Copper Homeostasis is influenced by Ics3 in Saccharomyces cerevisiae
Lucas Tang, Aseel AlKaabi, Damon Meyer

TL;DR
This study shows that the ICS3 gene in yeast helps maintain copper balance, which is important for cell growth.
Contribution
The study confirms ICS3's role in copper homeostasis using a new method and clarifies its specificity to copper.
Findings
ICS3 positively affects copper homeostasis in yeast cells.
ics3Δ mutants show no significant growth difference with zinc stress.
Mating types MATa and MATα do not affect sensitivity to copper or zinc.
Abstract
Saccharomyces cerevisiae has several unique open reading frames that contribute to overall fitness. This study examined ICS3 , a contributor of Cu 2+ homeostasis. Our results successfully replicate previous work by the Monteiro laboratory using a different method, confirming the positive effect of ICS3 on maintaining copper homeostasis for cell growth. No significant difference in growth of ics3Δ mutants was observed for Zn 2+ , another oxidative stressor. In addition, MATa and MATα mating types did not influence sensitivity to Cu 2+ or Zn 2+ .
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
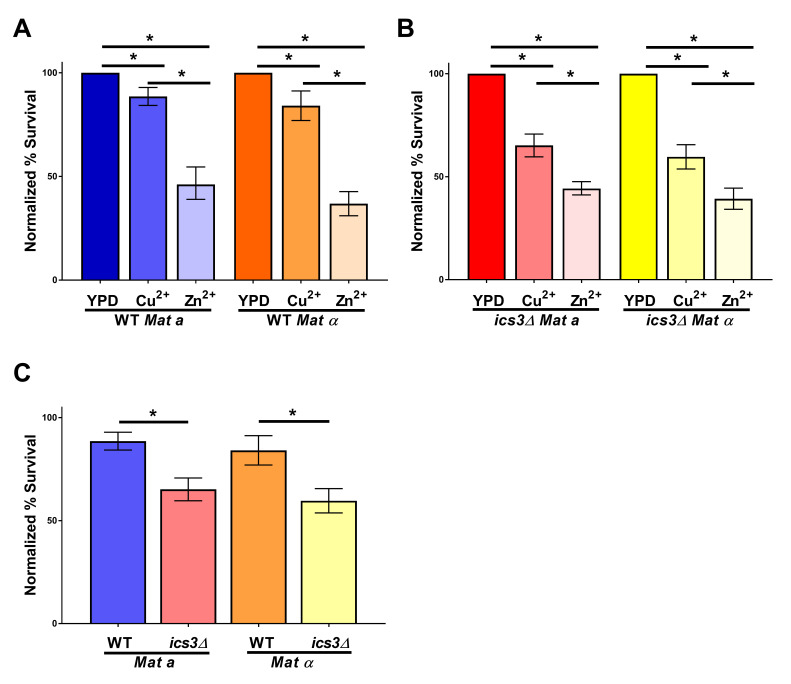 Figure 1
Figure 1|
Strain |
Genotype |
Source |
|
BY4741 |
|
California Northstate University, College of Health Sciences, Rancho Cordova, California, 95670. |
|
BY4742 |
|
The University of Pittsburgh, School of Medicine, Department of Computational and Systems Biology, Carvunis Lab |
|
yCHS90 |
|
The University of Pittsburgh, School of Medicine, Department of Computational and Systems Biology, Carvunis Lab |
|
yCHS91 |
|
The University of Pittsburgh, School of Medicine, Department of Computational and Systems Biology, Carvunis Lab |
|
Reagent |
Final Concentration |
Company |
|
Copper (II) Sulfate |
10 mM |
VWR |
|
Zinc (II) Sulfate |
10 mM |
VWR |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsTrace Elements in Health · Aluminum toxicity and tolerance in plants and animals · Fungal and yeast genetics research
Description
Saccharomyces cerevisiae is a model organism commonly used for genetic study due to ease of manipulation. In addition to the known 6,000 annotated open reading frames (ORFs) that are part of the canonical genome translated into proteins (Goffeau et al., 1996) , non-canonical, novel protein coding genes thought to have arisen through de novo gene birth have been identified as contributing to organism fitness (Carvunis et al., 2012; Wacholder et al., 2023). First named in a high throughput study by Entian et al. (1999) as an ORF of unclear function, *ICS3 * has since been associated with oxidative stress homeostasis, specifically with copper homeostasis (Alesso et al., 2015). Cu ^2+^ ions are necessary for cell growth in *S. cerevisiae * in trace amounts and can quickly become toxic as an oxidative stressor producing reactive oxygen species — demanding the need for tight cellular regulation of homeostasis (Culotta et al., 1995). Elucidating the role of an uncharacterized ORF in such a biochemical pathway can inform the role of non-canonical ORFs and de novo gene birth in organism fitness as proposed by Carvunis et al. (2012). This study follows up on the research by Alesso et al. by using a different methodology to evaluate cellular viability of wildtype and *ics3Δ * mutants in the presence of increasing amounts of copper and zinc.
Alesso et al. (2015) reported expression of *ICS3 * to be vital towards cell survival in the presence of excess copper. This was tested using a strain where *ICS3 * was deleted in the BY4741 background (Alesso et al., 2015). In the current study, a quantitative analysis of individual colony-forming units (CFUs) exposed to copper was performed in the same BY4741 background in an attempt to complement the findings of Alesso et al. (2015). The colony formation assay was used since it is a robust measure of individual cell reproductive potential while the serial dilution spot assay used by Alesso et al. (2015) provides an estimate of growth and survival that is less precise. Experiments in this study involved *MATa * and MAT𝛼 haploid wildtype and ics3Δ mutants exposed to 10mM of CuSO 4 in liquid YPD for four hours then plated onto solid YPD medium and incubated for 48-72 hours, after which individual colony counts were recorded. Since previous work indicated that pH influenced growth results in CuSO 4 , all media was adjusted to a pH of 6.0 (Alesso et al., 2015). Our results showed CuSO 4 had a significant effect on cell survival that was independent of mating type in both wildtype and ics3Δ mutants ( Figure 1A- C). This supports the previous results by Alesso et al. (2015), using another method of measurement, which confirms the role of Ics3 in regulating Cu ^2+^ ions. Therefore, the fitness of S. cerevisiae is impacted during acute exposure to environmental copper that can come from natural sources or human activity (Poggere et al., 2023).
In addition to our results with Cu ^2+^ , we wanted to explore if the effects observed are due to a general oxidative stress response to metal cations or is specific to copper homeostasis regulation. To address this, the same experiments were repeated in wildtype and *ics3Δ * mutants when exposed to ZnSO 4 , which produce Zn ^2+^ ions that are another cationic oxidative stressor similar to Cu ^2+^ . Zn ^2+^ is a bioessential metal for S. cerevisiae , playing a role in protein and membrane stabilization (White & Gadd, 1987), and as an essential catalytic component of many enzymes (Eide, 2003). Similarly to Cu ^2+^ , Zn ^2+^ becomes toxic to the cell in excess and is tightly regulated (Eide, 2003). Alesso et al. (2015) found negative results for the effects of Fe ^2+^ and Co ^2+^ as oxidative stressors on cell growth when ICS3 was deleted. This suggests that Ics3 may function more broadly to protect cells exposed to cationic oxidizers beyond just copper. To test this, the same protocol of a four hour exposure to the experimental stressor Zn ^2+^ was performed on *S. cerevisiae * cells in YPD with 10mM ZnSO 4 , followed by plating, and incubation for 48-72 hours before colony numbers were counted. Although overall growth was significantly decreased following the four hour exposure to zinc compared to when cells were only in YPD, this was observed for both the wildtype and *ics3Δ * mutants that were independent of mating type ( Figure 1A- C). Interestingly, a significant decrease in cell survival was observed in cells exposed to Zn ^2+ ^ compared to Cu ^2+^ across all genotypes examined ( Figure 1A- C).
While cell survival in both wildtype and *ics3Δ * mutants was affected by exposure to Cu ^2+^ and Zn ^2+^ , the specific contribution of Ics3 in protecting cells after Cu ^2+ ^ exposure required a direct comparison between wildtype and *ics3Δ * mutants. Analysis across both MATa and MATα , the same significant decrease in cell survival in *ics3Δ * mutants exposed to Cu ^2+ ^ when compared to wildtype was observed ( Figure 1C ). In contrast, exposure to Zn ^2+^ did not show a differential effect in wildtype compared to *ics3Δ * mutants ( Figure 1A- C).Therefore, it is likely the Ics3 protein does not contribute towards cell fitness in the presence of Zn ^2+^ and is more selective in which cationic oxidative stressors it regulates. Taken together, our results support Alesso et al’s (2015) findings by showing a specific role of Ics3 in protecting cells from the toxic effects of copper exposure.
The literature discussing the function of Ics3 in the cell has so far only considered Cu ^2+^ (Entian et al., 1999; Alesso et al., 2015), Co ^2+^ and Fe ^2+^ (Alesso et al., 2015). This study supports the role of Ics3 in Cu ^2+^ homeostasis, which is in alignment with previous studies, but also provides evidence that Ics3 does not play a role in Zn ^2+^ homeostasis, suggesting it can discriminate against specific cationic oxidizers. Information from the LoQAtE database, which provides protein localization information under various stress conditions, suggests that the Ics3 protein localizes to the mitochondria (Breker et al., 2013). However, additional experiments would be useful to specifically locate its position in the mitochondria, which has yet to be answered. This evidence that the Ics3 protein localizes to the mitochondria (Breker et al., 2013), combined with reports of the mitochondria regulating Mn ^2+^ homeostasis (Bleackley & MacGillivray, 2011; Reddit et al., 2009), indicate that further investigation is required to determine if Ics3 is truly limited to Cu ^2+^ homeostasis regulation. Potential future studies could include other bioavailable transition metal cations that are oxidative stressors and regulated via the mitochondria (Stoiber, 2010).
Methods
All strains used in this study were isogenic to the BY4741 or BY4742 background. Wildtype and ics3Δ mutants were grown in liquid YPD, liquid YPD supplemented with 10mM CuSO 4 , or liquid YPD supplemented with 10mM ZnSO 4 . Following the protocol described by Alesso et al. (2015) an overnight starter culture of wildtype or
- ics3Δ* mutant cells were diluted into fresh media (YPD, YPD with 10mM CuSO _4 _ , and YPD with 10mM ZnSO 4 ) to an initial OD 600 of 0.4. Cultures were incubated for four hours in liquid media with or without metal cation before making an appropriate dilution and plating to YPD. Cells were incubated for 48-72 hours until colonies were large enough to be counted. Survival on 10mM CuSO _4 _ or 10mM ZnSO 4 was normalized to growth on YPD by taking the total colony count from a metal exposed culture and dividing by the total colony count from YPD without metal and displayed as a percent (%). Statistical analysis was performed using Microsoft Excel and data visualization using GraphPad Prism.
Reagents
**: **
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Alesso C.A. Discola K.F. Monteiro G. 201591 The gene ICS 3 from the yeast Saccharomyces cerevisiae is involved in copper homeostasis dependent on extracellular p H Fungal Genetics and Biology 821087-1845435010.1016/j.fgb.2015.06.00726127016 · doi ↗ · pubmed ↗
- 2Bleackley Mark R. Mac Gillivray Ross T. A. 2011410 Transition metal homeostasis: from yeast to human disease Bio Metals 2450966-084478580910.1007/s 10534-011-9451-421479832 · doi ↗ · pubmed ↗
- 3Breker Michal Gymrek Melissa Schuldiner Maya 2013318 A novel single-cell screening platform reveals proteome plasticity during yeast stress responses Journal of Cell Biology 20061540-814083985010.1083/jcb.20130112023509072 PMC 3601363 · doi ↗ · pubmed ↗
- 4Carvunis Anne-Ruxandra Rolland Thomas Wapinski Ilan Calderwood Michael A. Yildirim Muhammed A. Simonis Nicolas Charloteaux Benoit Hidalgo César A. Barbette Justin Santhanam Balaji Brar Gloria A. Weissman Jonathan S. Regev Aviv Thierry-Mieg Nicolas Cusick Michael E. Vidal Marc 2012624 Proto-genes and de novo gene birth Nature 48774070028-083637037410.1038/nature 1118422722833 PMC 3401362 · doi ↗ · pubmed ↗
- 5Cizewski Culotta Valeria Joh Hung-Dong Lin Su-Ju Hudak Slekar Kimberly Strain Jeffrey 1995121 A Physiological Role for Saccharomyces cerevisiae Copper/Zinc Superoxide Dismutase in Copper Buffering Journal of Biological Chemistry 270500021-9258299912999710.1074/jbc.270.50.299918530401 · doi ↗ · pubmed ↗
- 6Eide David J. 200351 Multiple Regulatory Mechanisms Maintain Zinc Homeostasis in Saccharomyces cerevisiae The Journal of Nutrition 13350022-31661532 S 1535 S 10.1093/jn/133.5.1532 s 12730459 · doi ↗ · pubmed ↗
- 7Entian K.-D. Schuster T. Hegemann J. H. Becher D. Feldmann H. Güldener U. Götz R. Hansen M. Hollenberg C. P. Jansen G. Kramer W. Klein S. Kötter P. Kricke J. Launhardt H. Mannhaupt G. Maierl A. Meyer P. Mewes W. Munder T. Niedenthal R. K. Rad M. Ramezani Röhmer A. Römer A. Rose M. Schäfer B. Siegler M.-L. Vetter J. Wilhelm N. Wolf K. Zimmermann F. K. Zollner A. Hinnen A. 1999121 Functional analysis of 150 deletion mutants in Saccharomyces cerevisiae by a systematic approach Molecular and General Genetics MGG 2624-50026-89256837021 · doi ↗ · pubmed ↗
- 8Goffeau A. Barrell B. G. Bussey H. Davis R. W. Dujon B. Feldmann H. Galibert F. Hoheisel J. D. Jacq C. Johnston M. Louis E. J. Mewes H. W. Murakami Y. Philippsen P. Tettelin H. Oliver S. G. 19961025 Life with 6000 Genes Science 27452870036-807554656710.1126/science.274.5287.5468849441 · doi ↗ · pubmed ↗