Characterization of four cluster A1 Mycobacterium phage genomes, Applejack, Hermia, LilBib, and QTRlifeCrisis
Noah E. Adelman, Leah Cromarty, Sonja M. Francis, Vejune E. Griciute, Jason N. Hart, Abigail Hoffpauir, Ryan Koss, Ronit Kumar, Melisa N. Matonsi, Emma Morrison, Grace Paiement, Autumn Perley, Adrian Plichta, Liliana Rodrigues, Melody N. Neely, Caitlin S. Wiafe-Kwakye

TL;DR
This paper describes four new Mycobacterium phage genomes and their unique genetic features.
Contribution
The study characterizes four new cluster A1 mycobacteriophages with unique enzymatic features.
Findings
Four new siphoviruses (Applejack, Hermia, LilBib, QTRlifeCrisis) were isolated and sequenced.
Applejack and LilBib encode metallophosphatases with inteins.
Applejack contains a DNA methylase with an out-of-frame HNH endonuclease.
Abstract
Four novel cluster A1 siphoviruses, Applejack, Hermia, LilBib, and QTRlifeCrisis, were isolated on Mycobacterium smegmatis. Their complete genome sequences are 49,633–50,325 bp in length and encode 80–91 protein-coding genes. Applejack and LilBib encode metallophosphatases containing inteins, and Applejack encodes a DNA methylase that contains an out-of-frame HNH endonuclease.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
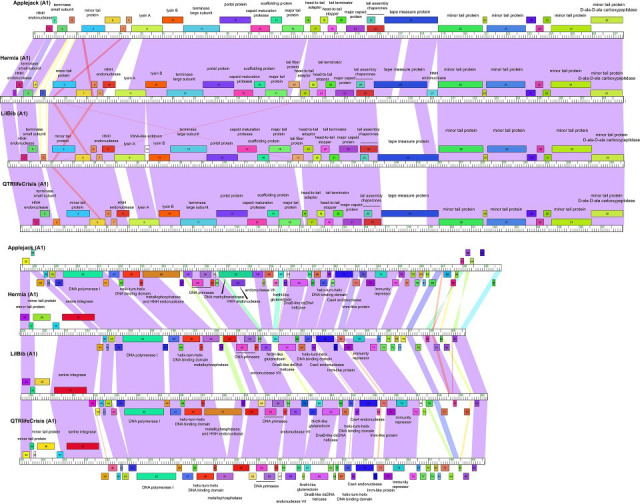 Fig 1
Fig 1| Isolation data | Applejack | Hermia | LilBib | QTRlifeCrisis |
|---|---|---|---|---|
| Soil GPS coordinates | 44.898789 N, 68.666355 W | 44.896201 N, 68.672347 W | 44.5549 N, 68.4206 W | 44.896201 N, 68.672347 W |
| Soil isolation location | Orono, ME | Orono, ME | Orono, ME | Old Town, ME |
| Isolation procedure | Enrichment | Enrichment | Direct | Direct |
| Rounds of plaque assays to purify plaques | 3 | 4 | 6 | 5 |
| Plaque morphology | 2–7 mm diameter, clear center turbid ring | 4 mm diameter, turbid | 4 mm diameter, turbid | 2–4 mm, clear center turbid ring |
| Particle morphology and average particle dimensions (± the standard error) | ||||
| Morphology | Siphoviridae | Siphoviridae | Siphoviridae | Siphoviridae |
| Tail length (nm) | 167 ± 2.03 | 171 ± 3.11 | 201 ± 2.26 | 199 ± 2.12 |
| Capsid diameter (nm) | 69.7 ± 0.724 | 72.3 ± 0.753 | 69.6 ± 1.01 | 72.4 ± 0.733 |
| Number of particles measured | 5 | 5 | 5 | 5 |
| Sequencing and genome characteristics | ||||
| Genome length | 49,791 | 49,670 | 50,325 | 49,633 |
| GC content | 63.5% | 63.9% | 63.8% | 63.7% |
| Number 250 bp single-end reads | 191,866 | 192,615 | 191,557 | 193,004 |
| Fold coverage | 708 | 836 | 625 | 928 |
| Number of protein-coding genes | 91 | 84 | 80 | 86 |
| Number of tRNAs | 0 | 0 | 0 | 0 |
- —National Institutes of Healthhttp://dx.doi.org/10.13039/100000002
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBacteriophages and microbial interactions · Plant and Fungal Interactions Research · Genomics and Phylogenetic Studies
ANNOUNCEMENT
Actinobacteriophage are diverse viruses that infect Actinobacteria, including non-pathogenic and pathogenic Mycobacterium species (1–6). Isolating and characterizing novel phages provides insight to phage evolution and opportunities to improve treatments for antibiotic-resistant mycobacterial infections (5, 7). Phages Applejack, Hermia, LilBib, and QTRlifeCrisis were isolated on the host Mycobacterium smegmatis MC^2^155 using direct and enrichment isolation procedures (Table 1). Soil extracts were prepared in 7H9 top agar, incubated for 1–2 h at 30°C and filtered on a 0.22-µM filter. For direct isolations, soil extracts were inoculated into M. smegmatis, incubated for 15 min and plated in 7H9 top agar onto L-agar plates. For enrichment isolations, soil extracts were inoculated with M. smegmatis, incubated for 2 d at 30°C with shaking before filtering onan 0.22-µM filter. Serially diluted extracts were plated with M. smegmatis and 7H9 top agar onto L-agar plates. Plates were incubated for 2daysd at 30°C and screened for plaques. Multiple rounds of standard plaque assays were performed to purify plaques (Table 1) (8). Hermia and LilBib consistently formed 4-mm turbid plaques. QTRlifeCrisis and Applejack plaques vary in size and have clear centers with turbid rings (Table 1). Transmission electron microscopy revealed particle morphologies consistent with Siphoviridae (Table 1).
Phage genomic DNA was isolated from high-titer lysates by phenol chloroform extraction (9) and prepared for sequencing using the Kapa Hyper Plus DNA library kit (Roche, South San Francisco, CA). Genomes were sequenced on an Illumina NovaSeq 6000 platform, and fold coverage is reported in Table 1. Newbler v.2.9 and Consed v29 were used to assemble genomes and check for completeness (10–12). The four genomes were 49,633–50,325 bp in length, have 63% GC content, and genome ends defined by 10 bp (CGGATGGTAA) 3′ overhangs (Table 1). They share >35% gene content with other cluster A mycobacteriophages in the Phamerator database (Actino_Draft v.589), and were assigned to subcluster A1 (2, 13).
Genomes were auto-annotated using Glimmer v.3.02 and GeneMark v.2.5 embedded in DNAMaster v.5.23.6 and PECAAN (https://blog.kbrinsgd.org/) (14, 15). Translational starts were manually determined based on inclusion of coding potential and conservation across homologs using Starterator (http://phages.wustl.edu/starterator/), BLASTP, and GeneMark.hmm output (14, 16). Putative gene functions were predicted using BLASTP and HHpred (16, 17). Transmembrane domains were predicted using Deep TMHMM (18). No tRNAs were detected using Aragorn v.1.2.38 and tRNAScan-SE2.0 (19, 20). The genomes encode 80–91 protein-coding genes (Table 1). Left arms encode forward-transcribed structural and assembly genes (Fig. 1). Right arms encode reverse-transcribed genes including DNA polymerase I, an immunity repressor, and overlapping DNA primases (Fig. 1). Hermia, LilBib, and QTRlifeCrisis encode a serine integrase that is absent in Applejack.
Genome map of cluster A1 Mycobacterium phages Applejack, Hermia, LilBib, and QTRlifeCrisis. The genome coordinates are represented by the ruler in units of kilobase pairs. The colored boxes above and below the ruler represent genes transcribed in the forward and reverse directions, respectively, and gene or gene product (gp) number is indicated within the box. Genes were assigned to a family using Phamerator in the Actino_draft database, and different families are indicated by colors (13). Colored shading between the genomes indicates nucleotide identity with violet of the color spectrum indicating the highest nucleotide identity. Predicted functions are centered above or below forward and reverse transcribed genes, respectively.
All the genomes encode multiple homing endonucleases and metallophosphatases. However, Applejack and LilBib metallophosphatases (gp48 and 51) contain an intein with high probability HHpred alignment to the Pyrococcus furiosis intein PI-Pfu (Fig. 1) (21). This intein includes a HINT domain for protein splicing and a homing endonuclease domain with two LAGLIDADG motifs, components of the active site (21, 22). The function of the intein is unknown but potentially provides a mechanism of post-translational regulation (23). Applejack also has a DNA methylase (gp54) with an out-of-frame HNH endonuclease (gp55). An HNH endonuclease is found upstream of the small subunit terminase in all genomes and likely plays a role in genome packaging (24).
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Pope WH, Mavrich TN, Garlena RA, Guerrero-Bustamante CA, Jacobs-Sera D, Montgomery MT, Russell DA, Warner MH, Hatfull GF. 2017. Bacteriophages of Gordonia spp. Display a spectrum of diversity and genetic relationships. m Bio 8:4. doi:10.1128/m Bio.01069-17PMC 555963228811342 · doi ↗ · pubmed ↗
- 2Pope WH, Jacobs-Sera D, Russell DA, Peebles CL, Al-Atrache Z, Alcoser TA, Alexander LM, Alfano MB, Alford ST, Amy NE, et al.. 2011. Expanding the diversity of mycobacteriophages: insights into genome architecture and evolution. P Lo S One 6:e 16329. doi:10.1371/journal.pone.001632921298013 PMC 3029335 · doi ↗ · pubmed ↗
- 3Pope Welkin H, Anders KR, Baird M, Bowman CA, Boyle MM, Broussard GW, Chow T, Clase KL, Cooper S, Cornely KA, et al.. 2014. Cluster M mycobacteriophages Bongo, Peg Leg, and Rey with unusually large repertoires of t RNA isotypes. J Virol 88:2461–2480. doi:10.1128/JVI.03363-1324335314 PMC 3958112 · doi ↗ · pubmed ↗
- 4Jacobs-Sera D, Abad LA, Alvey RM, Anders KR, Aull HG, Bhalla SS, Blumer LS, Bollivar DW, Bonilla JA, Butela KA, et al.. 2020. Genomic diversity of bacteriophages infecting Microbacterium spp. P Lo S One 15:e 0234636. doi:10.1371/journal.pone.023463632555720 PMC 7302621 · doi ↗ · pubmed ↗
- 5Hatfull GF. 2022. Mycobacteriophages: from petri dish to patient. P Lo S Pathog 18:e 1010602. doi:10.1371/journal.ppat.101060235797343 PMC 9262239 · doi ↗ · pubmed ↗
- 6Cresawn SG, Pope WH, Jacobs-Sera D, Bowman CA, Russell DA, Dedrick RM, Adair T, Anders KR, Ball S, Bollivar D, et al.. 2015. Comparative genomics of cluster O mycobacteriophages. P Lo S One 10:e 0118725. doi:10.1371/journal.pone.011872525742016 PMC 4351075 · doi ↗ · pubmed ↗
- 7Dedrick RM, Guerrero-Bustamante CA, Garlena RA, Russell DA, Ford K, Harris K, Gilmour KC, Soothill J, Jacobs-Sera D, Schooley RT, Hatfull GF, Spencer H. 2019. Engineered bacteriophages for treatment of a patient with a disseminated drug-resistant Mycobacterium abscessus. Nat Med 25:730–733. doi:10.1038/s 41591-019-0437-z 31068712 PMC 6557439 · doi ↗ · pubmed ↗
- 8Poxleitner M, Pope W, Jacobs-Sera D, Sivanathan V, Hatfull G. 2018. Phage discovery guide. Howard Hughes Medical Institute, Chevy Chase, MD.