In silico structural analysis of Oryza sativa RAD51 reveals key interactions for nucleoprotein filament assembly and regulation
Ayesha Azeem, Syed Farhat Ali, Rana Salman Anjum

TL;DR
This study uses computer modeling to understand how a rice protein called OsRAD51 interacts with DNA and another protein, OsBRCA2, to repair DNA damage.
Contribution
The study reveals structural details of OsRAD51 and how OsBRCA2 regulates its assembly into DNA repair filaments in rice.
Findings
OsRAD51 has conserved ATP- and DNA-binding sites and is regulated by phosphorylation and ubiquitination.
OsRAD51 interacts with OsBRC repeats via hydrophobic and polar interactions, regulating nucleoprotein filament assembly.
OsBRCA2 contains eight BRC repeats with a conserved motif similar to human BRC4, important for interaction with OsRAD51.
Abstract
Radiation sensitivity 51 (RAD51) is important for homologous recombination and DNA repair. The interaction between BRCA2 and RAD51 is crucial for the successful repair of DNA double strand breaks by homologous recombination. In the present study, through in silico analysis, we structurally characterized OsRAD51, a eukaryotic RAD51 ortholog from Oryza sativa Japonica A1 cultivar. Multiple sequence alignment showed the presence of conserved amino acids at the ATP- and DNA-binding sites. Several phosphorylation and ubiquitination sites were also predicted in OsRAD51 indicating its regulation by post-translational modifications. Structural modelling of OsRAD51 revealed two important regions at the protomer interface – one near the ATP-binding site (Walker A motif) and the other comprising of mainly hydrophobic residues. Polar and charge-charge interactions were noticeable at DNA-OsRAD51…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Fig 1
Fig 1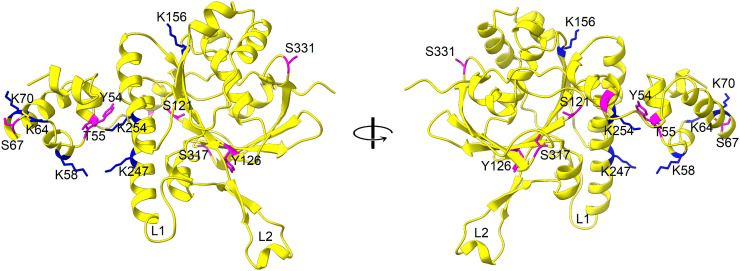 Fig 2
Fig 2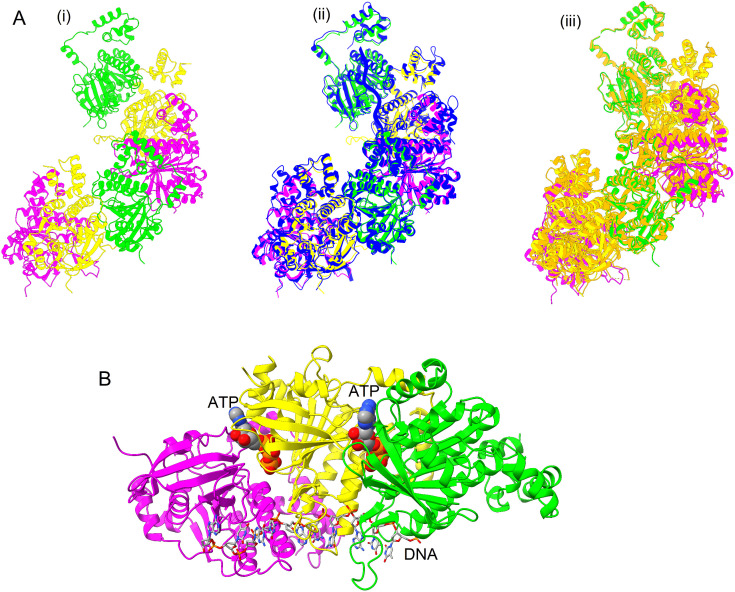 Fig 3
Fig 3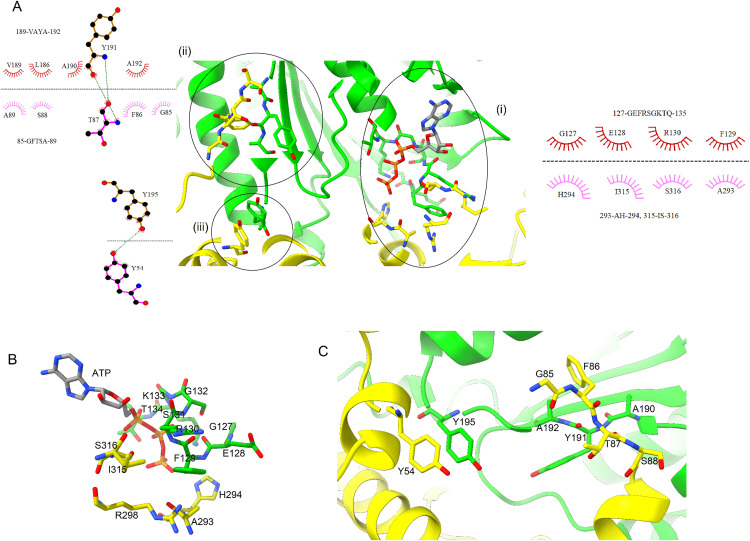 Fig 4
Fig 4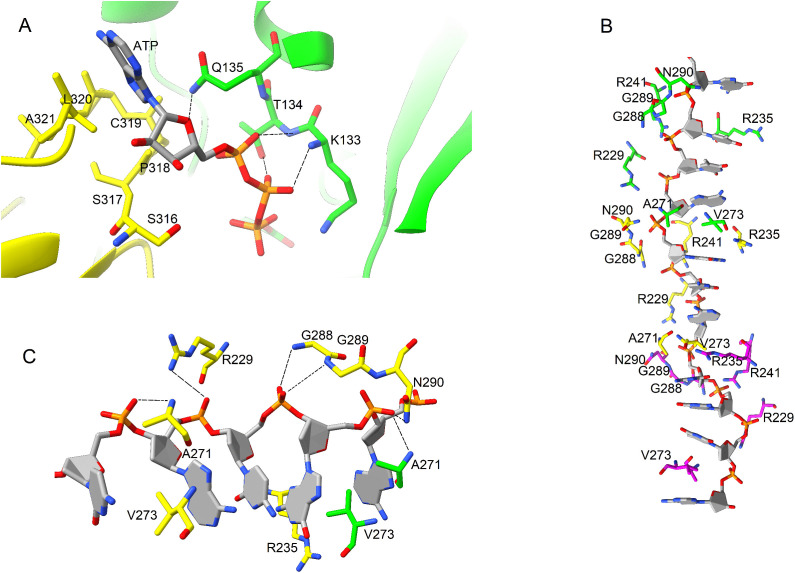 Fig 5
Fig 5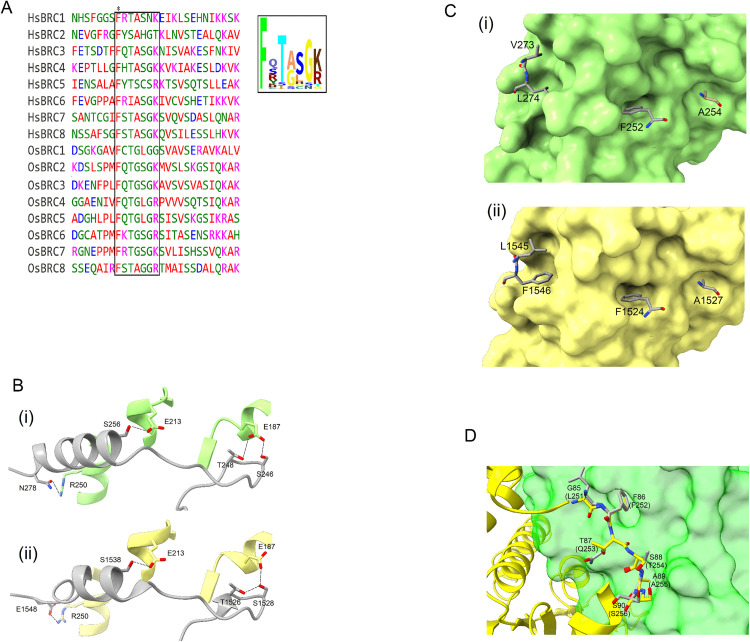 Fig 6
Fig 6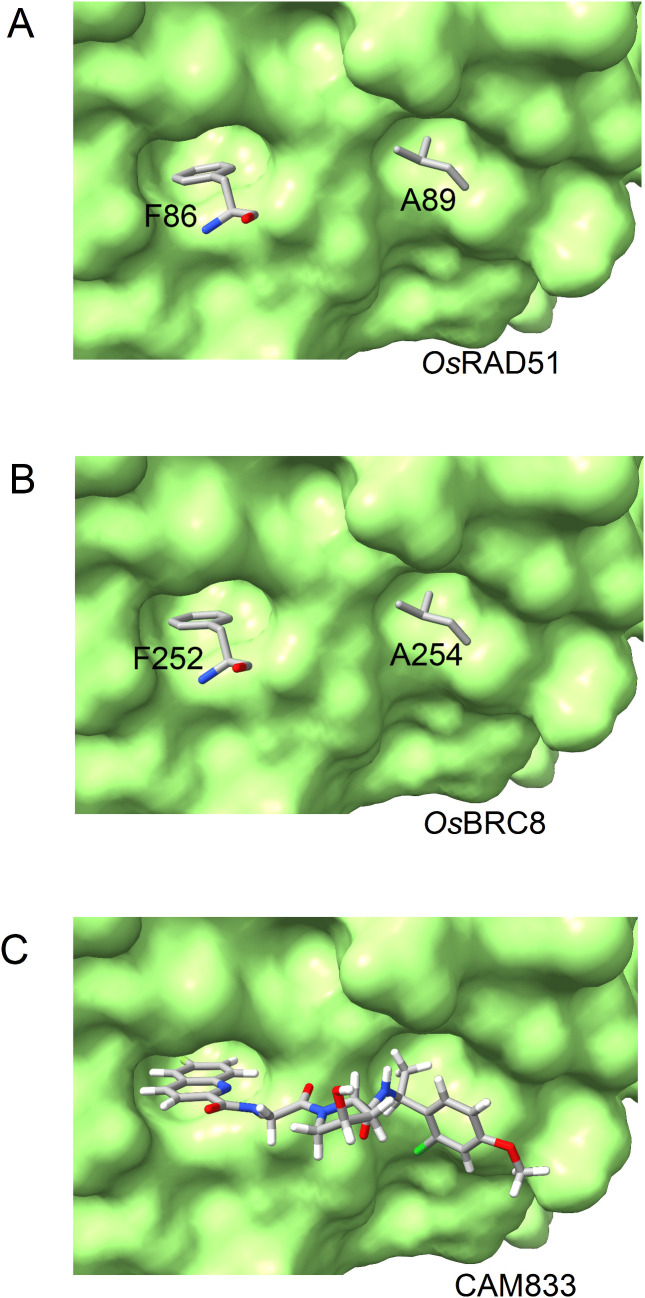 Fig 7
Fig 7- —http://dx.doi.org/10.13039/501100010221Higher Education Commision, Pakistan
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsDNA Repair Mechanisms · PARP inhibition in cancer therapy · Genetic Neurodegenerative Diseases
Introduction
Cellular DNA is under continuous stress through endogenous or exogenous agents. To tolerate such lesions in the DNA by faulty replication or due to ionizing radiations; nature has blessed cells with repair mechanisms to rehabilitate the damage [1]. As a result of DNA double strand break (DSB), homologous recombination (HR) repair pathway is the cell’s best choice which is active especially in the S phase of the cell cycle [2]. Despite its role in DNA repair in mitotic cells, HR is involved in creating genetic variability in meiotic cells [3]. Moreover, the phenomenon of HR can be applied for genome editing and genetic modifications in plants [4].
RAD51 is the eukaryotic homologue of RecA/RadA recombinases that perform a key step in the HR pathway in all three domains of life. Common feature of such recombinases is to assemble onto the sites of DNA double strand breaks to form helical nucleoprotein filaments which are the active species for homology search and strand exchange reaction [5]. RAD51 is comprised of two globular domains with its C-terminal domain homologous to the core ATPase domain of Escherichia coli (E. coli) recombinase A (RecA) which is structurally similar to the ATPase domain of F1 ATPase. The core ATPase domain of RAD51 participates in multimerization while its N-terminus is proposed to interact with DNA to make a stable nucleoprotein filament [6]. Recombinase activity of RAD51 is inevitably dependent upon interaction with other accessory factors, central to which is the interaction with breast cancer susceptibility protein type 2 (BRCA2) [7]. Despite its variable size among different organisms, it interacts with RAD51 through evolutionarily conserved BRC repeats. BRC repeats binding to RAD51 mimic the mechanism of RAD51 multimerization to regulate its loading and unloading at ds-ssDNA junction. As a result of loss of BRCA2 function at cellular level, the sensitivity to cross linking agents increases [8,9].
On a global scale, rice is a major staple food with a worldwide production of nearly 787 million tons in 2021 [10]. Meiotic recombination is essential for reproduction and seed formation. Oryza sativa RAD51 (OsRAD51) plays an important role in homologous recombination for DNA repair [11]. In O. sativa, several paralogs of RecA/Rad51 are present. Among these RAD51C and RAD51D are important for regulating crossover maturation during meiosis [12,13]. OsRAD51 mutant rice plants showed vegetative growth but were sterile – signifying its role in meiotic recombination [14]. In rice, another ortholog OsRAD51D promotes HR and inhibits non-homologous interactions thus ensuring recombination during rice meiosis [15]. Likewise, a RAD51 ortholog of Zea mays (ZmRAD51C) has been reported to be critical for DSB repair and meiosis in maize with ZmRAD51C mutations resulting in sterility [16]. In addition to the established role in DNA recombination and repair, RAD51 can contribute to resistance against diseases in plants. It was found that overexpression ZmRAD51A in rice and Arabidopsis improved disease resistance in these plants [17]. Interaction of BRCA2 with RAD51 is important for regulating DNA recombination and repair. It has been reported that O. sativa BRCA2 (OsBRCA2) is critical for loading of OsRAD51 to DSB during meiosis in rice [18].
Despite the advancements in elucidating and understanding the function of OsRAD51, no significant structural studies have been reported. To address this aspect, we modelled the structure of OsRAD51, identified important structural features and interactions including formation of nucleoprotein filament and interaction with BRC repeats.
Materials and methods
Sequence search and multiple sequence alignment
Homo sapiens RAD51 (HsRAD51) (Uniprot ID: Q06609) was used for a BLAST search against O. sativa genome. An ortholog OsRAD51A1 was identified (Uniprot ID: Q8SBB9) and used to perform multiple sequence alignment by using Clustal Omega.
The sequence of OsBRCA2 (accession number: Os01g0164800) was obtained from GeneBank [18]. The coding exons were analyzed by PHI–BLAST for the presence of BRC repeats (OsBRC).
Analysis of post-translational modification
In silico analysis of post translational modifications (phosphorylation and ubiquitination) of OsRAD51 was done by using PhosphoSVM [19], MuSuiteDeep [20], NetPhos3.1 [21] and GPS 6.0 [22]. Prediction of phosphorylation was done for serine, threonine and tyrosine residues of OsRAD51 and lysine residues were analyzed for ubiquitination. The sites detected by at least three of the prediction programs were selected for analysis.
Structural modelling and analysis
AlphaFold 3 was used to model the structure of OsRAD51 [23]. Models of monomeric OsRAD51 were generated using both “Use PDB template” and “Turn off template” settings. In addition, a homology model of OsRAD51 was also generated by MODELLER 10.5 [24] using HsRAD51 as a template (PDB ID 5H1B). Five models were generated and the best model was selected using the DOPE score.
To study the formation of nucleoprotein filament, an oligomeric model of OsRAD51 was generated by using AlphaFold 3 with “Turn off template” setting. Six OsRAD51 monomers and six ATP molecules were modelled with ssDNA (5’-GTTACGATGTCAGTACGTTAG-3’). To evaluate the effect of a previously reported inhibitor [8], CAM833 (PDB ID 6TW9) was docked into OsRAD51 model using Autodock Vina [25]. Structural analysis and comparison was done by using ChimeraX.
Results
Sequence comparison of eukaryotic RAD51 proteins
In pairwise alignment, OsRAD51 showed high sequence similarity to other eukaryotic orthologs including Zea mays RAD51 (94% identity), Arabidopsis thaliana RAD51 (86% identity), Saccharomyces cerevisiae RAD51 (55% identity) and HsRAD51 (69% identity) as shown in S1 Fig. In addition, multiple sequence alignment of OsRAD51 with human, yeast and other plant RAD51 orthologs revealed conservation of functionally important residues (Fig 1). The three main regions of HsRAD51 involved in protomer-protomer association [26] are also conserved in OsRAD51. These include the Walker A motif (which is also involved in ATP binding), a conserved aromatic interface (Y195 and Y54) and a short motif including F86 of OsRAD51. The loops L1 and L2 required for DNA interaction [27] are also conserved in OsRAD51 and other plant RAD51 orthologs. OsRAD51, like its human ortholog, contained two closely placed acidic residues namely D184 and E187 (acidic patch). In HsRAD51, this acidic patch is important for regulation of nucleoprotein filament assembly [28]. Taken together, these findings strongly suggest that OsRAD51can bind ATP and BRC repeats, and function in homologous recombination in a manner very similar to its eukaryotic counterparts (including HsRAD51).
Multiple sequence alignment of RAD51 homologs showed conserved sequence features of OsRAD51.Protomer-protomer interfaces, Walker A and B motif, and DNA interaction loops (L1 and L2) are indicated. (ScRad51, Saccharomyces cerevisiae RAD51 (Uniprot ID: P25454); HsRad51, Homo sapiens RAD51 (Uniprot ID: Q06609); AtRad51, Arabidopsis thaliana RAD51 (Uniprot ID: P94102); OsRad51, Oryza sativa RAD51 (Uniprot ID: Q8SBB9); ZmRad51, Zea mays RAD51 (Uniprot ID: Q67EU8). Predicted phosphorylation and ubiquitination sites of OsRAD51 are indicated by magenta and blue asterisks, respectively.
Post translational modifications of OsRAD51
RAD51 activity is regulated by post-translational modifications (PTMs) including both phosphorylation and ubiquitination [29]. So, to identify these PTMs, phosphorylation and ubiquitination sites were predicted in OsRAD51. It has been reported that ubiquitination of RAD51 hinders its interaction with BRCA2 [29]. Several ubiquitination sites were predicted in OsRAD51 including K58, K64, K70, K156, K247 and K254 (indicated by blue asterisks in Fig 1 and blue sticks in Fig 2). So, ubiquitination on these predicted sites can affect interaction of OsRAD51 with BRCA2 as well as its proteasome-mediated degradation [30]. Moreover, phosphorylation can also affect the activity of RAD51 [31]. Several potential phosphorylation sites were predicted in OsRAD51 (indicated by magenta asterisks in Fig 1 and magenta sticks in Fig 2) including Y54 (which forms the aromatic protomer-protomer interface), T55, S67, S90 (adjacent to protomer-protomer interface), S121, Y126 (adjacent to Walker A motif) S317 and S331. So, phosphorylation/dephosphorylation of these residues may influence the activity of OsRAD51 including ATP-binding and nucleoprotein filament assembly.
Predicted post-translational modification (PTM) sites in OsRAD51.OsRAD51 is represented as ribbon (yellow). Predicted phosphorylation sites are shown as magenta sticks and predicted ubiquitination sites as blue sticks. DNA-binding loops L1 and L2 are also shown.
Structural modelling and formation of nucleoprotein filament
Three types of monomeric OsRAD51 models were generated. Two models were generated with AlphaFold 3 using a template (“Use PDB template” setting) and without using a template (“Turn off template” setting). It was found that AlphaFold 3 selected HsRAD51 (PDB ID 5JZC) as a template when “Use PDB template” setting was used. A homology model of OsRAD51 was also generated by using MODELLER 10.5 with HsRAD51as a template. All three models were compared (S2 Fig) and backbone RMSD was calculated (S1 Table). All three models showed similar structure and domains (S2 Fig). Both AlphaFold models (using a template and without using a template) were very similar to each other with a backbone RMSD of 0.075. However, the model generated by MODELLER had a greater structural variability (with a backbone RMSD of nearly 1.0) when compared with both AlphaFold models. However, using HsRAD51 as a template may bias the resulting model. So, AlphaFold OsRAD51 model (generated without using a template) was used for further analysis and nucleoprotein filament modeling.
An AlphaFold model of nucleoprotein filament was generated (without using a template) with six monomers of OsRAD51, ssDNA and ATP (Fig 3A i). Superposition of OsRAD51 nucleoprotein filament model over ScRAD51 (PDB ID 9B2D, Fig 3A ii, shown in blue) and HsRAD51 (PDB ID 5H1B, Fig 3A iii, shown in orange) revealed significant structural similarity among the three nucleoprotein filaments.
Formation of nucleoprotein filament of OsRAD51.(A) i: Six protomers of OsRAD51 nucleoprotein filament (two sets of three protomers). The repeated protomers are colored in green, yellow and magenta. ii: Superposition of OsRAD51 nucleoprotein filament over ScRAD51 (PDB ID 9B2D, blue). iii: Superposition of OsRAD51 nucleoprotein filament over HsRAD51 (PDB ID 5H1B, orange) (B) Overview of OsRAD51 nucleoprotein filament structure. The protomers are shown in same colors as in A. ATP, shown as spheres, binds at the protomer-protomer interface. Bound ssDNA is shown as sticks.
Overall structure of OsRAD51 (3 protomers) with ssDNA and ATP is shown in Fig 3B. ATP is bound at the interface of the protomers with ssDNA surrounded by conserved loops L1 and L2.
Protomer-protomer interaction
Overall structure of OsRAD51 protomer-protomer interface is shown in Fig 4A. The protomers contact each other in two distinct regions. Firstly, the Walker A motif of first subunit of OsRAD51 (127-GEFRSGKTQ-135) interacts with a region near loop L2 of the second subunit (including A293, H294, R298, I315 and S316) (Fig 4A i). Interestingly, these regions are highly conserved between HsRAD51 and OsRAD51 except I315 and S316 of OsRAD51 which are replaced by Y315 and D316 in HsRAD51. Together, these regions of the two OsRAD51 subunits also form the ATP binding site (Fig 4B). Secondly, a short hydrophobic region of the first protomer (190-AYA-192) assembles with a hydrophobic region (85-GFTSA-89) of the second protomer (Fig 4A ii). In addition, another aromatic interaction is found near this site. Y195 of the first protomer interacts with Y54 of the second protomer (Fig 4A iii, 4C). These conserved interactions are found among the other neighboring protomers as well.
Protomer-Protomer interaction of OsRAD51.(A) Overview of protomer interaction. The interacting residues are shown as a 2D plot. ATP is shown as sticks at the Walker A motif. (B) Close-up view of Walker A motif interaction. (C) Hydrophobic interaction between the two protomers. (For residues shown as sticks: carbon atoms are colored according to the chain, nitrogen atoms blue and oxygen atoms red).
Interaction with ATP and DNA
As shown in Fig 5A, ATP is bound at the interface of the two protomers of OsRAD51. K133 and T134 of Walker A motif of the first protomer interact with the phosphate groups and Q135 with the ribose sugar. S315-A321 form the interacting surface of the second subunit. Moreover, OsRAD51 contained a conserved E163 located near AMP-PNP. In addition, D316 of HsRAD51 forms a salt bridge with ATP [32]. However, in OsRAD51 this is replaced by S316 at the ATP binding site. Likewise, AtRAD51 and ZmRAD51 also contain serine at the corresponding position (Fig 1). Collectively, these regions at the interface of the two protomers form the ATP binding site.
Interaction of OsRAD51 with ATP and DNA.(A) ATP binds at the interface of the two protomers (green and yellow) at Walker A motif. (B) Interaction of the protomers with DNA with loops L1 and L2 of each protomer surrounding the DNA. Important polar and charged amino acids of the protomers are shown. (C) Close-up view of OsRAD51 interaction with DNA triplets. Protomers interact with the backbone of DNA. Interacting residues of protomer 1 (green) and protomer 2 (yellow) are shown. (Interacting residues are marked with dashed lines. Residues represented as sticks are colored as in Fig 4).
The loops L1 and L2 (Fig 1) are important for DNA binding and strand exchange [27]. The model of OsRAD51 with DNA showed that the protomers mainly interact with the backbone of ssDNA with loops L1 and L2 surrounding DNA (Fig 5B). The nucleotides of bound ssDNA interact with protomers as triplets. As represented in Fig 5C, the valine residues of the protomers separate the triplets. A271 of first protomer and N290 of the second protomer interacts with the first phosphate; G288 and G289 interact with the second phosphate; and R229 with the third phosphate of the triplet.
Interaction with BRC repeat
BRCA2 is important for the recruitment and assembly of RAD51 [7]. PHI–BLAST was used to find BRC repeats in OsBRCA2. Similar to HsBRCA2, OsBRCA2 was found to contain 8 BRC repeats. Sequence alignment of HsBRC and OsBRC repeats showed the presence of a conserved motif with a notable phenylalanine. HsBRC repeats contained a conserved sequence FxxA. However, in OsBRC repeats, glycine was the predominant residue in this motif instead of alanine, i.e., FxxG (Fig 6A). To evaluate the effect of BRC repeats of OsBRCA2, models of OsRAD51 were generated separately with all 8 OsBRC repeats using AlphaFold. It has been reported that HsBRC3 and HsBRC4 bind to distinct regions of HsRAD51. HsBRC3 interacts with N-terminal domain whereas HsBRC4 binds to the nucleotide-binding core of HsRAD51 [33]. However, AlphaFold predicted all OsBRC repeats to bind to the same site at the nucleotide-binding core of OsRAD51 with the conserved FxxG/A motif occupying the protomer-protomer interface important for oligomerization of OsRAD51 (S3 Fig). So, based on the high sequence similarity of OsBRC8 to HsBRC4 (and presence of FxxA motif similar to HsBRC4), it was modelled with OsRAD51. It was found that OsBRC8 interact with OsRAD51 via both polar and hydrophobic interactions. R250 of OsRAD51 interact with N278 of OsBRC8, E213 with S256 and E187 with T248 and S246, respectively (Fig 6B i). Likewise, HsBRC4 showed a similar pattern of polar interactions with HsRAD51 (Fig 6B ii, PDB ID 1N0W). In addition, the non-polar residues (notably F252, A254 and L274) of OsBRC8 are buried into hydrophobic pockets of OsRAD51 (Fig 6C i) which is comparable to the non-polar interaction of HsBRC4-HsRAD51 (Fig 6C ii, PDB ID 1N0W). Overlay of OsBRC8 with OsRAD51 reveals that the former binds and blocks the non-polar interaction (Fig 4C) between OsRAD51 protomers. Fig 6D shows the conserved motif of second protomer (85-GFTSA-89, yellow) interacts with the short hydrophobic sheet (190-AYA-192, green) of the first protomer. The non-polar residues of OsBRC8 interact and mask this region. In particular, F252 of OsBRC8 resides in a cavity which is occupied by F86 of the second promoter – an interaction required for filament assembly [28]. As this phenylalanine is highly conserved in both HsBRC and OsBRC repeats (Fig 6A), OsBRC repeat may lead to the disassembly of OsRAD51.
Interaction of BRC repeat with OsRad51.(A) Sequence alignment of BRC repeats in HsRAD51 and OsRAD51 shows the presence of conserved Fxx(A/G) motif. A logo of this conserved motif is shown in the inset. (B) i: Polar contacts of OsBRC8 (grey) with OsRAD51 (green). ii: Polar contacts of HsBRC4 (grey) with HsRAD51 (yellow, PDB ID 1N0W). (C) i: Non-polar residues of OsBRC8 (grey) occupy the hydrophobic pockets of OsRAD51 (green surface). ii: Non-polar residues of HsBRC4 (grey) in the hydrophobic pockets of HsRAD51 (yellow surface, PDB ID 1N0W) (D) OsBRC8 (grey) binds to the protomer-protomer interface of OsRAD51. The interacting residues of second protomer (yellow) are shown as sticks. OsBRC8 residues are labelled in parenthesis. (residues represented as sticks are coloured as in Fig 4).
Interaction with CAM833
CAM833 has been shown to occupy the FxxA-binding pocket of HsRAD51 [8]. This binding may prevent the oligomerization of RAD51. To evaluate this effect, OsRAD51 was modelled with CAM833. It was observed that CAM833 binds the hydrophobic cavities of OsRAD51 required for its oligomerization via FxxA motif. During oligomerization, the Phe and Ala binding pockets of OsRad51 are occupied by F86 and A89 of the second OsRAD51 protomer, respectively (Fig 7A). OsBRC8 of BRCA2 (by its FxxA motif) can bind the same hydrophobic pockets (Fig 7B) and may prevent the oligomerization of OsRAD51. Likewise, CAM833 may interfere with OsRAD51 oligomerization site by binding these Phe and Ala cavities (Fig 7C).
Phe and Ala binding pockets of OsRAD51 (green) bound by (A) F86 and A89 of OsRAD51, (B) F1524 and A1527 of BRC repeat and (C) CAM833.
Discussion
RAD51 plays a significant role in HR and DSB repair pathway. Resection of the break point is followed by RAD51 recruitment and assembly as a nucleoprotein filament which invades a homologous region and mediates recombination [5]. Moreover, RAD51 is also important in preventing transcription-replication conflicts [34], stabilizing replication fork [35] and protecting abasic sites [36]. BRCA2 is a key regulator of RAD51 assembly with BRC repeats preventing RAD51 nucleoprotein filament formation while its C-terminal domain (TR2) stabilizing RAD51 binding to double strand DNA [37].
In this study, sequence and in silico structural analysis of OsRAD51 was done. Multiple sequence alignment of OsRAD51 with other plant, yeast and well-studied human (HsRAD51) orthologs was done. The analysis showed typical sequence features in OsRAD51. Walker A and Walker B motif, required for ATP-binding and hydrolysis [5], were identified in OsRAD51. In addition, Walker A motif also contributes to protomer-protomer interaction [26]. HsRAD51 contains two conserved acidic residues (D184 and D187). These are important for binding of TR2 domain of BRCA2. TR2 stabilizes RAD51 and prevent its disassembly [28,38]. Sequence analysis showed that OsRAD51 contained conserved acidic residues on the corresponding positions (D184 and E187) thus indicating a similar mechanism of nucleoprotein filament stabilization.
The activity of RAD51 is regulated by PTMs including ubiquitination and phosphorylation. Covalent attachment of ubiquitin and ubiquitin-like proteins mediate many regulatory processes including protein turnover [39]. It has been reported that RAD51 ubiquitination interferes with its interaction with BRCA2 [29]. Also, p53 upregulated modulator of apoptosis (PUMA) can facilitate ubiquitination of RAD51 thus repressing HR repair [40]. Several lysine residues of RAD51 have been shown to be important for its ubiquitination including K57, K58, K64, K107 and K156 [41]. We identified six lysine residues as potential ubiquitination sites in OsRAD51 (Fig 2). Among these, K58, K64 and K156 were identical to those of HsRAD51 as reported earlier [41]. Moreover, K57 and K107 of HsRAD51 corresponded to R57 and R107 in OsRAD51 (Fig 1) indicating these residues as potential ubiquitination sites in OsRAD51. In addition to ubiquitination, RAD51 activity can also be regulated by its phosphorylation. Several putative phosphorylation sites in OsRAD51 were identified. Some of these reside within or adjacent to the protomer-protomer interface and ATP-binding site. Y54 of OsRAD51 was identified as a possible phosphorylation site. This residue forms the aromatic protomer interface and interacts with Y195 of the next protomer. So, phosphorylation of Y54 is expected to hinder this hydrophobic interaction. This is also supported by the observation that phosphorylation of the corresponding residue in HsRAD51 resulted in inhibition of DNA-binding and strand exchange [42]. Another potential phosphorylation site of OsRAD51 is Y126 which lies adjacent to Walker A motif. Phosphorylation of the corresponding residue (Y77) of Deinococcus radiodurans RAD51 has been reported to increase its affinity for dsDNA [43]. S131 of OsRAD51 lies within the Walker A motif and is a predicted phosphorylation site. Phosphorylation of the corresponding residue (T131) of HsRAD51 was shown to result in DNA-independent ATPase activity [44]. In HsRAD51, the phosphorylation of T13 and S14 is important for its interaction with MRN complex [45]. However, in both multiple sequence alignment and phosphorylation-site prediction, corresponding residues were not detected in OsRAD51 or other plant RAD51 orthologs. So, this phosphorylation-based regulation might not occur in OsRAD51.
Several studies have described the structure of RAD51 nucleoprotein filament [28,46–48]. After forming a nucleoprotein filament and homologous DNA binding, RAD51 mediates strand exchange. We evaluated the formation of OsRAD51 nucleoprotein filament. AlphaFold was used to model OsRAD51 nucleoprotein filament. Conserved interacting residues of OsRAD51, as identified in Fig 1, were found at protomer-protomer interface (Fig 4). The protomers interacted via both polar and non-polar interactions. A conserved ATP binding site was also found at the interface of OsRAD51 protomers [49]. Recently, it was reported that S192 in Walker A motif of ScRAD51 has an important role in ATP hydrolysis. It is suggested that the side chain of S192 could interact with ATP and together with K191 can facilitate ATP hydrolysis [50]. In OsRAD51 these conserved positions of Walker A motif correspond to T134 and K133, respectively. Moreover, all plant and human orthologs in Fig 1 have a conserved threonine and lysine at these positions, respectively, indicating a similar conserved mechanism of ATP hydrolysis. RAD51 interacts with bound DNA through the conserved loops L1 and L2 leading to formation of a hetroduplex structure [27,51,52]. Likewise, our model of OsRAD51 showed that L1 and L2 surround the DNA and mainly interact with the DNA backbone (Fig 4B). The nucleotides of bound DNA interact with OsRAD51 as triplets (Fig 4C). A conserved valine in L2 (V273 of OsRAD51) inserts between the triplets to partition them. Similarly, a recent study reported that DNA nucleotides interact with ScRAD51 as triplets with a conserved V331 separating the nucleotide triplets. Moreover, R293 of ScRAD51, important for twisting and unstacking of DNA bases, was found to have multiple conformations. The dynamic nature of R293 may have an important role in inserting V331 to separate the bound DNA triplets [50]. In OsRAD51, the corresponding residue R235 was found to occupy a similar position close to the base triplets with V273 (corresponding to ScRAD51 V331) inserted between the triplets (Fig 5B).
HsBRCA2 contains 8 BRC repeats which are important for the regulation of HsRAD51. Among these, HsBRC3 and HsBRC4 have been shown to bind to distinct regions of HsRAD51 [33]. However, in our AlphaFold model of OsRAD51, all OsBRC repeats were predicted to bind to the same region (S3 Fig). This is probably because of the reason that all OsBRC repeats contain the FxxA/G motif with some conservation in the neighbouring region as well. And in the absence of a template, AlphaFold rely on sequence alignments to generate a model. So, this may have led to modelling all OsBRC repeats in the same region of OsRAD51. Moreover, BRCA2 contains two types of motifs which can regulate RAD51 assembly. The first one, FxxA motif (A-motif), is found in BRC repeats and prevent nucleoprotein filament assembly. The second, FxPP motif (P-motif), binds and stabilizes the nucleoprotein filament [53]. The FxxA motif of HsBRC repeat competes with HsRAD51 and prevents its oligomerization. Similarly, OsBRCA2 also contains 8 BRC repeats with a conserved phenylalanine (FxxG). Presence of this motif in OsBRC repeats indicates that OsBRC repeats can regulate OsRAD51 assembly in a manner similar to HsBRC repeats. In addition, HsBRCA2 contain two FxPP motifs. This protects the nucleoprotein filament against disruption by BRC repeats. FxPP interacts with loops 148-IDRGGGE-154 and 178-YGLS-181 of HsRAD51 [54]. OsRAD51 was found to contain corresponding sequences with conserved residues suggesting its regulation by OsBRCA2. This suggests that OsBRCA2 may contribute to both stability and disruption of OsRAD51 nucleoprotein filaments. Moreover, RAD51 has an intrinsic ability to oligomerize. This structural complexity makes it difficult to obtain RAD51 in monomeric form. One strategy recently reported was based on generation of a double mutant (F86E, A89E) in which the oligomerization interface is mutated and results in producing monomeric RAD51 [55]. However, oligomerization inhibitors may be used as an alternate strategy. CAM833 may inhibit oligomerization of RAD51 by occupying the F86 and A89 binding pockets [8]. To evaluate this, a model of OsRAD51 with CAM833 was generated. Our result suggests that CAM833 might block the oligomerization interface of OsRAD51 (Fig 7C). So, this approach may be useful in studying OsRAD51 in monomeric state. Moreover, this strategy may also be beneficial in studying HR and genetic stability in O. sativa. A recent report described the mechanism of strand exchange by HsRAD51 and identified several residues for D-loop formation. These include F279 in loop L2 (strand separation); R303, K304, R306, K313 (DNA capture); K39, K40, K64, K70 and K73 (arm duplex binding) [56]. Sequence alignment (Fig 1) showed that all these important residues are conserved in OsRAD51 as well.
So, taken together, these findings suggest that OsRAD51 may form nucleoprotein filament and can be regulated by OsBRCA2. OsRAD51 is essential for HR and meiosis in rice signified by the observation that OsRAD51 mutant rice plants are sterile [13,14]. In addition to these physiological roles, the DNA binding domain (DBD) of OsRAD51 can used to improve DNA editing systems. Although the physiological role of OsRAD51 is much explored, to our knowledge, no structural investigations have been done. Our study sheds light on structure and regulation of assembly of OsRAD51 providing structure-function relationship.
Supporting information
S1 FigPairwise alignment of OsRad51 with other eukaryotic homologs.(Percent identity is shown in brackets). ZmRad51, Zea mays RAD51, Uniprot ID: Q67EU8 (94%). AtRad51, Arabidopsis thaliana RAD51, Uniprot ID: P94102 (86%). ScRad51, Saccharomyces cerevisiae RAD51, Uniprot ID: P25454 (55%). HsRad51, Homo sapiens RAD51, Uniprot ID: Q06609 (69%).(PDF)
S2 FigSuperposition of OsRAD51 monomeric models generated by AlphaFold 3 and MODELLER10.5.AlphaFold OsRAD51 model without using template (cyan), AlphaFold OsRAD51 model using template (yellow) and homology model of OsRAD51 by using MODELLER (green). Loops L1 and L2 are shown.(PDF)
S3 FigSuperposition of modelled OsBRC repeats with OsRAD51 (green surface).Conserved phenylalanine and alanine/ glycine of FxxA/G motif are shown as sticks. OsBRC1 (red), OsBRC2 (yellow), OsBRC3 (cyan), OsBRC4 (magenta), OsBRC5 (wheat), OsBRC6 (grey), OsBRC7 (orange) and OsBRC8 (blue).(PDF)
S1 TableComparison of OsRAD51 models.(PDF)
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Carusillo A, Mussolino C. DNA Damage: From Threat to Treatment. Cells. 2020;9(7):1665. doi: 10.3390/cells 9071665 32664329 PMC 7408370 · doi ↗ · pubmed ↗
- 2Chatterjee N, Walker GC. Mechanisms of DNA damage, repair, and mutagenesis. Environ Mol Mutagen. 2017;58(5):235–63. doi: 10.1002/em.22087 28485537 PMC 5474181 · doi ↗ · pubmed ↗
- 3Hinch R, Donnelly P, Hinch AG. Meiotic DNA breaks drive multifaceted mutagenesis in the human germ line. Science. 2023;382(6674):eadh 2531. doi: 10.1126/science.adh 2531 38033082 PMC 7615360 · doi ↗ · pubmed ↗
- 4Chen H, Neubauer M, Wang JP. Enhancing HR Frequency for Precise Genome Editing in Plants. Front Plant Sci. 2022;13:883421. doi: 10.3389/fpls.2022.883421 35592579 PMC 9113527 · doi ↗ · pubmed ↗
- 5Bonilla B, Hengel SR, Grundy MK, Bernstein KA. RAD 51 Gene Family Structure and Function. Annu Rev Genet. 2020;54:25–46. doi: 10.1146/annurev-genet-021920-092410 32663049 PMC 7703940 · doi ↗ · pubmed ↗
- 6Sun Y, Mc Corvie TJ, Yates LA, Zhang X. Structural basis of homologous recombination. Cell Mol Life Sci. 2020;77(1):3–18. doi: 10.1007/s 00018-019-03365-1 31748913 PMC 6957567 · doi ↗ · pubmed ↗
- 7Kwon Y, Rösner H, Zhao W, Selemenakis P, He Z, Kawale AS, et al. DNA binding and RAD 51 engagement by the BRCA 2 C-terminus orchestrate DNA repair and replication fork preservation. Nat Commun. 2023;14(1):432. doi: 10.1038/s 41467-023-36211-x 36702902 PMC 9879961 · doi ↗ · pubmed ↗
- 8Scott DE, Francis-Newton NJ, Marsh ME, Coyne AG, Fischer G, Moschetti T, et al. A small-molecule inhibitor of the BRCA 2-RAD 51 interaction modulates RAD 51 assembly and potentiates DNA damage-induced cell death. Cell Chem Biol. 2021;28(6):835-847.e 5. doi: 10.1016/j.chembiol.2021.02.006 33662256 PMC 8219027 · doi ↗ · pubmed ↗