miR-28-5p and miR-708-5p Share a Common Seed with Different Functions in Lung Cancer Patients
Cristina Alexandra Ciocan, Cecilia Bica, Liviuta Budisan, Lajos Raduly, Sergiu Chira, Claudia-Cristina Burz, Ovidiu Farc, Antonia Harangus, Marioara Simon, Constantin-Ioan Busuioc, Stefan Strilciuc, Cornelia Braicu, Ioana Berindan-Neagoe

TL;DR
This study explores miR-28-5p and miR-708-5p in lung cancer, finding that they share a seed sequence but have different roles, with miR-708-5p showing potential for clinical use.
Contribution
The study reveals distinct functional roles of two miRNAs with a shared seed sequence in lung cancer.
Findings
miR-28-5p and miR-708-5p are overexpressed in advanced-stage lung cancer tissues.
miR-708-5p shows consistent overexpression and potential diagnostic value in lung cancer.
Both miRNAs regulate cancer-related pathways like ECM-receptor interaction and Hippo signaling.
Abstract
Lung cancer remains the leading cause of cancer-related mortality worldwide, accounting for nearly 1.8 million deaths annually. The present study aimed to investigate the role of miR-28-5p and miR-708-5p in lung cancer and to analyze the relationship between target gene profiles and transcriptional factor regulation. Both miRNAs that share a common seed sequence were found to be overexpressed in a cohort of 32 paired tumor and adjacent normal tissue samples collected from patients diagnosed at advanced stages (III and IV) of disease. Data from the dbDEMC database revealed that miR-28-5p exhibited variable expression across lung cancer subtypes, whereas miR-708-5p showed consistent overexpression, reinforcing its potential clinical diagnostic significance. Using the TransmiR database, we identified complex TF–miRNA regulatory networks, with both shared and distinct transcription factors…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
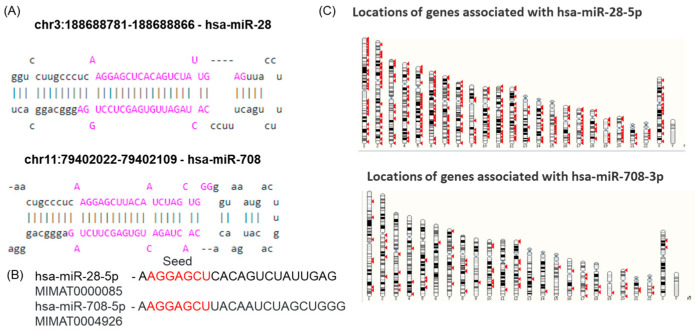 Figure 1
Figure 1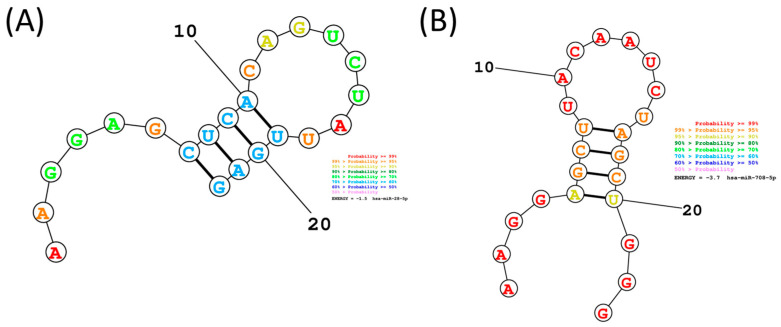 Figure 2
Figure 2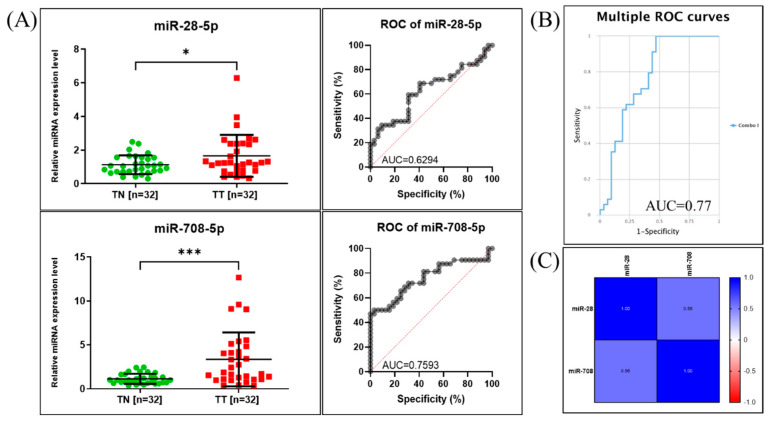 Figure 3
Figure 3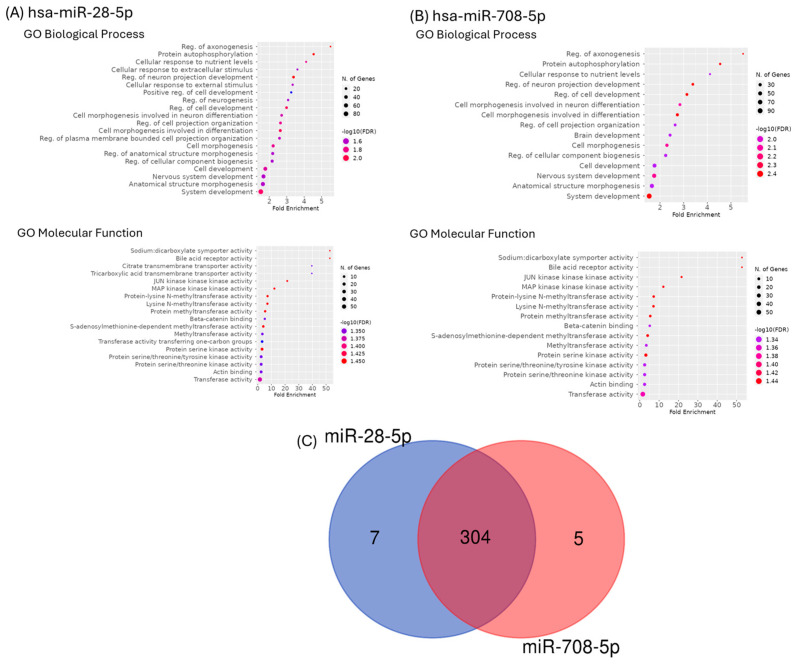 Figure 4
Figure 4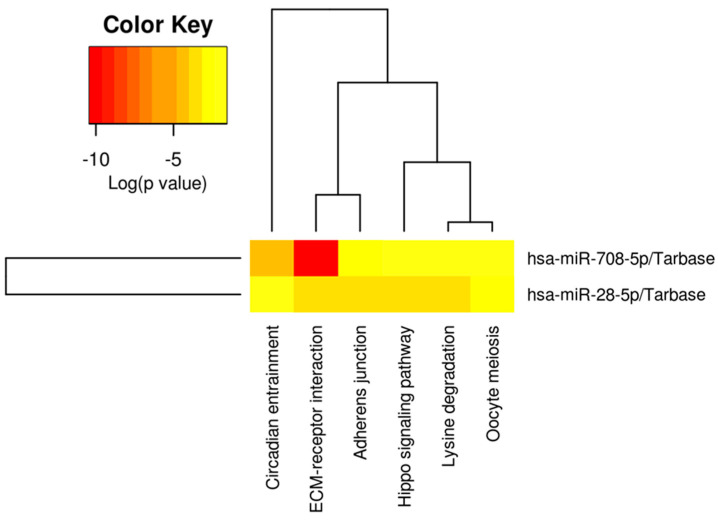 Figure 5
Figure 5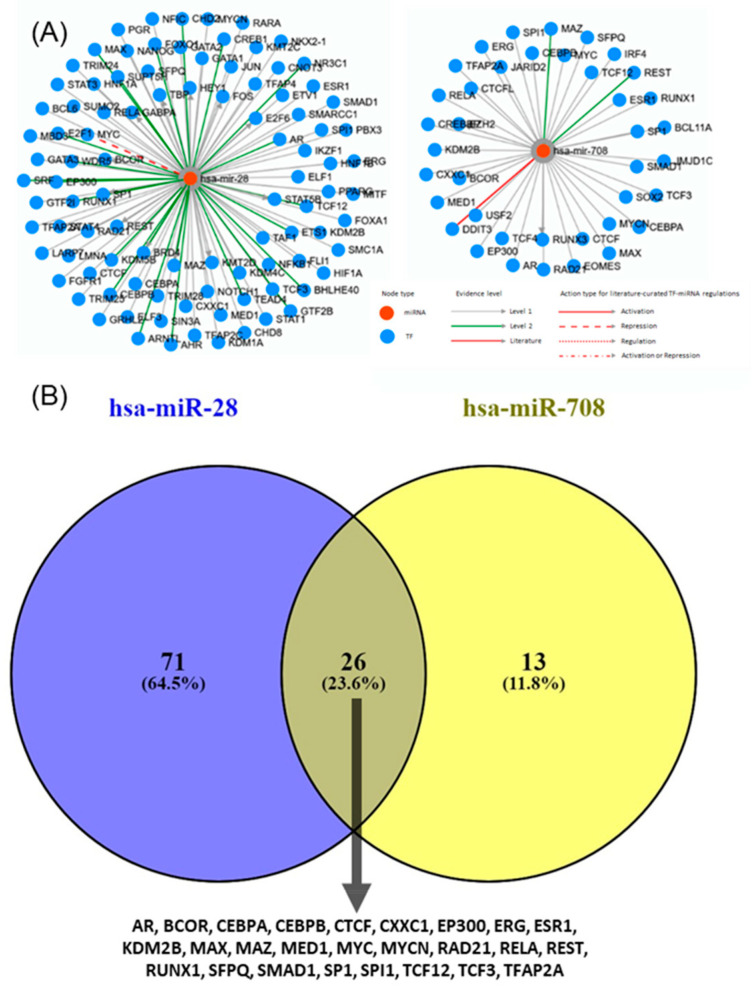 Figure 6
Figure 6- —Competitiveness Operational Program, 2014–2020
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMicroRNA in disease regulation · Hippo pathway signaling and YAP/TAZ · Circular RNAs in diseases
1. Introduction
Lung cancer includes three major histological subtypes: non-small cell lung cancer (NSCLC), which accounts for approximately 80–85% of cases, small cell lung cancer (SCLC), representing about 10–15%, and malignant pleural mesothelioma (MPM), a rare subtype contributing to less than 1% of primary lung malignancies. Meanwhile, the NSCLC was subclassified as lung adenocarcinoma (LUAD) and lung squamous cell carcinoma (LUSC). This condition is the leading cause of cancer-related deaths, with an estimated 1.8 million deaths due to late diagnosis [1]. Therefore, the mortality rate is very high, and the 5-year overall survival rate is only 15% [2]. Identifying novel molecular candidates with clinical utility to understand better this malignancy remains a key objective in clinical research and development. The pathogenesis of lung cancer is complex, involving both genetic and transcriptomic alterations [3].
MiRNAs are small non-coding RNAs, 19–24 nucleotides in length, that serve as critical regulators of gene expression through a post-transcriptional mechanism, thereby impacting various physiological and pathological cellular processes [4,5,6,7]. Mounting evidence has shown that miRNAs play various regulatory roles, depending on the target genes, acting as oncogenes or tumor suppressors [6,7,8,9,10]. Significant alterations of miRNA expression patterns in tumor tissue versus normal adjacent tissue make them potential molecule candidates with clinical utility [7,11]. The precise contribution of individual miRNAs remains incompletely understood [3,12,13,14,15,16,17], including in lung cancer [3,15,16,17].
miR-28 and miR-708 are key transcripts with a common seed sequence implicated in cancer biology [18,19]. MiR-28-5p has been proven to have emerging roles in cancers targeting critical pivotal factors related to cell proliferation, migration, invasion, and metastasis [20,21,22]. Similarly, miR-708-5p is dysregulated in various cancers, where it can be either overexpressed or underexpressed compared to normal adjacent tissue [23,24]. Therefore, miR-708-5p appears to have both tumor-suppressor and tumor-promoting functions [23,25].
This study investigated the roles of miR-28-5p and miR-708-5p in lung cancer, with a specific focus on their potential clinical applications. The novelty of this work lies in the comparative analysis of two miRNAs with same seed sequence, highlighting their differential expression patterns in lung cancer and the regulatory networks they influence. We evaluated their expression in paired NSCLC tumor and adjacent normal tissues and validated our findings with additional data from the dbDEMC database. Furthermore, we identified target genes, regulatory transcription factors networks, and miRNA-gene associated networks, highlighting signaling pathways regulated by these transcripts.
2. Results
2.1. miR-28-5p and miR-708-5p Share a Common Seed Sequence
Genomic mapping indicated that miR-28-5p is encoded on chromosome 3 within an intronic region of the LPP (LIM domain containing preferred translocation partner in lipoma) gene, whereas miR-708-5p is encoded on chromosome 11, in the intronic region of the gene TENM4 (teneurin transmembrane protein 4), as observed from the MIRIAD database. The stem-loop structures of these transcripts, downloaded from miRBase, are presented in Figure 1A.
The specific sequence region responsible for target recognition is called the “seed sequence”, which comprises 2–8 nucleotides at the 5′ end, as shown in Figure 1B for the two selected transcripts. Since miRNAs can regulate multiple target genes, their functional effectiveness depends not only on seed complementarity but also on interactions mediated by the non-seed region. Variations in this region can change target specificity and binding affinity, ultimately leading to different regulatory outcomes. Structural differences outside the seed region may also influence miRNA stability, accessibility, and interaction with mRNAs. Furthermore, the two miRNAs differ in both the number and chromosomal distribution of their predicted targets (Figure 1C), supporting the idea that, despite sharing a seed sequence, miR-28-5p and miR-708-5p likely have distinct biological roles.
The secondary structure prediction and analysis of miR-28-5p and miR-708-5p, using RNAstructure, provide insights into their thermodynamic stability, which is directly linked to their biological function. As can be observed from Figure 2, the free energy for miR-28-5p is −1.5, meanwhile for the case of miR-708-5p is −3.7; the secondary structure forms due to the folding of the sequence into paired and unpaired regions, contributing to its overall stability and potential interactions with target mRNAs, therefore miR-708-5p have a higher stability, it is expected that transcripts might have a context dependent functionality.
2.2. qRT-PCR for miR-28-5p and miR-708-5p Expression Assessment in NSCLC Patients
When analyzing the relative expression levels of the tested miRNAs, we observed increased levels for both miRNAs when comparing tumor to adjacent non-tumor tissue samples (Figure 3). The relative expression in tumor tissue was 1.625 ± 1.245 (p = 0.0311) for miR-28-5p and 3.351 ± 3.066 (p = 0.0002) for miR-708-5p.
To further assess their clinical relevance, we performed ROC curve analysis, which evaluates each miRNA’s ability to differentiate between tumor and non-tumor samples. The area under the curve (AUC) provides a quantitative measure of diagnostic accuracy, with values closer to 1.0 indicating better discriminatory power and a value of 0.5 indicating no discriminative ability. The analysis revealed that miR-28-5p had an AUC of 0.6294, suggesting modest discriminatory capacity, while miR-708-5p achieved an AUC of 0.7593, indicating a higher ability to distinguish tumor from non-tumor tissues.
The combined ROC curve, generated using the CombiROC online tool [26], displayed an AUC value of 0.77 (Figure 3B), demonstrating slighly improved diagnostic performance when the two miRNAs were analyzed together compared to individual assessment. Figure 3C shows the statistical correlation between the expressions of the two evaluated miRNAs.
2.3. Validation of the Expression Levels of miR-28-5p and miR-708-5p in Lung Cancer Using Public Available Datasets
The expression profiles of miR-28-5p and miR-708-5p in lung cancer, obtained from the dbDEMC database, are presented in Table 1 and Figure S1, supporting validation of laboratory data through external datasets [27]. The expression of miR-28-5p shows some variation across different lung cancer subtypes, suggesting potential context-dependent roles. In contrast, the expression of miR-708-5p appears consistent across lung cancer datasets, indicating a more uniform regulatory pattern.
2.4. Functional Classification and Enrichment Analysis of miR-28-5p and miR-708-5p Target Genes
GO enrichment approaches were used to identify the putative roles of the main targets for the selected two transcripts (Figure 4A,B). GO enriched categories in biological process groups were primarily linked to system development, anatomical structure morphogenesis, or nervous system development. GO enriched categories in molecular function were mainly related to transferase activity, actin binding, or protein serine/threonine kinase activity. A diagram Venn was generated, and we can observe 304 common genes (Figure 4C).
2.5. Pathway Analysis for miR-28-5p and miR-708-5p
The pathway and target analysis of miR-28-5p and miR-708-5p was performed using DIANA Tools mirPath v.3 (Table 2). In addition, Figure 5 represents the heatmap showing the pathway analysis for the two transcripts, revealing ECM–receptor interaction (Figure S2), adherence junctions (Figure S3), and Hippo signaling (Figure S4) as key elements.
While these analyses provide a comprehensive in silico overview of target gene networks and signaling pathways, we acknowledge that they remain predictive in nature. Experimental validation of target gene expression and pathway activity will be essential to confirm the functional impact of these findings. Nonetheless, this computational framework identifies clinically relevant candidates and prioritizes ECM–receptor interactions, adhesion molecules, and Hippo signaling as key mechanisms for future translational studies in lung cancer.
2.6. Transcription Factors and Regulatory Networks
TransmiR is a database that provides regulatory relationships between transcription factors (TFs) and miRNAs. Predicting the regulation of miR-28-5p and miR-708-5p based on TF binding motifs revealed complex interactions embedded within broader transcriptional networks involving multiple TFs. Both miRNAs were linked to extensive TF regulatory networks (Figure 6A). Although a subset of 26 TFs was shared, each miRNA also displayed unique TF associations, reflecting distinct layers of transcriptional control (Figure 6B). Interestingly, despite possessing a common seed sequence, these miRNAs appear to be regulated by different TFs and may therefore contribute to divergent biological roles in cancer, underscoring the complexity of their transcriptional regulation.
3. Discussion
Lung cancer patients are diagnosed in late stages, when the treatment strategies are related to metastatic disease, and the patients usually develop drug resistance [11,28,29,30,31]. In this study, we examined miR-28-5p and miR-708-5p, whose overexpression suggests potential oncogenic roles in lung cancer progression. Both miRNAs were significantly upregulated in tumor tissues compared with adjacent normal tissues (Figure 3), with further validation in the dbDEMC database. Notably, miR-28-5p displayed subtype-specific expression variability, indicating a possible role in tumor heterogeneity, whereas miR-708-5p showed consistent overexpression. ROC curve analysis reinforced these findings, as miR-708-5p demonstrated stronger discriminative ability between tumor and non-tumor tissues, underscoring its potential clinical utility in diagnosis.
MiR-28-5p expression is upregulated in NSCLC tumor tissues and cell lines [8,22]. In a recent study, the miR-28/PTEN axis is a crucial pathway that may serve as a potential clinical target for diagnosis, treatment, and prognosis [8]. Another study demonstrated that miR-28-5p can promote cancer progression by regulating the level of HIF-1α [22].
MiR-708-5p was identified as overexpressed in several lung cancer datasets [32], being associated with an increased risk of death after adjustments for all clinically significant factors, including age, sex, and tumor stage [33]. Overexpression of miR-708 in tumors is also related to poor overall survival, particularly in never-smoking lung adenocarcinoma patients [33]. Moreover, miR-708-5p has been associated with WNT regulation [33], β-catenin signaling [34], EMT, drug resistance and metastasis [35]. To our knowledge, no direct study that presents interaction with genes related to ECM, adhesion molecules, or Hippo signaling has been published. The oncogenic effects of miR-708 could be attributed to its interactions with specific target genes and pathways that regulate cell proliferation, survival, and metastasis [23,33,34,36]. The present study identified a significant number of genes targeted by the transcripts mentioned above, which appear to be involved in ECM–receptor interaction, including five that are correlated with the overall survival rate of NSCLC patients. A deeper comprehension of the structural and functional role of the ECM in NSCLC progression can be used to identify novel potential therapeutic targets, particularly in the case of high-risk groups [37]. These genes are known to be involved in cancer cell invasion [37,38].
Adherens junction proteins act as tumor suppressors or promoters of cell transformation [39]. The alteration of this class of genes has been widely reported in several solid tumors [40], which might have a different impact depending on the cancer type and stage [40]. Several of these genes were proved to be related to the differential infiltration of immune cells in the NSCLC [41].
MiRNAs were connected to the Hippo pathway in tumor progression [42], this finding is also supported by the present study, which highlights several target genes involved in this pathway; importantly, a substantial proportion of these genes show a correlation with overall survival. No direct information related to the connection of miR-28 and miR-708 and Hippo signaling was previously reported, even though the Hippo pathway has been reported to be closely associated with lung cancer [42].
A limitation of the present study is that our analysis related to target genes is confined to in silico assessments. Therefore, the clinical significance of our data has yet to be evaluated. Additional research is needed to better understand the system governing ECM, adherens, or Hippo signaling. Further studies, including functional experiments, mechanistic investigations, and analysis on larger patient cohorts, are needed to comprehensively understand the roles of miR-28-5p and miR-708-5p overexpression in NSCLC and to determine their potential as therapeutic targets or diagnostic/prognostic markers.
MiR-28-5p and miR-708-5p share a common seed sequence. However, their distinct biological properties may arise from differences in their target genes and cellular contexts, as well as their genomic localization. The seed sequence of miRNA is vital for target recognition and binding. Nevertheless, other factors, including target site accessibility, secondary structure, and interactions with other molecules, also influence the miRNA’s regulatory effects [43]. The presence of a shared seed sequence supports partial functional redundancy at the post-transcriptional level. Simultaneously, differences in TF regulation imply they could be activated under different cellular or pathological conditions, adding flexibility to cancer-related gene regulatory networks. Currently, there are no experimentally verified entries linking specific TFs to these transcripts. This gap indicates that the upstream transcriptional regulation of miR-28-5p and miR-708-5p remains underexplored. Understanding which TFs regulate their expression in NSCLC may help explain their different behaviors despite their shared seed sequence. This duality, with shared targets yet distinct regulation, emphasizes the complexity of miRNA–TF networks and may explain why these TE-derived miRNAs exhibit both overlapping and unique roles in tumor biology.
4. Materials and Methods
4.1. Genomic Localisation and Sequence Analysis
The genomic loci of miR-28-5p and miR-708-5p were identified using the MIRIAD database (https://www.miriad-database.org/miRNA/human/hsa-mir-708, accessed 12 August 2025). Pre-miR and the mature sequences of miR-28-5p and miR-708-5p were retrieved from miRbase [44], and their seed sequences (nucleotides 2–7/8) were identified and highlighted to assess sequence conservation. Sequences for mature hsa-miR-28-5p (accession number MIMAT0000085) and hsa-miR-708-5p (accession number MIMAT0004926) were retrieved from miRBase (http://mirbase.org, accessed 12 August 2025) and used as input to generate the secondary structures by RNAstructure, version 6.5 software [45].
4.2. Patients
This study included 32 patients diagnosed with NSCLC according to internationally accepted criteria. All the patients included in the study signed informed consent. For each patient, we stored fresh frozen tumors (TT) and their paired adjacent non-tumor tissue (TN) in a biobank. Table 3 summarizes the clinicopathological data for the NSCLC cases used for the present study.
4.3. Evaluation of miR-28-5p and miR-708-5p by Quantitative Real-Time PCR (RT-PCR)
Fresh frozen tissue was used for RNA extraction using the classical TriReagent-based method, as previously described [4]. For the evaluation of expression levels of the two transcripts, TaqMan MicroRNA Transcription kit (ThermoFischer Scientific, Waltham, MA, USA) and TaqMan microRNA primer assay (ThermoFischer Scientific, Waltham, MA, USA) were used, in parallel with the evaluation of U6 and RNU48 as housekeeping miRNAs as previously described [4]. Table 4 presents the sequence of the primers of the tested miRNAs. The data was analyzed using the ΔΔCT method and graphical representation. Statistical analysis of the relative expression levels in TT versus TN was done using GraphPad Prism software v.9 (GraphPad Software, San Diego, CA, USA).
4.4. miR-28-5p and miR-708-5p Expression Levels in Lung Cancer Public Database
Expression data for these two transcripts for lung cancer were retrieved from dbDEMC (Database of Differentially Expressed miRNAs in Human Cancers) [27,46], a curated resource that integrates high-throughput expression profiles from microarray and RNA-seq studies. dbDEMC provides systematically annotated information on miRNA expression across multiple cancer types, enabling comparison between tumor and normal samples [27,46].
4.5. Statistical Analysis
The differences between TT and TN were calculated using the t-test, with a significance of p < 0.05. Correlation analysis was performed employing the Pearson correlation coefficient. All analyses were conducted using GraphPad Prism software v.9 (GraphPad Software, San Diego, CA, USA). Additional graphical representation of receiving operator characteristics (ROC curve) analysis for the individual transcripts was done using GraphPad Prism software v.9, and the combined ROC curve for the two transcripts was performed using the CombiROC online tool (http://combiroc.eu/) [26].
4.6. Predicted Target Genes Enrichment Analysis
Target genes prediction for hsa-miR-28-5p and hsa-miR-708-5p were made using miRDB webtool [47]. For each miRNA, a gene set enrichment analysis of the predicted target genes was performed using ShinyGO version 0.80 [48]. Charts for gene ontology (GO) biological processes and molecular functions were generated using a false discovery rate (FDR) cutoff limit of 0.05, and data was sorted by FDR on X axis.
4.7. Biological Pathways Associated with miR-28b-5p and miR-708-5p
Predictions of potential targets of these transcripts were performed by computational algorithms based on ‘seed regions’ between miRNAs and target genes using TargetScan, 8.0 (http://www.targetscan.org, 20 August 2024) [49] and TFs TransmiR v3.0 (Scilab.cn, 22 August 2024) was used [50].
DIANA-miRPath v3.0 (http://www.microrna.gr/miRPathv3, accessed on 22 August 2024), an online software suite, was used for the assessment of miRNA regulatory roles and the identification of pathways modulated by the two miRNAs [49,51]. We were able to identify the main pathways regulated by the selected two transcripts. The interactions between miR-28-5p and miR-708-5p, as well as their related target genes, were identified using the miRNet online tool [52].
5. Conclusions
Our study highlights the oncogenic potential of miR-28-5p and miR-708-5p in lung cancer, as both were found to be overexpressed in tumor tissues compared with normal tissues, a finding validated using dbDEMC. While the two miRNAs share a common seed sequence and a relatively high number of target genes, they appear to be regulated by different TF. Notably, miR-28-5p displayed subtype-specific variability as observed from data downloaded from dbDEMC, potentially contributing to tumor heterogeneity, whereas miR-708-5p showed consistent expression patterns and stronger diagnostic performance, reinforcing its clinical utility.
While our findings provide comparative insights into two sequence-related miRNAs with distinct roles in lung cancer biology and clinical diagnosis, the study is limited by its reliance on in silico analyses. Future experimental validation will be necessary to confirm the regulatory mechanisms and strengthen their clinical application.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Sung H. Ferlay J. Siegel R.L. Laversanne M. Soerjomataram I. Jemal A. Bray F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries CA Cancer J. Clin.20217120924910.3322/caac.2166033538338 · doi ↗ · pubmed ↗
- 2Fu L. Wang H. Wei D. Wang B. Zhang C. Zhu T. Ma Z. Li Z. Wu Y. Yu G. The value of CEP 55 gene as a diagnostic biomarker and independent prognostic factor in LUAD and LUSCP Lo S ONE 202015 e 023328310.1371/journal.pone.023328332437446 PMC 7241791 · doi ↗ · pubmed ↗
- 3Zhang W. Zhang Q. Che L. Xie Z. Cai X. Gong L. Li Z. Liu D. Liu S. Using biological information to analyze potential mi RNA-m RNA regulatory networks in the plasma of patients with non-small cell lung cancer BMC Cancer 20222229910.1186/s 12885-022-09281-135313857 PMC 8939143 · doi ↗ · pubmed ↗
- 4Aghiorghiesei O. Zanoaga O. Raduly L. Aghiorghiesei A.I. Chiroi P. Trif A. Buiga R. Budisan L. Lucaciu O. Pop L.A. Dysregulation of mi R-21-5p, mi R-93-5p, mi R-200c-3p and mi R-205-5p in Oral Squamous Cell Carcinoma: A Potential Biomarkers Panel?Curr. Issues Mol. Biol.2022441754176710.3390/cimb 4404012135723379 PMC 9164081 · doi ↗ · pubmed ↗
- 5Osan C. Chira S. Nutu A.M. Braicu C. Baciut M. Korban S.S. Berindan-Neagoe I. The Connection between Micro RN As and Oral Cancer Pathogenesis: Emerging Biomarkers in Oral Cancer Management Genes 202112198910.3390/genes 1212198934946938 PMC 8700798 · doi ↗ · pubmed ↗
- 6Braicu C. Zimta A.A. Harangus A. Iurca I. Irimie A. Coza O. Berindan-Neagoe I. The Function of Non-Coding RN As in Lung Cancer Tumorigenesis Cancers 20191160510.3390/cancers 1105060531052265 PMC 6563001 · doi ↗ · pubmed ↗
- 7Braicu C. Calin G.A. Berindan-Neagoe I. Micro RN As and cancer therapy—From bystanders to major players Curr. Med. Chem.2013203561357310.2174/092986731132029000223834177 · doi ↗ · pubmed ↗
- 8Cui F. Zhou Q. Xiao K. Qian H. Micro RNA-28 promotes the proliferation of non-small-cell lung cancer cells by targeting PTEN Mol. Med. Rep.2020212589259610.3892/mmr.2020.1103332236614 · doi ↗ · pubmed ↗