Exploring the vector potential of Lipoptena cervi (Diptera: Hippoboscidae): first record of Setaria cervi (Filarioidea: Onchocercidae) DNA in deer keds
Klaudia Mária Švirlochová, Dana Zubriková, Veronika Blažeková, Lucia Vargová, Eva Čisovská Bazsalovicsová, Ján Čurlík, Ivana Heglasová, Bronislava Víchová

TL;DR
This study found evidence that deer keds may carry a parasitic worm, suggesting they could help spread it between animals.
Contribution
First molecular evidence of Setaria cervi DNA in Lipoptena cervi deer keds.
Findings
Two L. cervi from red deer in eastern Slovakia carried S. cervi DNA.
One red deer liver sample also tested positive for S. cervi DNA.
Abstract
Setaria cervi is a filarial nematode that infects both wild and domestic ungulates. It is primarily transmitted by mosquitoes, although the possible role of other hematophagous insects is still uncertain. We investigated 83 wingless deer keds (Lipoptena spp.) from red and fallow deer in northern and eastern Slovakia, as well as 43 red deer liver samples for the presence of filarial DNA. Deer keds were identified as Lipoptena cervi (n = 80) and Lipoptena fortisetosa (n = 3). Genomic DNA from individual ectoparasites was screened by PCR targeting a mitochondrial cox1 gene fragment of filaroid nematodes. Two L. cervi from red deer in Hrabušice (eastern Slovakia) tested positive for S. cervi DNA, with 100% sequence identity with worms recently isolated from Slovak game animals. Additionally, S. cervi DNA was detected in one liver from a red deer in the Vranov nad Topl’ou district. This…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1 Figure 2
Figure 2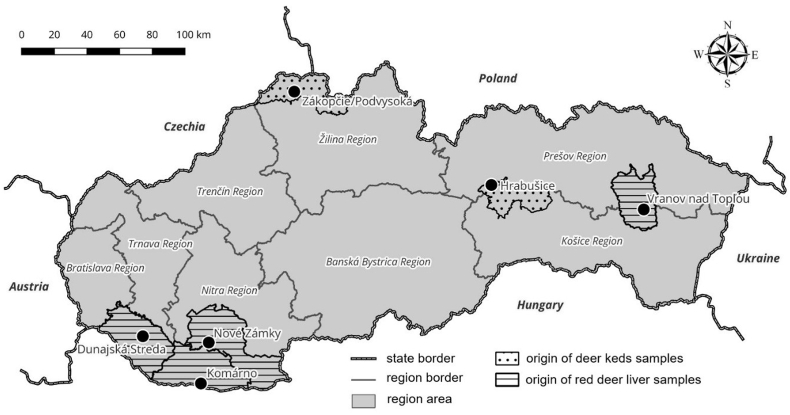 Figure 3
Figure 3Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBartonella species infections research · Coccidia and coccidiosis research · Insects and Parasite Interactions
Introduction
1
The filarioid nematode Setaria cervi (Rudolphi, 1819) (Filarioidea: Onchocercidae) is a vector-borne parasite that commonly parasitizes various ungulate hosts, mainly members of the families Cervidae and Bovidae, residing in the peritoneal or abdominal cavity (Anderson, 2000; Sundar and D’Souza, 2015). Although S. cervi is usually found in wild and domestic ruminants, it has also been detected in atypical hosts, such as sheep and goats (Lanková et al., 2021).
In Europe, S. cervi was first documented in Germany in red deer (Cervus elaphus) (Schwangart, 1940), with subsequent reports confirming its occurrence in red deer, sika deer (Cervus nippon), roe deer (Capreolus capreolus), and European moose (Alces alces) across several central and southern European countries (Blažek et al., 1968; Sugár, 1978; Shimalov and Shimalov, 2003; Rehbein and Visser, 2007; Kuzmina et al., 2010; Alasaad et al., 2012; Angelone-Alasaad et al., 2016; Oloś et al., 2019; Lanková et al., 2021). In Slovakia, S. cervi was only recently identified in red deer by Lazár et al. (2024).
Although Setaria spp. infections in wild cervids are usually asymptomatic and considered harmless, they can occasionally cause peritonitis, which is potentially life-threatening (Lazár et al., 2024). Additionally, if the parasites reach nervous tissues, they can cause locomotion problems and further paraplegia in the affected animals (Taylor et al., 2015).
Setaria spp. are transmitted by the blood-feeding insects, mainly mosquitoes (Diptera: Culicidae) (Oloś et al., 2019). However, blackflies (Diptera: Simuliidae) and some muscid flies (Diptera: Muscidae) have also been proposed as potential vectors (Czajka et al., 2012; Demiaszkiewicz et al., 2015; Kemenesi et al., 2015; Čurlík et al., 2019; Rydzanicz et al., 2019), suggesting that these filarioids may lack strict vector specificity and may also develop in other insect species. Consequently, the broader range of potential vectors could facilitate their wider geographical spread and affect the transmission dynamics.
Infection of the insect vector happens when it ingests first-stage larvae (microfilariae, L1) during a blood meal on an infected host. Inside the insect, L1 larvae develop into the infective third-stage larvae (L3). This development is temperature-dependent and lasts for approximately 14 days (Laaksonen et al., 2009). Transmission to a new mammalian host occurs when L3 larvae are transmitted during the vector’s next blood meal. After transmission, the larvae initially localize in the connective tissue before migrating to the abdominal cavity of the definitive host, where they continue to develop and mature into adult males and females.
Deer keds (Lipoptena spp.) are essential ectoparasites of cervids, and in Slovakia, there are two documented species (L. cervi and L. fortisetosa) (Oboňa et al., 2019, 2022). Infestation levels of Lipoptena spp. on individual animals can be very high, with reports of 240 individuals on a red deer, and up to 1400 on a single moose. Co-infestations involving L. cervi and L. fortisetosa on the same deer host have also been documented (Klepeckienė et al., 2020). Adult deer keds (L. cervi and L. fortisetosa) are winged when searching for a host but shed their wings permanently upon landing, after which they only occasionally switch hosts (Härkönen et al., 2010; Gałęcki et al., 2021).
In recent years, increasing attention has been directed toward the vector potential of deer keds for bacterial pathogens. These ectoparasites have been shown to harbor a range of bacteria, including Anaplasma spp., Bartonella spp., Borrelia spp., and Coxiella-like endosymbionts (Peña-Espinoza et al., 2023; Pearson et al., 2025). Among these, vector competence has so far been experimentally demonstrated only for Bartonella spp. (Dehio et al., 2004; Korhonen et al., 2015; Lee et al., 2016). In addition to bacteria, several protozoan pathogens, such as Theileria luwenshuni, T. ovis, and Trypanosoma spp., have also been detected in Lipoptena spp. (Böse and Petersen, 1991; Lee et al., 2016; Werszko et al., 2020). In Slovakia, molecular evidence has linked L. cervi to A. phagocytophilum (Víchová et al., 2011), with additional pathogens potentially involved (Švirlochová et al., unpublished).
Although interest in the ecology and epidemiology of Lipoptena spp. is growing, their potential role as vectors of nematodes remains unexplored - even though infestations by deer keds in cervid hosts are typically very high. Given the limited mobility of wingless keds and the restricted location of adult S. cervi worms, the transmission of microfilariae by L. cervi is likely infrequent, yet still possible, and probably limited to mechanical transfer events. This study aimed to examine the presence of S. cervi DNA in Lipoptena spp. collected from wild cervids to assess their possible involvement in the transmission cycle of the filarioid nematode.
Materials and methods
2
Sampling of deer keds
2.1
During September 2024, adult deer keds (Fig. 1) were collected by hunters from the fur of legally hunted fallow deer (Dama dama, n = 1) and red deer (Cervus elaphus, n = 6) in Podvysoká/Zákopčie, northern Javorníky Mountains. Additionally, ectoparasites were collected from three red deer (one male and two females) shot during legal hunting in Hrabušice village (Spišská Nová Ves District), on the northern slopes of the Slovak Paradise in eastern Slovakia (Fig. 2). Deer keds were preserved in 70% ethanol and transported to the Institute of Parasitology, Slovak Academy of Sciences (IP SAS). The species and sex of ectoparasites were determined under a stereomicroscope using an identification key (Oboňa et al., 2022).Fig. 1ALipoptena cervi, dorsal view. BLipoptena fortisetosa, dorsal view. Images were captured using a stereomicroscope with an Opta-Tech camera and processed in Capture One 2.2.1; final stacking and editing performed in Helicon Focus 8 software. Magnification: 30× .Fig. 1. Fig. 2Map of Slovakia with sampling sites.Fig. 2
Animal samples collection
2.2
Additionally, genomic DNA was isolated from 43 liver samples from red deer legally hunted in the Vranov nad Topľou District of eastern Slovakia (n = 25) and in the southern Slovakia districts (Dunajská Streda, Komárno, Nové Zámky; n = 18) of the country (Fig. 2). These tissue samples were provided by hunters some time ago and stored at the IP SAS for further analysis of the presence of vector-borne pathogens. Subsequently, they were included in this study and molecularly screened for the presence of Setaria spp. DNA.
DNA extraction and PCR analysis
2.3
Before DNA extraction, wingless deer keds were rinsed three times in sterile distilled water for 1–2 min each to remove residual ethanol. The specimens were then placed on filter paper and air-dried for 10–15 min. The whole body of each deer ked was subsequently homogenized individually with a sterile disposable scalpel in a 1.5-ml tube. Genomic DNA was extracted using the NucleoSpin Tissue Kit (Macherey-Nagel, Düren, Germany) following the manufacturer’s instructions, and the extracts were stored at −20 °C until PCR analysis. Total DNA from deer tissue samples was isolated using the same kit and protocol.
All DNA samples (from deer keds and animal tissues) were screened for the presence of filarioid parasites by PCR amplification of a partial (*c.*300 bp long) fragment of the mitochondrial cytochrome c oxidase subunit 1 (cox1) gene using the primers Cbcox1F (5′-CGG GTC TTT GTT GTT TTT ATT GC-3′) and Cox1intR (5′-ATAAGTACGAGTATCAATATC-3′) designed initially by Casiraghi et al. (2001) and later applied by Latrofa et al. (2012), and Bezerra-Santos et al. (2022).
Amplified PCR products were purified and sequenced bidirectionally using the PCR primers. Nucleotide sequences were analyzed using MEGA 11 software (Tamura et al., 2021) and compared with data available in the GenBank database using Basic Local Alignment Search Tool (BLAST).
Results
3
A total of 83 adult wingless deer keds were collected. In particular, 40 deer keds were removed from red deer in Hrabušice and identified as L. cervi (10 males and 30 females). Similarly, 40 L. cervi (26 males and 14 females) were removed from red and fallow deer shot in Podvysoká/Zákopčie. Additionally, three keds (1 female, and 2 males) removed from hunted red deer were identified as L. fortisetosa.
Among the examined DNA samples, two L. cervi DNA samples tested positive for the presence of filarioid DNA (2.5%, 2 out of 80 L. cervi examined). Both samples originated from red deer shot in Hrabušice in eastern Slovakia. Sequence analyses of the partial cox1 (303 bp) gene revealed an identical sequence structure in both analyzed specimens, with no intraspecific variation. Comparison of the obtained sequences with data deposited in the GenBank database revealed 100% sequence identity with the partial cox1 of S. cervi (GenBank: PP198315, PP198852, PP198854, and PP208633), obtained from the greater omentum of the rumen of red deer from the Slanské Mountains in Slovakia (Lazár et al., 2024). The newly generated sequences of S. cervi were deposited in the GenBank database under the accession numbers PV918858 and PV918859.
Overall, S. cervi DNA was detected in 1 of 43 red deer liver samples (2.3%). The positive sample originated from the Vranov nad Topľov District. The S. cervi cox1 nucleotide sequence (PX063956) obtained from a red deer liver sample was 100% identical with both sequences obtained from deer keds and adult parasites previously isolated from red deer (Lazár et al., 2024).
Discussion
4
Deer keds are widely distributed throughout central and eastern Europe, with high prevalences in red, fallow, and roe deer (Kadulski, 1996). In the present study, filarial DNA of S. cervi was identified in two specimens of L. cervi (prevalence of 2.5%) collected from red deer in Hrabušice, eastern Slovakia. This represents the first molecular evidence of S. cervi in deer keds and raises the possibility that L. cervi may act either as an incidental host of this filarioid nematode or a mechanical vector. Adult S. cervi reside primarily in the peritoneal cavity of deer, while microfilariae are released into the circulating peripheral blood. This blood circulation makes microfilariae accessible to hematophagous ectoparasites and is a key step in the potential transmission of the parasite.
The observed prevalence in L. cervi is comparable to that of Setaria tundra in mosquitoes in Finland, which ranges from 0.5% to 2.5% (Laaksonen et al., 2009). While the mechanism of transmission remains unconfirmed, our findings add to recent reports of Setaria spp. infection in Slovak cervids, including a prevalence of 6.7% in the red deer population (Lazár et al., 2024).
In our study, S. cervi DNA was detected in one liver sample (2.3%) from a red deer collected in the Vranov nad Topl’ou District in eastern Slovakia. However, as the liver is not the most suitable tissue for detecting Setaria spp. microfilariae, we acknowledge that the prevalence of Setaria spp. infection in the examined samples might be significantly underestimated. This finding supports the notion that S. cervi is circulating among wild ungulate populations in Slovakia. Nevertheless, to accurately determine the infection rate in game animals from this region, future studies should include screening of blood samples from the specific cervid hosts.
Deer keds are primarily host-specific to cervids, but sporadic infestations in domestic animals (Dehio et al., 2004; Sokół and Gałęcki, 2017) and humans (Härkönen et al., 2009; Kortet et al., 2010; Shimizu et al., 2025) have been documented. The overlapping habitats with livestock and increasing human-wildlife interactions raise concerns about potential interspecific transmission of pathogens, particularly in areas such as forest pastures, where domestic cattle and deer coexist.
Interestingly, in neighbouring Austria, one L. cervi individual collected from a red deer featured a cox1 sequence of an unknown onchocercid nematode (Filarioidea), most similar (95.1%) to Mansonella perforata isolated from Sika deer (Cervus nippon) in Japan (GenBank: AM749265). This observation not only raises questions about the presence of Mansonella spp. or related species in European deer populations but also highlights the possibility that additional filarioid nematodes may circulate among cervids. Whether this onchocercid sequence truly belongs to M. perforata or represents another, previously unsequenced species of Mansonella remains to be clarified (Peña-Espinoza et al., 2023).
The findings suggest that Lipoptena spp. may play a broader role in the transmission dynamics of filarial nematodes than previously recognized. There is potential for Lipoptena spp. to harbour and possibly disseminate a wider diversity of filarial species. Moreover, with the frequent movement of domestic and wild animals across borders, the risk of spreading filarial infections should not be underestimated. While our results do not confirm the vector competence of L. cervi for S. cervi, they suggest that deer keds may facilitate the ecological maintenance or mechanical transmission of this parasite.
However, as whole-body homogenates of deer keds were used for molecular screening, it remains unclear whether the detected S. cervi DNA originated from developing larval stages (L1 to L3) within the insect tissues or merely from ingested microfilariae derived from the blood meal. In mosquitoes, Setaria spp. microfilariae migrate to the thoracic muscles, where they develop into infective L3 larvae that subsequently localize in the mouthparts and are transmitted during feeding (Laaksonen et al., 2009). Given the anatomical and behavioral differences between mosquitoes and deer keds, it is uncertain whether Lipoptena spp. can support a comparable developmental cycle of filarioid nematodes.
Nevertheless, repeated feeding and close physical contact among cervid hosts increase the likelihood of microfilarial transmission in residual blood meal through contaminated mouthparts or co-feeding. This mechanical route may have epidemiological relevance in densely populated host populations. The present study provides initial evidence linking L. cervi to S. cervi, alongside the detection of related filarial DNA in red deer in Slovakia, and underscores the importance of investigating deer keds as possible participants in the transmission dynamics of filarioid nematodes.
Conclusions
5
Our findings reveal a previously undocumented association between L. cervi and S. cervi. A combination of microscopic or histological examinations with molecular analyses will be essential to determine whether L. cervi serves as a biological vector or represents a mechanical or dead-end host for S. cervi. Although the mechanism of transmission remains unclear, this study highlights the importance of continued surveillance for filarial parasites in both arthropod vectors and their wildlife hosts. As interactions between humans, domestic animals, and wildlife intensify, adopting a One Health approach will be crucial for anticipating and mitigating emerging parasitic risks.
Ethical approval
Not applicable. The wild animals were legally hunted by professional hunters under Slovak legislation. Liver samples were obtained post-mortem in accordance with national ethical standards and legal regulations of the Slovak Republic. All samples were provided to the Institute of Parasitology, Slovak Academy of Sciences, for scientific examination. No animals were killed specifically for this study.
CRediT authorship contribution statement
Klaudia Mária Švirlochová: Investigation, Methodology, Resources, Writing – review & editing. Dana Zubriková: Investigation, Methodology, Validation, Writing – original draft. Veronika Blažeková: Investigation, Methodology, Writing – review & editing. Lucia Vargová: Investigation, Methodology, Writing – review & editing. Eva Čisovská Bazsalovicsová: Funding acquisition, Investigation, Methodology, Project administration, Writing – review & editing. Ján Čurlík: Validation, Resources, Writing – review & editing. Ivana Heglasová: Investigation, Methodology, Resources. Bronislava Víchová: Conceptualization, Data curation, Funding acquisition, Project administration, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing.
Funding
This work was supported by the projects APVV-22-0440, 10.13039/501100006109VEGA 2/0033/25, 10.13039/501100006109VEGA 2/0051/24, and SK-10.13039/501100022871SRB 23–0046.
Declaration of competing interests
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Alasaad S.Pascucci I.Jowers M.J.Soriguer R.C.Zhu X.-Q.Rossi L.Phylogenetic study of Setaria cervi based on mitochondrial cox 1 gene sequences Parasitol. Res.110201228128510.1007/s 00436-011-2486-121695569 · doi ↗ · pubmed ↗
- 2Anderson R.C.Nematode Parasites of Vertebrates: Their Development and Transmission 2000 CABI Wallingford, UK
- 3Angelone-Alasaad S.Jowers M.J.Panadero R.Pérez-Creo A.Pajares G.Díez-Baños P.First report of Setaria tundra in roe deer (Capreolus capreolus) from the Iberian Peninsula inferred from molecular data: epidemiological implications Parasites Vectors 9201652110.1186/s 13071-016-1793-x 27682456 PMC 5041566 · doi ↗ · pubmed ↗
- 4Bezerra-Santos M.A.Moroni B.Mendoza-Roldan J.A.Perrucci S.Cavicchio P.Cordon R.Wild carnivores and Thelazia callipaeda zoonotic eyeworms: a focus on wolves Int. J. Parasitol. Parasites Wildlife 17202223924310.1016/j.ijppaw.2022.03.005PMC 892450735309037 · doi ↗ · pubmed ↗
- 5Blažek K.DykováI.Páv J.The occurrence and pathogenicity of Setaria cervi Rud., in the central nervous system of deer Folia Parasitol.151968123130
- 6Böse R.Petersen K.Lipoptena cervi (Diptera), a potential vector of Megatrypanum trypanosomes of deer (Cervidae)Parasitol. Res.77199172372510.1007/BF 009286911805220 · doi ↗ · pubmed ↗
- 7Casiraghi M.Anderson T.J.Bandi C.Bazzocchi C.Genchi C.A phylogenetic analysis of filarial nematodes: comparison with the phylogeny of Wolbachia endosymbionts Parasitology 12220019310310.1017/s 003118200000714911197770 · doi ↗ · pubmed ↗
- 8Czajka C.Becker N.Poppert S.Jöst H.Schmidt-Chanasit J.Krüger A.Molecular detection of Setaria tundra (Nematoda: Filarioidea) and an unidentified filarial species in mosquitoes in Germany Parasites Vectors 520121410.1186/1756-3305-5-1422236560 PMC 3277475 · doi ↗ · pubmed ↗