW25113 and KO strains were exposed to colistin for up to 5 h. Samples of 0.02 mL were taken from each culture at 0.5, 1, 2, and 5 h. Those volumes were diluted 1:10 in 0.180 mL of complex media, prepared for each strain row-wise in a 96-well plate. Diluted aliquots were plated out in solid complex media in standard Petri dishes and incubated overnight at 37 °C. Viable cells were counted manually from plates with colonies clearly visible and separated from each other
Gene Complementation
E. coli KO strains ΔclcB, ΔptsI and ΔycaM were transformed with the recombinant plasmids pCA24N carrying either their cognates genes, or empty plasmids as controls. The presence of those plasmids in their respective strains was verified by enzyme restriction analysis before assessing their antibiotic sensitivity. The IC_50_ assays for colistin were carried out as described above.
Larval Survival Assay
Larvae of the Greater wax moth Galleria mellonella were purchased from Livefoods UK Ltd. (Somerset, UK). Larvae were injected the same day upon delivery. Healthy large larvae (approximately 250 – 350 mg body weight) were selected. They had a uniform cream color without spots or marks indicating melanisation. Ten larvae per condition were injected with 10^4^ cells of E. coli (for both reference strain and KO strains) in10 μL in either of the penultimate prolegs, using a 10 μL Hamilton 750 syringe (Hamilton Company, UK). After injections, the larvae were placed into Petri dishes and incubated at 37 °C. Syringes were cleaned with 70% ethanol between groups of injection. Larvae mortality was followed for 96 h by counting death individuals (black or dark brown in color) every 24 h. Colistin was injected in the opposite penultimate proleg, 30 min after they had been injected with bacteria. The control group was injected with phosphate buffer saline (PBS).
Data Analysis
The inhibitory concentrations of colistin that kills half of the bacteria population (IC_50_) were calculated with the four-parameter logistic model as implemented in the R package drc
?,? and python package py50,? following the guidelines for relative calculations according to the spread of the data.? The Shapiro-Wilk test, Levene’s test, Welch-Anova and Games-Howell’s posthoc tests (pairwise comparisons) were carried out using the required packages in R.? Visualization and annotation of bacterial genomes and plasmid sequences used Proksee.? Survival analyses of larvae were carried out with the time to event model Kaplan–Meier which estimates survival differences in survival times between compared groups as implemented in the R packages survival and survminer.
Supplementary Material
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Andrade F. F.Silva D.Rodrigues A.Pina-Vaz C.Colistin Update on Its Mechanism of Action and Resistance, Present and Future Challenges Microorganisms 2020811171610.3390/microorganisms 811171633147701 PMC 7692639 · doi ↗ · pubmed ↗
- 2World Health Organization (WHO) WHO List of Medically Important Antimicrobia: antimicrobial Resistance Division (AMR) Department of Global Coordination and Partnership of Antimicrobial Resistance; WHO: Geneva, Switzerland, 2024.
- 3Salcedo-Sora J. E.Kell D. B.A Quantitative Survey of Bacterial Persistence in the Presence of Antibiotics: Towards Antipersister Antimicrobial Discovery Antibiotics 20209850810.3390/antibiotics 908050832823501 PMC 7460088 · doi ↗ · pubmed ↗
- 4Cui P.Niu H.Shi W.Zhang S.Zhang H.Margolick J.Zhang W.Zhang Y.Disruption of Membrane by Colistin Kills Uropathogenic Escherichia coli Persisters and Enhances Killing of Other Antibiotics Antimicrob. Agents Chemother.201660116867687110.1128/AAC.01481-1627600051 PMC 5075097 · doi ↗ · pubmed ↗
- 5Ito M.Aida K.Uemura T.Biosynthesis of colistin by Bacillus colistinus Koyama Biochim. Biophys. Acta, Protein Struct. Mol. Enzymol.1970213124424710.1016/0005-2787(70)90030-44321273 · doi ↗ · pubmed ↗
- 6Tambadou F.Caradec T.Gagez A. L.Bonnet A.Sopena V.Bridiau N.Thiery V.Didelot S.Barthelemy C.Chevrot R.Characterization of the colistin (polymyxin E 1 and E 2) biosynthetic gene cluster Arch. Microbiol.2015197452153210.1007/s 00203-015-1084-525609230 · doi ↗ · pubmed ↗
- 7Davis S. D.Iannetta A.Wedgwood R. J.Activity of colistin against Pseudomonas aeruginosa: inhibition by calcium J. Infect. Dis.1971124661061210.1093/infdis/124.6.6104331350 · doi ↗ · pubmed ↗
- 8Koike M.Iida K.Matsuo T.Electron microscopic studies on mode of action of polymyxin J. Bacteriol.196997144845210.1128/jb.97.1.448-452.19694303542 PMC 249634 · doi ↗ · pubmed ↗