Assessment of bacterial proteome remodeling under virulence-inducing conditions for molecular pharming applications
Kiyan Kheradvar, Nicholas Prudhomme, Michael D. McLean, Doug Cossar, Jennifer Geddes-McAlister

TL;DR
The study explores how bacteria change their proteins when triggered by virulence-inducing conditions, which could help in making plant-based pharmaceuticals.
Contribution
Leaf extracts are proposed as a cost-effective alternative to acetosyringone for inducing virulence in bacteria for molecular pharming.
Findings
Proteomics revealed increased abundance of virulence proteins in Agrobacterium tumefaciens after exposure to inducers.
Leaf extract from Nicotiana benthamiana is a potential low-cost alternative to acetosyringone for virulence induction.
Abstract
Using mass spectrometry-based proteomics, we identified virulence proteins from Agrobacterium tumefaciens with significantly increased abundance following exposure to chemical (acetosyringone) and biological (leaf extract from Nicotiana benthamiana) inducers. Leaf extracts could be a cost-effective alternative to acetosyringone for inducing pathogen virulence for the molecular pharming industry.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
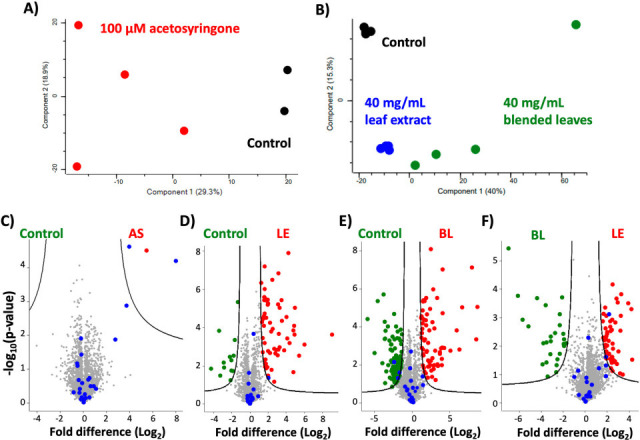 Fig 1
Fig 1- —Natural Sciences and Engineering Research Council of Canadahttp://dx.doi.org/10.13039/501100000038
- —Ontario Graduate Scholarship
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsTransgenic Plants and Applications · Toxin Mechanisms and Immunotoxins · Plant tissue culture and regeneration
ANNOUNCEMENT
Molecular pharming is the process of producing biologics (e.g., antibodies, vaccines) using the natural infection process between the plant bacterial pathogen, Agrobacterium tumefaciens, and the host, Nicotiana benthamiana (1, 2). The technology supports increased scalability, enhanced safety, lower production costs, and proper molecular modifications compared with cell production systems (3). During the infection process, A. tumefaciens delivers transfer (T)-DNA from its tumor-inducing (Ti) plasmid through the type-IV secretion system (T4SS) into the plant cell for production of specific targets (4–6). In this study, we measured changes in the bacterial proteome under chemical and biological conditions known to influence infectivity (7).
Plant leaves were harvested from N. benthamiana (KDFX plant line derived from USDA accession “TW16”), sterilized (i.e., 30 s wash in 70% ethanol, 10 s wash in sterile Milli-Q [MQ] water, 10 min wash in 0.6% active bleach, and two 10 s washes in sterile MQ water), and dried aseptically prior to flash freezing and storage at −80 °C. A. tumefaciens (strain At564A, derived from C58, containing the Trastuzumab vector pFC0058) was grown in quadruplicate in Luria-Bertani broth with kanamycin (50 µg/mL) and rifampin (25 µg/) overnight at 28 °C with shaking (200 rpm) in the dark and sub-cultured to a normalized OD_600nm_ of 0.2, as we previously described (8, 9). Bacterial virulence was induced at room temperature for 24 h in the dark by: 100 µM acetosyringone (TCI Chemicals) in agroinfiltration medium , 40 mg/mL plant leaves blended in agroinfiltration medium, or supernatant of 40 mg/mL segmented plant leaves pre-incubated with agroinfiltration medium for 48 h at room temperature (referred to as leaf extract). An uninduced control contained only agroinfiltration medium. Notably, the acetosyringone and untreated control experiment was performed independently from the biological (leaf extract and blended leaf) experiments.
) overnight at 28°C with shaking (200 rpm) in the dark and sub-cultured to a normalized OD_600nm_ of 0.2, as we previously described (8, 9). Bacterial virulence was induced at room temperature for 24 h in the dark by: 100 µM acetosyringone (TCI Chemicals) in agroinfiltration medium , 40 mg/mL plant leaves blended in agroinfiltration medium, or supernatant of 40 mg/mL segmented plant leaves pre-incubated with agroinfiltration medium for 48 h at RT (referred to as leaf extract). An uninduced control contained only agroinfiltration medium. Notably, the acetosyringone and untreated control experiment was performed independently from the biological (leaf extract and blended leaf) experiments.
Cell pellets were collected at 4,696×g for 10 min, washed twice with phosphate-buffered saline (PBS), and processed as previously described (9, 10). Briefly, pellets were resuspended in 100 mM Tris-HCl (pH 8.5) containing a proteinase inhibitor cocktail tablet, lysed in 2% sodium dodecyl sulfate (SDS) via probe sonication on ice bath (30% amplitude, 3 cycles of 30 s on/30 s off). After the addition of dithiothreitol (DTT; 10 mM final concentration), samples were incubated at 95 °C for 10 min at 800 rpm, cooled, and iodoacetamide (IAA; 55 mM final concentration) was added, followed by incubation in the dark at RT for 20 min. Proteins were precipitated overnight using acetone (80% final volume) at −20 °C, centrifuged at 16,100×g, washed twice with 80% acetone, air-dried, resolubilized in 8 M Urea/40 mM HEPES, and quantified by tryptophan release (11). Samples (25 µg) were digested overnight at 37 °C with trypsin/LysC (protein:enzyme 50:1 w/w) and purified on STAEG-tips (12). Dried peptides were quantified at OD_205nm_ and 3 µg of peptides was resuspended in 0.1% (v/v) trifluoroacetic acid, separated by liquid chromatography (RSLC EasyNano, Thermo Fisher) and electrospray ionization (50 cm PepMap column of 75 µm and C18 resin) coupled to an Orbitrap Exploris 240 (Thermo Fisher) mass spectrometer operated in data dependent-acquisition mode (MS1 scan for top10 peptides, MS2 scan for peptide sequencing, m/z range 400–1,600, resolution 60,000). Peptides were analyzed using a 180 min gradient for chemical conditions (acetosyringone) as the positive control; peptides were analyzed using a 90 min gradient for biological conditions (leaf extract and blended leaves) for instrument time optimization.
Mass spectrometry data were analyzed using MaxQuant (v. 2.1.0.0) and Perseus (v. 2.0.7.0) software (13, 14). The reference sequence of A. tumefaciens (16 December 2019; 5,344 proteins) supplemented with Trastuzumab heavy chain, light chain, two P19 sequences, and neomycin phosphotransferase II (15) was used for searching. Default parameters were set in MaxQuant with label-free quantification (minimum ratio count of 2), minimum peptides of 2, and match between runs enabled. In Perseus, filtering for modified by site (e.g., post-translational modification), potential contaminants, and reverse peptides was performed and proteins present in three of four replicates within one group were retained. NaN values were imputed from a normal distribution (width 0.3, down-shift 1.8), and “Majority protein IDs” were annotated using the UniprotKB database (16). Volcano plots evaluated significant changes in protein abundance by Student’s t-test, P < 0.05, Benjamini-Hochberg (17) false discovery rate = 0.05, S_0_ = 1.
We observed clear separation between the controls and chemical (acetosyringone) (Fig. 1A) or biological (leaf extract and blended leaves) (Fig. 1B) treated samples. Only three significantly different proteins were defined by chemical treatment (Fig. 1C) compared with 76 proteins significantly different between the control and leaf extract (Fig. 1D), and 174 proteins significantly different between control and blended leaves (Fig. 1E). Importantly, virulence-associated bacterial proteins (e.g., VirH1, VirH2, T4SS) (2) were significantly increased under chemical treatment (acetosyringone), biological treatment (leaf extract), and upon comparison between leaf treatments (Fig. 1E). Overall, this study demonstrates proteome remodeling of A. tumefaciens in response to chemical and biological treatments, but limited insight into regulation of virulence factor production is evident.
Proteome profiling of A. tumefaciens under chemical and biological induction of virulence. (A) Principal component analysis of control (agroinfiltration medium) and chemical (acetosyringone) conditions. (B) Principal component analysis of control (agroinfiltration medium) and biological (leaf extract and blended leaves) conditions. (C) Volcano plot for control (agroinfiltration medium) and chemical (AS; acetosyringone) conditions. (D) Volcano plot for control (agroinfiltration medium) and biological (LE; leaf extract) conditions. (E) Volcano plot for control (agroinfiltration medium) and biological (BL; blended leaves) conditions. (F) Volcano plot for leaf extract (LE) and blended leaves (BL). Blue = known virulence-associated proteins. For volcano plots, Student’s t-test P-value < 0.05, FDR = 0.05, S0 = 1. Performed in biological quadruplicate.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Fischer R, Buyel JF. 2020. Molecular farming - the slope of enlightenment. Biotechnol Adv 40:107519. doi:10.1016/j.biotechadv.2020.10751931954848 · doi ↗ · pubmed ↗
- 2Prudhomme N, Pastora R, Thomson S, Zheng E, Sproule A, Krieger JR, Murphy JP, Overy DP, Cossar D, Mc Lean MD, Geddes‐Mc Alister J. 2024. Bacterial growth‐mediated systems remodelling of Nicotiana benthamiana defines unique signatures of target protein production in molecular pharming . Plant Biotechnol J 22:2248–2266. doi:10.1111/pbi.1434238516995 PMC 11258984 · doi ↗ · pubmed ↗
- 3Chen Q, Davis KR. 2016. The potential of plants as a system for the development and production of human biologics. F 1000 Res 5:F 1000 Faculty Rev-912. doi:10.12688/f 1000 research.8010.1PMC 487687827274814 · doi ↗ · pubmed ↗
- 4Finer JJ. 2010. Plant nuclear transformation. In Kempken F, Jung C (ed), Genetic modification of plants. biotechnology in agriculture and forestry. Vol. 64. Springer, Berlin, Heidelberg.
- 5Li YG, Christie PJ. 2018. The Agrobacterium Vir B/Vir D 4 T 4SS: mechanism and architecture defined through in vivo mutagenesis and chimeric systems. Curr Top Microbiol Immunol 418:233–260. doi:10.1007/82_2018_9429808338 PMC 7011205 · doi ↗ · pubmed ↗
- 6Aguilar J, Zupan J, Cameron TA, Zambryski PC. 2010. Agrobacterium type IV secretion system and its substrates form helical arrays around the circumference of virulence -induced cells . Proc Natl Acad Sci USA 107:3758–3763. doi:10.1073/pnas.091494010720133577 PMC 2840527 · doi ↗ · pubmed ↗
- 7Geddes-Mc Alister J, Prudhomme N, Gutierrez Gongora D, Cossar D, Mc Lean MD. 2022. The emerging role of mass spectrometry-based proteomics in molecular pharming practices. Curr Opin Chem Biol 68:102133. doi:10.1016/j.cbpa.2022.10213335378382 · doi ↗ · pubmed ↗
- 8Prudhomme N, Pastora R, Muselius B, Mc Lean MD, Cossar D, Geddes-Mc Alister J. 2021. Exposure of Agrobacterium tumefaciens to agroinfiltration medium demonstrates cellular remodelling and may promote enhanced adaptability for molecular pharming. Can J Microbiol 67:85–97. doi:10.1139/cjm-2020-023932721220 · doi ↗ · pubmed ↗