Structural and Functional Basis for Biased Agonism at the 5-hydroxytryptamine 5A Receptor
Xiaoyu Zhang, Linshan Xie, Peipei Chen, Jingjing Yu, Xiaowen Tian, Lei Wang, Jiali Wei, Zhenhua Shao, Wei Yan, Zheng Xu

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
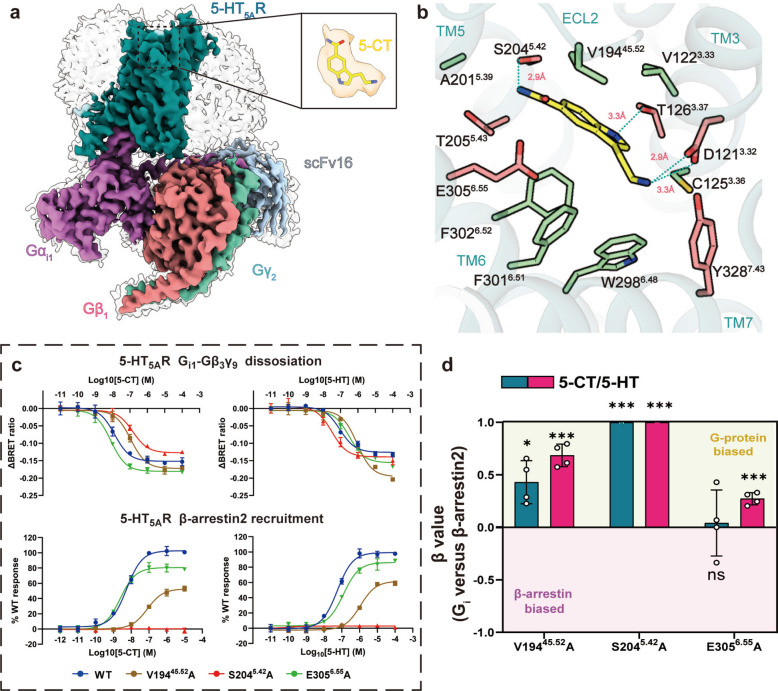 Figure 1
Figure 1- —National Key Research and Development Program of China
- —http://dx.doi.org/10.13039/501100001809National Natural Science Foundation of China
- —Sichuan Science and Technology Program
- —1.3.5 Project for Disciplines of Excellence, West China Hospital, Sichuan University
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsNeurotransmitter Receptor Influence on Behavior · Nicotinic Acetylcholine Receptors Study · Neuroscience and Neuropharmacology Research
Dear Editor,
The 5-hydroxytryptamine 5A receptor (5-HT_5A_R) belongs to the serotonin receptor family, which is classified as a group of G protein-coupled receptors (GPCRs) that are primarily associated with neuropsychiatric diseases. 5-HT_5A_R is widely expressed in the central nervous system, including the cerebral cortex, hippocampus, cerebellum, olfactory bulb and dentate gyrus. It is involved in a wild spectrum of neuropsychiatric functions, including cognition, memory, circadian rhythms, and antinociception effects [1]. 5-HT_5A_R predominantly signal through G_i/o_, and also recruit β-arrestin upon activation. Recent studies indicate that β-arrestin recruitment influence the efficacy of numerous drugs targeting serotonin receptors. Although several structures of inactive and active 5-HT_5A_R have been resolved, revealing interactions with several ligands as well as G_i/o_ proteins [2, 3], the role of 5-HT_5A_R-targeting drugs in β-arrestin signaling remains poorly understood. The synthetic agonist 5-carboxamidotryptamine (5-CT) is commonly used as a tool ligand to evaluate the function of serotonin receptors, particularly the 5-HT_5A_R, given its ten-fold higher affinity compared to 5-HT [4]. While 5-HT induces diverse signaling profiles across many other serotonin receptors, the biased agonism of 5-HT and 5-CT acting on 5-HT_5A_R remain unclear.
To understand the molecular mechanism of 5-HT_5A_R signaling through G_i_ and β-arrestin, we analyzed the structure of 5-CT stimulates 5-HT_5A_R/G_i1_ complex by cryo-electron microscopy (EM) with an overall resolution of 3.13 Å (Fig. S1a-b). The residues of 5-HT_5A_R and ligand were well defined in the density map, except for the intracellular loop 3 (ICL3) due to the high flexibility (Fig. 1a). By constructing the receptor residue-ligand atom contact matrix, we found 5-CT form tight hydrophobic interactions with C125^3.36^ and F301^6.51^ and salt bridge with D121^3.32^ (Fig. 1b). The amide group and the indole amine of 5-CT also interact with S204^5.42^ and T126^3.36^ by hydrogen bond (Fig. 1b). The binding pose of 5-CT exhibits slight conformational differences compared to that in a previous determined 5-CT/5-HT_5A_R/G_i_ complex structure, indicating heterogeneity of 5-CT/5-HT_5A_R interaction. This heterogeneity aligns with observations across multiple GPCR structures and likely captures near-native dynamics inherent to pharmacological mechanism of receptor-drugs interactions.Fig. 1. The structure and signaling characterization of 5-CT binding to 5-HT_5A_R. a Cryo-EM density map and atomic model of the 5-CT/5-HT_5A_R/G_i1_/scFv16 complex. (5-HT_5A_R: dark cyan; Gα_i1_: orchid; Gβ_1_: light coral; Gγ_2_: medium aquamarine; 5-CT: yellow). b The binding mode of 5-CT in the binding pocket of 5-HT_5A_R, the side chains of residues are shown as stick. c The effects of 5-CT and 5-HT on different mutants of the 5HT_5A_R. The response curves for 5-CT induced G_i1_ signaling (upper) and β-arrestin2 recruitment (lower) were measured by BRET and NanoBiT, respectively. d Bias factors (β value) of mutations relative to wild type (WT) calculated among three-agonist-induced receptor activation. Bias factors were derived from the curve fit parameters. Data are presented as mean ± SEM from four independent experiments, performed in triplicate. ∗ p < 0.033, ∗ ∗ p < 0.002 and ∗ ∗ ∗ p < 0.001 by one-way ANOVA followed by Dunnett’s multiple comparisons test.
In the contact matrix, S204^5.42^ and E305^6.55^ exhibit the highest contact frequencies with 5-CT (data not shown). To investigate the roles of these residues in 5-CT recognition, we introduced alanine substitutions at S204^5.42^ and E305^6.55^ into the receptor, following characterization of G_i1_ activation by bioluminescence resonance energy transfer (BRET) and β-arrestin2 recruitment by NanoBiT. S204^5.42^A abolished the β-arrestin2 recruitment and retained efficacy in G_i1_ dissociation (Fig. 1c), indicating that the polarity of 5.42 was essential for β-arrestin2 recruitment. Additionally, E305^6.55^A slightly enhanced the efficacy in G_i1_ signaling (ΔEmax = 0.229 ± 0.07, p = 0.03) and potency in the β-arrestin2 recruitment (ΔpEC_50_ = 0.452 ± 0.08, p = 0.006) of 5-CT. These results indicated that S204^5.42^ are essential for signaling bias of 5-CT stimulating 5-HT_5A_R, and interaction with E305^6.55^ may not be essential for 5-CT to function. Although V194^45.52^ did not form intense interaction with 5-CT, V194^45.52^A reduced potency for both G_i_ dissociation (ΔpEC_50_ = -0.841 ± 0.1, p < 0.001) and β-arrestin2 recruitment (ΔpEC_50_ = -1.151 ± 0.08, p < 0.001), indicating its importance in ligand binding. Given its strategic location on ECL2, this residue is hypothesized to function as a putative gatekeeper for the binding pocket. The loss of its bulky side chain is expected to accommodate easier ligand dissociation, thereby preventing sustained receptor activation.
We also characterized signaling of 5-HT_5A_R mutants activated by endogenous ligand 5-HT as reference. S204^5.42^A decreased potency of 5-CT (ΔpEC_50_ = -0.952 ± 0.12, p < 0.001), but enhanced potency of 5-HT in G_i_ dissociation (ΔpEC_50_ = 0.694 ± 0.05, p < 0.001). V194^45.52^A exhibited similar impacts on 5-CT and 5-HT, reducing potency of 5-CT (G_i_: ΔpEC_50_ = -0.841 ± 0.1, p < 0.001; β-arrestin2: ΔpEC_50_ = -0.952 ± 0.12, p < 0.001) and 5-HT (G_i_: ΔpEC_50_ = -1.151 ± 0.08, p < 0.001; β-arrestin2: ΔpEC_50_ = -1.206 ± 0.03, p < 0.001) in both pathways, whereas E305^6.55^A differently altered signaling of the two ligands (Fig. 1c). Specifically, E305^6.55^A slightly enhanced potency of 5-CT in β-arrestin recruitment (ΔpEC_50_ = 0.452 ± 0.08, p = 0.006), while it reduced potency of 5-HT in β-arrestin recruitment (ΔpEC_50_ = -0.262 ± 0.04, p = 0.01) thereby biasing signaling of 5-HT towards G_i_ dissociation (Fig. 1d). These findings confirmed that S204^5.42^ plays a crucial role in β-arrestin2 signaling by 5-HT_5A_R. In contrast, E305^6.55^ exhibits differential interaction modes with hydroxyl group of 5-HT compared to amide group of 5-CT. Mutagenesis and molecular dynamics simulations reveal distinct functional roles: D121^3.32^ (Energy = -41.5 ± 9.5 kcal/mol, 55.4% of total) and E305^6.55^ (Energy = -17.7 ± 10.8 kcal/mol, 23.6% of total) dominate 5-CT binding energetics, whereas S204^5.42^ facilitates β-arrestin recruitment through conformational rearrangement without significant binding energy contributions (Fig. S1c). Most serotonin receptors do not possess a negatively charged residue at position 6.55. Notably, the exceptions, 5-HT_1E_R and 5-HT_1F_R, exhibit low affinity for 5-CT. This correlation suggests that the presence of an acidic acid at 6.55 (E^6.55^) may negatively regulate 5-CT binding. Interestingly, in 5-HT_5A_R simulation, E305^6.55^ contributes a substantial favorable portion to the binding energy of 5-CT. We noticed that residues at 6.55 and 45.52 occupy roughly opposing positions. In 5-HT_1E_R and 5-HT_1F_R the amide-binding cavity is already constricted by the bulky I^45.52^, and the presence of E^6.55^ introduces steric clash. In contrast, V194^45.52^ in 5-HT_5A_R widens this cavity, allowing E305^6.55^ to form a productive hydrogen bond with the 5-carboxamide group of 5-CT, thereby playing a positive, selectivity-determining role.
Together, our results demonstrated that S204^5.42^A differentially affected the activation of downstream signaling by 5-CT and 5-HT, showing a clear β-arrestin2 signaling bias. The residue at 5.42 has been discovered contributing into the β-arrestin biased pharmacologic action of ligands in the serotonin receptors, as literatures reported [5]. The diversity of the molecular mechanism between serotonin receptors raised the possibility of designing selectively biased ligand for specific receptor in serotonin receptor family.
Supplementary Information
Supplementary Material 1.