Targeted Metabolomics of Tityus Scorpion Venoms: Unveiling Small-Molecule Components
Nathalia Baptista Dias, Bibiana Monson de Souza, Geovanny Barroso, Javier Ortiz Leiva, Gabriela Mendonça Paula, Hipócrates M. Chalkidis, Valquíria Abrão Coronado Dorce, Osmar Malaspina, Mario Sergio Palma

TL;DR
This study identifies small-molecule components in scorpion venoms that may play roles in prey capture and defense.
Contribution
A targeted metabolomic approach was used to identify 45 low molecular weight compounds in Tityus scorpion venoms.
Findings
45 compounds were reliably identified, including amino acids, organic acids, biogenic amines, and alkaloids.
Most compounds are neurotransmitters or neurotoxins, potentially aiding in prey paralysis and predator defense.
Some compounds may disrupt homeostasis or influence venom diffusion in victims.
Abstract
Scorpion venoms consist of proteins, peptides, and various low molecular weight (LMW) organic compounds, which act as toxins. Despite their potential significance, these compounds in scorpion venoms have been little investigated and their full range has not been well characterized. In this work, a targeted metabolomic approach was used in combination with an HPLC-QTOF-MS methodology to create a library of 55 LMW standard compounds, for the analysis of venoms from three Tityus species scorpions. This strategy enabled reliable identification of 45 compounds, including 20 amino acids, 4 organic acids, 12 biogenic amines, 6 nitrogenated bases and derivatives, 2 β-carboline-derived alkaloids, and 1 amphetamine. Most of the compounds identified were neurotransmitters and/or neurotoxins, while others can act as homeostasis disruptors or affect the diffusion of venom through the bodies of…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 1
1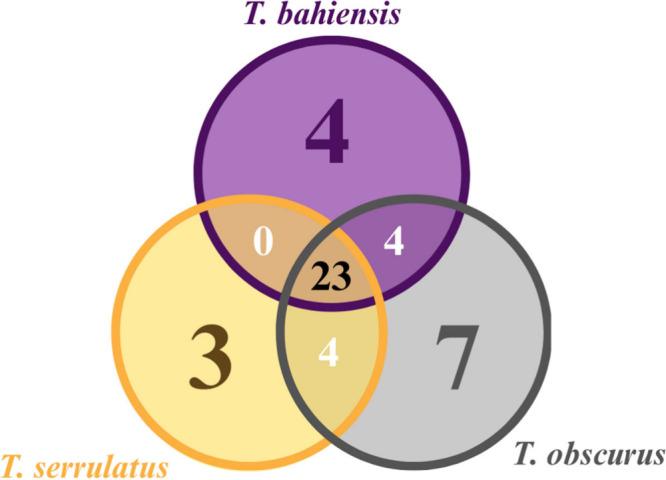 2
2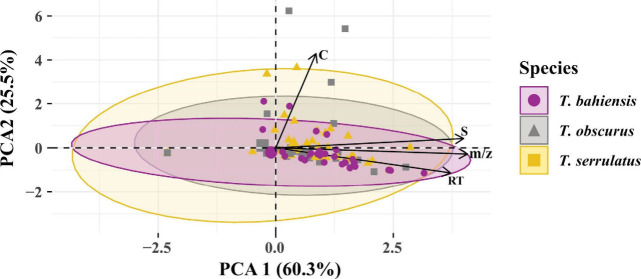 3
3| Main fragment-ions | |||||||
|---|---|---|---|---|---|---|---|
| No. |
| Rt (min) | Molecular Formula | Standard compounds |
|
|
|
|
| 75.0843 | 12.00 | C3H10N2 | 1.3-Diaminopropane | 58.06 | ||
|
| 122.0891 | 21.00 | C8H11N | 2-Phenylethylamine | 105.06 | 91.05 | |
|
| 134.0448 | 2.10 | C4H7NO4 | Aspartic acid | 116.03 | 117.03 | |
|
| 148.0604 | 2.80 | C5H9NO4 | Glutamic acid | 130.05 | 102.05 | 84.04 |
|
| 169.0422 | 9.00 | C8H8O4 | 3,4-Dihydroxyphenyl acetic acid | 123.04 | ||
|
| 152.0473 | 6.00 | C8H8O3 | 4-Hydroxyphenyl acetic acid | 135.03 | 152.05 | |
|
| 192.0582 | 11.50 | C10H9NO3 | 5-Hydroxyindoleacetic acid | 146.06 | 174.06 | |
|
| 176.0633 | 23.00 | C10H9NO2 | Indoleacetic acid | 130.06 | 158.87 | |
|
| 214.1001 | 14.50 | C10H15NO4 | Kainic acid | 196.09 | 168.10 | 150.09 |
|
| 117.0109 | 1.50 | C4H4O4 | Maleic acid | 99.01 | 134.04 | |
|
| 428.0294 | 1.50 | C10H15N5O10P2 | Adenosine diphosphate | 348.07 | 136.06 | |
|
| 136.0618 | 7.30 | C5H5N5 | Adenine | 119.04 | ||
|
| 330.0525 | 3.00 | C10H12N5O6P | 3′,5′-cyclic adenosine monophosphate | 136.06 | 177.06 | |
|
| 348.0704 | 1.50 | C10H14N5O7P | Adenosine monophosphate | 250.09 | ||
|
| 268.0967 | 3.60 | C10H13N5O4 | Adenosine | 136.06 | 136.06 | |
|
| 90.0550 | 3.00 | C3H7NO2 | Alanine | |||
|
| 175.1790 | 10.00 | C6H14N4O2 | Arginine | 158.09 | 129.11 | 116.07 |
|
| 133.0608 | 2.00 | C4H8N2O3 | Asparagine | 116.03 | 88.04 | |
|
| 118.0784 | 2.00 | C5H10NO2 | Betaine | 235.16 | ||
|
| 103.1230 | 13.20 | C5H14N2 | Cadaverine | 86.09 | ||
|
| 112.0432 | 4.50 | C4H5N3O | Cytosine | 95.02 | ||
|
| 154.0789 | 10.50 | C8H11NO2 | Dopamine | 137.06 | 123.04 | |
|
| 184.0895 | 7.00 | C9H13NO3 | Epinephrine | 166.09 | 135.08 | |
|
| 146.1652 | 22.00 | C7H19N3 | Spermidine | 129.14 | ||
|
| 203.2230 | 27.60 | C10H26N4 | Spermine | 129.14 | ||
|
| 166.0859 | 17.60 | C9H11NO2 | Phenylalanine | 120.08 | 149.06 | |
|
| 104.0706 | 4.50 | C4H9NO2 | GABA | 87.04 | 58.07 | |
|
| 76.0393 | 2.00 | C2H5NO2 | Glycine | |||
|
| 147.0764 | 2.30 | C5H10N2O3 | Glutamine | 130.05 | 101.07 | 84.04 |
|
| 152.0567 | 6.00 | C5H5N5O | Guanine | 135.03 | ||
|
| 284.0989 | 7.00 | C10H13N5O5 | Guanosine | 152.06 | 135.03 | |
|
| 364.0653 | 1.50 | C10H14N5O8P | Guanosine monophosphate | 152.06 | ||
|
| 444.0243 | 1.50 | C10H15N5O11P2 | Guanosine diphosphate | 152.06 | ||
|
| 288.1746 | 23.50 | C15H21N5O | Hydroxytrypargine | |||
|
| 112.0869 | 13.50 | C5H9N3 | Histamine | 95.06 | ||
|
| 156.0694 | 7.00 | C6H9N3O2 | Histidine | 139.06 | 95.06 | 81.05 |
|
| 132.0655 | 1.80 | C5H9NO3 | Hydroxyproline | 86.06 | ||
|
| 132.1019 | 14.00 | C6H13NO2 | Isoleucine | 102.06 | 86.10 | |
|
| 132.1019 | 14.50 | C6H13NO2 | Leucine | 86.09 | 89.00 | |
|
| 147.1055 | 8.50 | C6H14N2O2 | Lysine | 130.08 | 84.09 | |
|
| 150.0510 | 10.50 | C5H11NO2S | Methionine | 102.06 | 133.03 | 104.05 |
|
| 154.0863 | 7.00 | C8H11NO2 | Octopamine | 136.07 | 119.04 | |
|
| 163.1156 | 16.00 | C10H14N2 | Nicotine | 132.08 | 120.09 | 106.07 |
|
| 116.0706 | 3.00 | C5H9NO2 | Proline | |||
|
| 89.1073 | 12.00 | C4H12N2 | Putrescine | 72.08 | ||
|
| 106.0499 | 2.00 | C3H7NO3 | Serine | 88.04 | 60.04 | |
|
| 177.0949 | 15.00 | C10H12N2O | Serotonin | 160.07 | 159.09 | 146.06 |
|
| 126.0219 | 1.20 | C2H7NO3S | Taurine | |||
|
| 127.0429 | 3.00 | C5H6N2O2 | Thymine | 110.10 | ||
|
| 138.0840 | 13.00 | C8H11NO | Tyramine | 121.06 | 107.05 | |
|
| 182.0738 | 13.00 | C9H11NO3 | Tyrosine | 165.06 | 164.07 | 136.08 |
|
| 120.0582 | 13.00 | C4H9NO3 | Threonine | 103.04 | 85.04 | |
|
| 272.1796 | 31.00 | C15H21N5 | Trypargine | 213.14 | 255.16 | 196.11 |
|
| 205.0898 | 21.50 | C11H12N2O2 | Tryptophan | 188.07 | 159.09 | |
|
| 113.0346 | 1.50 | C4H4N2O2 | Uracil | 113.03 | 95.01 | |
|
|
|
|
|
|
|
|
|
|
| Concentration |
|---|---|---|---|---|---|---|---|---|---|---|
| Aspartic acid | C8H8O4 | 2.09 | 2.10 | –0.01 | 134.0501 | 134.0448 | 0.0053 | ++++ | 235 | 1.91 ± 0.40 |
| Kainic acid | C10H9NO3 | 14.67 | 14.50 | 0.17 | 214.1232 | 214.1001 | 0.0231 | ++ | 5873 | 11.62 ± 0.23 |
| Glutamic acid | C10H15NO4 | 2.06 | 2.80 | –0.74 | 148.0701 | 148.0604 | 0.0097 | ++++ | 2117 | 23.45 ± 0.47 |
| Indoleacetic acid | C4H7NO4 | 23.06 | 23.00 | 0.06 | 176.0910 | 176.0633 | 0.0277 | ++ | 13433 | 103.71 ± 2.60 |
| Adenine | C10H15N5O10P2 | 7.54 | 7.30 | 0.24 | 136.0599 | 136.0618 | –0.0019 | +++ | 283 | 0.38 ± 0.01 |
| Alanine | C3H7NO2 | 2.93 | 3.00 | –0.07 | 90.0710 | 90.0550 | 0.016 | +++ | 835 | 230.00 ± 4.60 |
| Arginine | C6H14N4O2 | 10.73 | 10.00 | 0.73 | 175.1802 | 175.1790 | 0.0012 | +++ | 141137 | 461.00 ± 9.23 |
| Betaine | C5H10NO2 | 2.03 | 2.00 | 0.03 | 118.0800 | 118.0784 | 0.0016 | ++++ | 7823 | 9.32 ± 0.19 |
| Cadaverine | C5H14N2 | 12.94 | 13.20 | –0.26 | 103.1245 | 103.1230 | 0.0015 | ++++ | 150 | 3.45 ± 0.07 |
| Cytosine | C4H5N3O | 4.65 | 4.50 | 0.15 | 112.0500 | 112.0432 | 0.0068 | ++++ | 2349 | 9.57 ± 0.19 |
| Dopamine | C8H11NO2 | 10.52 | 10.50 | 0.02 | 154.0732 | 154.0789 | –0.0057 | +++ | 1771 | 29.67 ± 0.59 |
| Epinephrine | C9H13NO3 | 7.10 | 7.00 | 0.10 | 184.1000 | 184.0895 | 0.0105 | +++ | 437 | 35.12 ± 0.70 |
| Spermidine | C7H19N3 | 22.16 | 22.00 | 0.16 | 146.1500 | 146.1652 | –0.0152 | ++++ | 6675 | 15.46 ± 0.31 |
| Spermine | C10H26N4 | 27.80 | 27.60 | 0.20 | 203.2245 | 203.2230 | 0.0015 | +++ | 407 | 1.22 ± 0.02 |
| Phenylalanine | C9H11NO2 | 17.70 | 17.60 | 0.10 | 166.0800 | 166.0859 | –0.0059 | ++++ | 2105 | 3.78 ± 0.08 |
| GABA | C4H9NO2 | 4.54 | 4.50 | 0.04 | 104.0901 | 104.0706 | 0.0195 | +++ | 466 | 3.50 ± 0.07 |
| Glycine | C2H5NO2 | 2.34 | 2.00 | 0.34 | 76.0396 | 76.0393 | 0.0003 | +++ | 474 | 843.00 ± 22.01 |
| Guanosine | C10H13N5O5 | 7.52 | 7.40 | 0.12 | 284.0900 | 284.0980 | 0.0080 | +++ | 393 | 2.90 ± 0.05 |
| Hydroxytrypargine | C15H21N5O | 23.61 | 23.50 | 0.11 | 288.1700 | 288.1746 | –0.0046 | ++++ | 2882 | 11.06 ± 0.23 |
| Histidine | C6H9N3O2 | 7.33 | 7.35 | 0.33 | 156.0828 | 156.0694 | 0.0134 | +++ | 19403 | 31.62 ± 0.63 |
| Isoleucine | C6H13NO2 | 14.01 | 14.00 | 0.01 | 132.0998 | 132.1019 | –0.0021 | ++++ | 15343 | 31.20 ± 0.62 |
| Leucine | C6H13NO2 | 14.34 | 14.50 | –0.51 | 132.1043 | 132.1019 | 0.0024 | ++++ | 75982 | 217.27 ± 4.36 |
| Lysine | C6H14N2O2 | 8.48 | 8.50 | –0.02 | 147.1103 | 147.1055 | 0.0048 | ++++ | 315662 | 761.00 ± 15.30 |
| Methionine | C5H11NO2S | 10.68 | 10.50 | 0.18 | 150.0654 | 150.0510 | 0.0144 | +++ | 5679 | 16.46 ± 0.31 |
| Octopamine | C8H11NO2 | 7.24 | 7.20 | –0.04 | 154.0813 | 154.0863 | –0.005 | ++++ | 1930 | 14.71 ± 0.29 |
| Proline | C5H9NO2 | 3.18 | 3.00 | 0.18 | 116.0810 | 116.0706 | 0.0104 | +++ | 5855 | 20.75 ± 0.43 |
| Tyramine | C8H11NO | 12.97 | 13.00 | –0.03 | 138.0740 | 138.0840 | –0.01 | ++ | 8612 | 48.80 ± 0.98 |
| Threonine | C4H9NO3 | 13.02 | 13.15 | 0.03 | 120.0700 | 120.0582 | 0.0118 | +++ | 919 | 46.24 ± 0.93 |
| Tryptophan | C11H12N2O2 | 21.90 | 21.50 | 0.40 | 205.1000 | 205.0898 | 0.0102 | +++ | 14865 | 46.84 ± 0.91 |
| Valine | C5H11NO2 | 8.76 | 8.75 | 0.01 | 118.0800 | 118.0863 | –0.0063 | ++++ | 5065 | 42.40 ± 0.85 |
|
|
|
|
|
|
|
|
|
|
| Concentration |
|---|---|---|---|---|---|---|---|---|---|---|
| 2-Phenylethylamine | C8H11N | 21.11 | 21.00 | 0.11 | 122.0904 | 122.0891 | 0.0013 | ++++ | 360 | 2.35 ± 0.04 |
| Kainic acid | C10H9NO3 | 14.67 | 14.50 | 0.17 | 214.1010 | 214.1001 | 0.0009 | ++++ | 6761 | 13.38 ± 0.25 |
| Glutamic acid | C10H15NO4 | 2.60 | 2.80 | –0.2 | 148.0611 | 148.0604 | 0.0007 | ++++ | 1222 | 13.53 ± 0.21 |
| Indoleacetic acid | C4H7NO4 | 23.08 | 23.00 | 0.08 | 176.0711 | 176.0633 | 0.0078 | +++ | 124 | 0.95 ± 0.02 |
| Maleic acid | C4H4O4 | 1.46 | 1.50 | –0.04 | 117.0134 | 117.0109 | 0.0025 | ++++ | 359 | 264.56 ± 5.34 |
| Adenosine | C5H5N5 | 3.75 | 3.60 | 0.15 | 268.0971 | 268.0967 | 0.0004 | ++++ | 2543 | 29.62 ± 0.56 |
| Alanine | C3H7NO2 | 2.93 | 3.00 | –0.07 | 90.0620 | 90.0550 | 0.0070 | +++ | 494 | 136.36 ± 2.86 |
| Arginine | C6H14N4O2 | 9.98 | 10.00 | –0.02 | 175.1801 | 175.1790 | 0.0011 | ++++ | 6939 | 22.68 ± 0.38 |
| Cytosine | C4H5N3O | 4.61 | 4.50 | 0.11 | 112.0500 | 112.0432 | 0.0068 | +++ | 843 | 3.43 ± 0.05 |
| Dopamine | C8H11NO2 | 10.72 | 10.50 | 0.22 | 154.0801 | 154.0789 | 0.0002 | ++++ | 886 | 14.84 ± 0.24 |
| Epinephrine | C9H13NO3 | 7.28 | 7.00 | 0.28 | 184.0923 | 184.0895 | 0.0028 | ++++ | 362 | 29.09 ± 0.25 |
| Spermine | C10H26N4 | 27.81 | 27.60 | 0.21 | 203.2241 | 203.2230 | 0.0011 | ++++ | 401 | 1.20 ± 0.02 |
| Phenylalanine | C9H11NO2 | 17.49 | 17.60 | –0.11 | 166.0899 | 166.0859 | 0.0040 | ++++ | 764 | 1.37 ± 0.02 |
| Glycine | C2H5NO2 | 2.34 | 2.00 | 0.34 | 76.0382 | 76.0393 | –0.0011 | +++ | 169 | 300.57 ± 5.78 |
| Glutamine | C5H10N2O3 | 2.31 | 2.30 | 0.01 | 147.0806 | 147.0764 | 0.0042 | ++++ | 341 | 3.16 ± 0.06 |
| Guanine | C5H5N5O | 6.19 | 6.00 | 0.19 | 152.0569 | 152.0567 | 0.2 | ++++ | 659 | 2.34 ± 0.05 |
| Guanosine | C10H13N5O 5 | 6.93 | 7.00 | –0.07 | 284.0945 | 284.0989 | –0.0044 | +++ | 1619 | 11.81 ± 0.18 |
| Hydroxytrypargine | C15H21N5O | 23.52 | 23.50 | 0.02 | 288.1711 | 288.1746 | –0.0035 | +++ | 445 | 1.70 ± 0.03 |
| Histidine | C5H9NO3 | 6.89 | 7.00 | –0.11 | 156.0728 | 156.0694 | 0.0034 | ++++ | 440 | 0.71 ± 0.01 |
| Isoleucine | C6H13NO2 | 14.27 | 14.00 | 0.27 | 132.1044 | 132.1019 | 0.0025 | ++++ | 9594 | 19.51 ± 0.35 |
| Leucine | C6H13NO2 | 14.58 | 14.50 | 0.08 | 132.0998 | 132.1019 | –0.0021 | +++ | 62903 | 179.86 ± 4.01 |
| Lysine | C6H14N2O2 | 8.56 | 8.50 | 0.06 | 147.1096 | 147.1055 | 0.0041 | ++++ | 7229 | 17.42 ± 0.28 |
| Methionine | C5H11NO2S | 10.68 | 10.35 | 0.33 | 150.0583 | 150.0510 | 0.0073 | +++ | 1223 | 3.54 ± 0.05 |
| Proline | C5H9NO2 | 3.18 | 3.00 | 0.18 | 116.0712 | 116.0706 | 0.6 | ++++ | 180 | 0.63 ± 0.01 |
| Serotonin | C10H12N2O | 15.00 | 15.00 | 0.00 | 177.1101 | 177.0949 | 0.0152 | +++ | 159 | 0.43 ± 0.01 |
| Tyramine | C8H11NO | 13.00 | 13.00 | 0.00 | 138.0900 | 138.0840 | 0.006 | +++ | 247 | 1.39 ± 0.02 |
| Tyrosine | C9H11NO3 | 13.00 | 13.15 | –0.05 | 182.0803 | 182.0738 | 0.0065 | +++ | 39163 | 158.64 ± 3.45 |
| Threonine | C4H9NO3 | 13.27 | 13.15 | 0.12 | 120.0601 | 120.0582 | 0.0019 | ++++ | 596 | 29.98 ± 0.60 |
| Trypargine | C15H21N5 | 31.00 | 31.00 | 0.00 | 272.1807 | 272.1796 | 0.0011 | ++++ | 1796 | 5.74 ± 0.10 |
| Tryptophan | C11H12N2O 2 | 21.61 | 21.50 | 0.11 | 205.0957 | 205.0898 | 0.0059 | +++ | 2023 | 6.37 ± 0.13 |
| Valine | C5H11NO2 | 8.78 | 8.75 | 0.03 | 118.0799 | 118.0863 | –0.0064 | +++ | 736 | 6.16 ± 0.15 |
|
|
|
|
|
|
|
|
|
|
| Concentration |
|---|---|---|---|---|---|---|---|---|---|---|
| 1,3-Diaminopropane | C3H10N2 | 12.31 | 12.00 | 0.31 | 75.0866 | 75.0843 | 0.0023 | ++++ | 1011 | 40.98 ± 0.09 |
| 2 Phenylethylamine | C8H11N | 20.62 | 21.00 | –0.38 | 122.0900 | 122.0891 | 0.9 | +++ | 7305 | 47.85 ± 0.07 |
| Aspartic acid | C4H7NO4 | 2.16 | 2.10 | 0.06 | 134.0400 | 134.0448 | –0.0048 | +++ | 492 | 4.00 ± 0.01 |
| Kainic acid | C10H15NO4 | 14.61 | 14.50 | 0.11 | 214.1200 | 214.1001 | 0.0199 | ++++ | 15303 | 30.30 ± 0.01 |
| Glutamic acid | C5H9NO4 | 2.78 | 2.80 | –0.02 | 148.0700 | 148.0604 | 0.0096 | ++++ | 1412 | 15.63 ± 0.03 |
| 4-Hydroxyphenylacetic acid | C8H8O3 | 5.79 | 6.00 | –0.21 | 153.0800 | 152.0473 | 1.0327 | +++ | 6201 | 27.64 ± 0.04 |
| 5-Hydroxyindoleacetic acid | C10H9NO3 | 11.39 | 11.50 | –0.11 | 192.0900 | 192.0582 | 0.0318 | +++ | 8317 | 58.60 ± 0.12 |
| Indoleacetic acid | C10H9NO2 | 23.12 | 23.00 | 0.12 | 177.1000 | 176.0633 | 1.0367 | ++++ | 2923 | 22.56 ± 0.04 |
| Maleic acid | C4H4O4 | 1.68 | 1.50 | 0.18 | 117.0670 | 117.0109 | 0.0561 | ++ | 3358 | 474.63 ± 7.35 |
| Alanine | C3H7NO2 | 3.10 | 3.00 | 0.1 | 90.0549 | 90.0550 | –0.1 | ++++ | 12682 | 500.70 ± 9.86 |
| Arginine | C6H14N4O2 | 10.31 | 10.00 | 0.31 | 175.1811 | 175.1790 | 0.0021 | ++++ | 19180 | 62.70 ± 1.36 |
| Asparagine | C4H8N2O3 | 2.20 | 2.00 | 0.2 | 133.0700 | 133.0608 | 0.0092 | ++ | 558 | 13.87 ± 0.02 |
| Cytosine | C4H5N3O | 4.48 | 4.50 | –0.02 | 112.0687 | 112.0432 | 0.0255 | +++ | 3379 | 13.76 ± 0.03 |
| Dopamine | C8H11NO2 | 10.49 | 10.50 | –0.01 | 154.0889 | 154.0789 | 0.0100 | ++++ | 8701 | 145.77 ± 2.89 |
| Epinephrine | C9H13NO3 | 6.89 | 7.00 | –0.11 | 184.1240 | 184.0895 | 0.0345 | ++ | 150 | 12.05 ± 0.02 |
| Spermidine | C7H19N3 | 22.11 | 22.00 | 0.11 | 146.1555 | 146.1652 | –0.0097 | ++++ | 5399 | 12.50 ± 0.01 |
| Spermine | C10H26N4 | 27.55 | 27.60 | –0.05 | 203.2218 | 203.2230 | –0.0012 | ++++ | 438 | 1.31 ± 0.01 |
| Phenylalanine | C9H11NO2 | 17.80 | 17.60 | 0.2 | 166.0880 | 166.0859 | 0.0021 | ++++ | 10855 | 19.49 ± 0.38 |
| GABA | C4H9NO2 | 4.29 | 4.50 | –0.21 | 104.0934 | 104.0706 | 0.0228 | ++++ | 632 | 4.74 ± 0.01 |
| Glycine | C2H5NO2 | 1.98 | 2.00 | –0.02 | 76.0401 | 76.0393 | 0.8 | ++++ | 1249 | 221.42 ± 4.38 |
| Guanosine | C10H13N5O5 | 7.15 | 7.00 | 0.15 | 284.0945 | 284.0989 | –0.0044 | ++++ | 807 | 5.88 ± 0.01 |
| Hydroxytrypargine | C15H21N5O | 23.19 | 23.50 | –0.31 | 288.1700 | 288.1728 | –0.0028 | ++++ | 33316 | 127.83 ± 2.27 |
| Histamine | C5H9N3 | 13.65 | 13.50 | 0.15 | 112.0850 | 112.0869 | –0.0019 | ++++ | 3651 | 10.98 ± 0.02 |
| Histidine | C6H9N3O2 | 7.13 | 7.00 | 0.13 | 156.0808 | 156.0694 | 0.0114 | ++++ | 52077 | 84.87 ± 016 |
| Isoleucine | C6H13NO2 | 14.21 | 14.00 | 0.21 | 132.0900 | 132.1019 | –0.0119 | ++++ | 9054 | 18.41 ± 0.35 |
| Leucine | C6H13NO2 | 14.52 | 14.50 | 0.02 | 132.1000 | 132.1019 | –0.0019 | ++++ | 7945 | 22.71 ± 0.03 |
| Lysine | C6H14N2O2 | 8.55 | 8.50 | 0.05 | 147.1100 | 147.1055 | 0.0045 | ++++ | 62893 | 151.63 ± 2.94 |
| Methionine | C5H11NO2S | 10.48 | 10.50 | –0.02 | 150.0600 | 150.0510 | 0.0090 | ++++ | 11281 | 32.69 ± 0.06 |
| Octopamine | C8H11NO2 | 7.03 | 7.00 | 0.03 | 154.0910 | 154.0863 | 0.0047 | +++ | 744 | 5.67 ± 0.01 |
| Proline | C5H9NO2 | 2.77 | 3.00 | –0.23 | 116.0900 | 116.0706 | 0.0194 | ++++ | 1786 | 6.33 ± 0.11 |
| Putrescine | C4H12N2 | 12.32 | 12.00 | 0.32 | 89.1000 | 89.1073 | –0.0073 | ++++ | 8318 | 88.32 ± 1.65 |
| Serotonin | C10H12N2O | 15.12 | 15.00 | 0.12 | 177.1100 | 177.0949 | 0.0151 | ++++ | 54698 | 150.46 ± 2.70 |
| Thymine | C5H6N2O2 | 2.93 | 3.20 | –0.27 | 127.0600 | 127.0429 | 0.0171 | ++++ | 24301 | 70.73 ± 1.37 |
| Tyramine | C8H11NO | 13.03 | 13.00 | 0.03 | 138.0710 | 138.0840 | –0.0130 | ++ | 7455 | 42.23 ± 0.85 |
| Tyrosine | C9H11NO3 | 13.11 | 13.00 | 0.11 | 182.0800 | 182.0738 | 0.0062 | ++++ | 4543 | 18.40 ± 0.34 |
| Threonine | C4H9NO3 | 12.98 | 13.15 | –0.17 | 120.0700 | 120.0582 | 0.0118 | ++++ | 1404 | 70.64 ± 1.45 |
| Tryptophan | C11H12N2O2 | 21.45 | 21.50 | –0.05 | 205.1045 | 205.0898 | 0.0147 | ++++ | 10460 | 32.95 ± 0.65 |
| Uridine | C9H13N2O9P | 1.56 | 1.50 | 0.06 | 352.0512 | 352.0431 | 0.0081 | ++ | 18760 | 167.91 ± 1.42 |
| Valine | C5H11NO2 | 8.70 | 8.75 | –0.05 | 118.0800 | 118.0863 | –0.0063 | ++++ | 25528 | 213.71 ± 4.21 |
|
| |||
|---|---|---|---|
|
| |||
|
|
|
|
|
| 1,3-Diaminopropane | - | - | 40.98 ± 0.09 |
| 2-Phenylethylamine | - | 2.35 ± 0.04 | 47.85 ± 0.07 |
| Adenine | 0.38 ± 0.01 | - | - |
| Adenosine | - | 29.62 ± 0.56 | - |
| Alanine | 230.00 ± 4.60 | 136.36 ± 2.86 | 500.70 ± 9.86 |
| Arginine | 461.00 ± 9.23 | 22.68 ± 0.38 | 62.70 ± 1.36 |
| Asparagine | - | - | 13.87 ± 0.02 |
| Aspartic Acid | 1.91 ± 0.40 | - | 4.00 ± 0.01 |
| Betaine | 9.32 ± 0.19 | - | - |
| Cadaverine | 3.45 ± 0.07 | - | - |
| Cytosine | 9.57 ± 0.19 | 3.43 ± 0.05 | 13.76 ± 0.03 |
| Dopamine | 29.67 ± 0.59 | 14.84 ± 0.24 | 145.77 ± 2.89 |
| Epinephrine | 35.12 ± 0.70 | 29.09 ± 0.25 | 12.05 ± 0.02 |
| GABA | 3.50 ± 0.07 | - | 4.74 ± 0.01 |
| Glutamic acid | 23.45 ± 0.47 | 13.53 ± 0.21 | 15.63 ± 0.03 |
| Glutamine | - | 3.16 ± 0.06 | - |
| Glycine | 843.00 ± 22.01 | 300.57 ± 5.78 | 221.42 ± 4.38 |
| Guanine | - | 2.34 ± 0.05 | - |
| Guanosine | 2.90 ± 0.05 | 11.81 ± 0.18 | 5.88 ± 0.01 |
| Histamine | - | - | 10.98 ± 0.02 |
| Histidine | 31.62 ± 0.63 | 0.71 ± 0.01 | 84.87 ± 016 |
| 5-Hydroxyindoleacetic acid | - | - | 58.60 ± 0.12 |
| 4-Hydroxyphenylacetic acid | - | - | 27.64 ± 0.04 |
| Hydroxytrypargine | 11.06 ± 0.23 | 1.70 ± 0.03 | 127.83 ± 2.27 |
| Indoleacetic acid | 103.71 ± 2.60 | 0.95 ± 0.02 | 22.56 ± 0.04 |
| Isoleucine | 31.20 ± 0.62 | 19.51 ± 0.35 | 18.41 ± 0.35 |
| Kainic acid | 11.62 ± 0.23 | 13.38 ± 0.25 | 30.30 ± 0.01 |
| Leucine | 217.27 ± 4.36 | 179.86 ± 4.01 | 22.71 ± 0.03 |
| Lysine | 761.00 ± 15.30 | 17.42 ± 0.28 | 151.63 ± 2.94 |
| Maleic acid | - | 264.56 ± 5.34 | 474.63 ± 7.35 |
| Methionine | 16.46 ± 0.31 | 3.54 ± 0.05 | 32.69 ± 0.06 |
| Octopamine | 14.71 ± 0.29 | - | 5.67 ± 0.01 |
| Phenylalanine | 3.78 ± 0.08 | 1.37 ± 0.02 | 19.49 ± 0.38 |
| Proline | 20.75 ± 0.43 | 0.63 ± 0.01 | 6.33 ± 0.11 |
| Putrescine | - | - | 88.32 ± 1.65 |
| Serotonin | - | 0.43 ± 0.01 | 150.46 ± 2.70 |
| Spermidine | 15.46 ± 0.31 | - | 12.50 ± 0.01 |
| Spermine | 1.22 ± 0.02 | 1.20 ± 0.02 | 1.31 ± 0.01 |
| Threonine | 46.24 ± 0.93 | 29.98 ± 0.60 | 70.64 ± 1.45 |
| Thymine | - | - | 70.73 ± 1.37 |
| Trypargine | - | 5.74 ± 0.10 | - |
| Tryptophan | 46.84 ± 0.91 | 6.37 ± 0.13 | 32.95 ± 0.65 |
| Tyramine | 48.80 ± 0.98 | 1.39 ± 0.02 | 42.23 ± 0.85 |
| Tyrosine | - | 158.64 ± 3.45 | 18.40 ± 0.34 |
| Valine | 42.40 ± 0.85 | 6.16 ± 0.15 | 213.71 ± 4.21 |
- —Fundação de Amparo à Pesquisa do Estado de São Paulo10.13039/501100001807
- —Fundação de Amparo à Pesquisa do Estado de São Paulo10.13039/501100001807
- —Coordenação de Aperfeiçoamento de Pessoal de Nível Superior10.13039/501100002322
- —Coordenação de Aperfeiçoamento de Pessoal de Nível Superior10.13039/501100002322
- —Coordenação de Aperfeiçoamento de Pessoal de Nível Superior10.13039/501100002322
- —Conselho Nacional de Desenvolvimento Científico e Tecnológico10.13039/501100003593
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsVenomous Animal Envenomation and Studies · Ion channel regulation and function · Nicotinic Acetylcholine Receptors Study
Introduction
Recent advances in omics technologies have greatly improved understanding of venom components.? These methodologies have revealed that venomous animals are an abundant source of diverse bioactive compounds including proteins, peptides, and many nonpeptidic organic toxins. The compounds in the last class (lipids, free amino acids, and organic acids), collectively known as low molecular weight (LMW) organic compounds, ?,? perform the general function of toxins. However, despite the pharmacological importance of these substances, sophisticated analytical platforms have been underutilized in venom studies. Consequently, the literature on animal venoms reports only a limited range of LMW compounds, ?,? revealing the continuing need to identify and characterize these components.
The metabolites produced by the venom gland tissue are secreted into the lumen of the gland, so they can be considered as extracellular components. In venomous arthropods, LMW organic toxins may also be biosynthesized by other glands and secreted into the hemolymph, followed by transport to the venom glands, where they are sequestered and stored.? These compounds are exometabolites that act in other organisms, after being injected into the victims of scorpion stinging. ?,?
The metabolome is the qualitative/quantitative complement of LMW metabolites of an organism or tissue, under specific physiological conditions.? If the complement of LMW toxins from animal venom is composed of extracellular metabolites, the chemical profile of these compounds can be considered a venom exometabolome.
Metabolomic studies of animal venom have adopted two different experimental strategies, namely targeted and untargeted approaches. Venom metabolic profiling has employed a targeted approach, focusing on molecules already identified as toxins, or those with potential pharmaceutical applications. However, few studies have undertaken holistic quantitative analyses of venom metabolite composition across various species, the parameters that influence it, or the similarities and differences in venom metabolic profiles among species.
In studies using targeted metabolomics, snake venoms have been investigated in the search for steroids? and polyamines. ?,? This strategy has also been used to investigate steroids in toad venoms. ?,? However, most of the research concerning animal venom metabolomics has used untargeted strategies. Examples are investigations of the presence of organic acids, nucleosides, amines, and polyamines in spider venoms, ?,? amino acids and amines in scorpion venoms,? amino acids, bufadienolides, and alkaloids in toad venoms,? kynurenic acid, amino acids, and sugars in frogs,? and amino acids, amines, and nucleosides in solitary and social wasps. ?,?
Annually, over 1.5 million scorpion sting cases are reported worldwide, with outcomes ranging from mild local reactions to severe health risks or death.? In Brazil, the Tityus genus is responsible for the most clinically significant scorpion envenomation, with Tityus serrulatus, T. bahiensis, and T. obscurus being responsible for the majority of human scorpion accidents.? Tityus serrulatus is the most dangerous scorpion species in Brazil, causing the most severe envenomation. It is widely distributed across several states, including São Paulo, Minas Gerais, and Rio de Janeiro.? T. bahiensis causes the most accidents in the Southeast, South, Midwest, and Bahia regions. In the Brazilian Amazon, T. obscurus is a primary species of medical concern.? In most cases, envenomation typically presents with local symptoms such as pain, swelling, and heat, as well as more general outcomes including headaches and sudoresis. Also common are digestive problems such as vomiting, neurological effects including tremors and dizziness, cardiovascular symptoms such as irregular heartbeats, and breathing difficulties.? The recommended treatment for scorpion stings is the administration of scorpion antivenom, derived from horses hyperimmunized with T. serrulatus venom. Symptomatic treatment focuses on pain relief, using either injection of 2% lidocaine, without a vasoconstrictor, at the sting site, or oral or parenteral administration of dipyrone or other analgesics.?
The search for bioactive molecules in animal venoms can contribute to the discovery of new candidates for pharmacological and industrial applications, in addition to providing a theoretical and technical basis for treating scorpionism. To improve these aspects, it is essential to understand the intra- and interspecific variability of scorpion venoms, as well as the influence of species distribution on their compositions.
The present work explores the metabolomic complexity of these venoms, identifying the metabolites associated with specific effects of scorpion sting envenomation. Mass spectrometry-based targeted metabolomics was used to identify and categorize metabolites in the venoms of three Brazilian Tityus species. Chromatographic separation of standard compounds, coupled with high-resolution mass spectrometry analysis, enabled the identification and quantification of metabolites.
Experimental
Section
Chemical Standards
The chemical standards used were as follows: 1,3-diaminopropane, 2-phenylethylamine, aspartic acid, glutamic acid, 3,4-dihydroxyphenylacetic acid, 4-hydroxyphenylacetic acid, 5-hydroxyindoleacetic acid, indoleacetic acid, malic acid, adenosine diphosphate, adenine, 3–5-cyclic adenosine monophosphate, adenosine 5-monophosphate, adenosine, alanine, arginine, asparagine, betaine, cadaverine, cytosine, dopamine, epinephrine, spermidine, spermine, phenylalanine, gamma-aminobutyric acid (GABA), glycine, glutamine, guanosine, guanine, guanosine monophosphate, guanosine, hydroxytrypargine, histamine, histidine, isoleucine, kainic acid, leucine, lysine, methionine, octopamine, proline, putrescine, serine, taurine, thymine, trypargine, tyramine, tyrosine, threonine, tryptophan, uracil, uridine, and valine. These compounds were obtained from Sigma-Aldrich (St. Louis, MO, USA).
Biological Materials
Three common Brazilian scorpion species were chosen for this study: T. serrulatus,? T. bahiensis,? and T. obscurus ? (all Scorpiones: Buthidae). T. serrulatus and T. bahiensis were reared and maintained at Butantan Institute (São Paulo, Brazil), and were supplied for the present study by the Department of Pharmacology of Butantan Institute. T. obscurus scorpions were collected in the region of Santarém (Amazonas state, in northern Brazil) by staff of Butantan Institute, with SISBIO/IBAMA authorization (protocol numbers 21483–2 and 20158–1). The animals were maintained in plastic boxes, with water provided ad libitum and regular feeding with cockroaches. Access to the genetic patrimony of this scorpion species was formally authorized by CGEN (protocol 010803/2013–0).
Venom Extraction
The venoms were obtained by electric stimulation of the telsons and collection using micropipettes, followed by lyophilization and storage at – 80 °C. Before analysis, the venom was solubilized in 50% (v/v) acetonitrile, filtered through an Amicon 3000 filter (Millipore), and centrifuged at 8,000g for 15 min at 4 °C. The pellets were discarded and the supernatants were lyophilized and stored at – 80 °C until further use. Three batches of crude venom from each species were processed to prepare three replicates.
Chromatographic and Mass
Spectrometric Analysis for Library Construction
Construction of the compound library began with optimization of the conditions for chromatographic separation, using an ultrafast liquid chromatograph (UFLC, Shimadzu) equipped with two LC-20AD pumps and an SIL-20AHT autosampler (Shimadzu). The analytes were separated on an XBridge BEH130 C18 column (2.1 × 100 mm, 3.5 μm) maintained at 38 °C. The mobile phases were (A) ultrapure water with 0.1% (v/v) heptafluorobutyric acid (HFBA), and (B) acetonitrile with 0.1% (v/v) HFBA. The gradient elution started at 2% (v/v) B and increased to 90% (v/v) B over 90 min. The eluent flow rate was 0.2 μL/min and the injection volume was 2 μL.
Mass spectrometric analysis employed a semi-μLC-ESI-microOToF-Q III instrument (Bruker Daltonics) coupled to the UFLC. The mass spectrometer was calibrated (in positive mode) using a 10 mM sodium formate solution, with calibration errors kept within 5 ppm and a calibration score of at least 90%. DataAnalysis v4.1 software (Bruker Daltonics) was used for data acquisition and processing.
The mass spectrometer was operated in both MS and MS/MS modes, with a scan range of m/z 50–650 and an acquisition rate of 1 Hz. The source parameters were set as follows: voltage of 4500 V, nebulizer gas pressure of 3 bar, drying gas flow rate of 8 L/min, drying gas temperature of 200 °C, and prepulse time of 4 μs. The collision energy was set at 8 eV for the MS mode and at 15 eV for the MS/MS mode. Broadband collision-induced dissociation (bbCID) was used to enable the simultaneous acquisition of MS and MS/MS data.
Quantification Methodology
The quantification employed QuantAnalysis v2.1 software (Bruker Daltonics), which allowed dynamic analysis of data from the total ion chromatogram (TIC) and facilitated the construction of calibration curves for the compounds. LC-MS/MS data were collected for each standard at four concentrations (3, 15, 69, and 125 ng/μL), with each concentration analyzed in triplicate. This approach enabled the generation of precise calibration curves for all the compounds, with calculation of the line equations and linear regression coefficients (R). Data management and further analyses were performed using Microsoft Excel (version 14).
Analysis of
LMW Compounds in Scorpion Venoms
The software packages used to construct the library and analyze data for the standard compounds and the scorpion venoms were Compass, QuantAnalysis v2.1, and DataAnalysis v4.1 (Bruker Daltonics). Compass was used for chromatographic and mass spectrometric data acquisition. DataAnalysis v4.1 was used to visualize and process the LC-MS data. This software was also used to determine the values of the parameters used to construct the library, including the retention times of the analytes, the m/z values of the protonated molecules of the compounds, the chromatographic peak areas, and the most important fragment ions generated for each compound in MS/MS mode. Additionally, high-resolution mass spectrometry data were used to determine the molecular formula of each compound, for both the library compounds and the venom samples.
After constructing a library of standard compounds, TargetAnalysis v1.2 software (Bruker Daltonics) was used for chromatographic fractionation, identification, and quantification of the compounds in the scorpion venoms. These analyses were performed using 30 μg of the venom extract from each scorpion species. The identification and validation of the compounds employed the exact m/z ratios (from high-resolution MS analysis), retention times, presence of qualifying ions (fragment ions), and isotopic patterns. TargetAnalysis software was used to determine the exact mass of each compound in the library, as a quasi-molecular [M + H]^+^ ion. This application also allowed the determination of mSigma, calculated based on the correlation between the theoretical monoisotopic pattern and that obtained experimentally for the ion of interest, providing additional validation by means of a score for the presence of the target compound. The deviations between the values for the validation parameters, such as retention time and m/z, as well as the presence of qualifying ions, provided a reliability score for the presence of the target compound. After completing the searches using TargetAnalysis, the results were exported to Microsoft Excel v14 (Microsoft, 2010) and organized in tables. The results obtained were manually checked by verifying the mass spectrometry and chromatography data using DataAnalysis. Subsequently, the concentration of each compound was calculated and expressed as μg of compound/μg of venom.
For the compound searches and identification using TargetAnalysis, the parameters were set as follows: 30 s retention time variation, 20 mDa tolerance between theoretical and expected m/z values for molecules in the monoprotonated form, signal-to-noise ratio limit of 5:1, narrow range mass tolerance value of 10 mDa, wide range mass tolerance of 20 mDa, narrow range mSigma value of 50, and wide range mSigma value of 1000. The initial construction of the metabolite library was performed considering a retention time interval limit among the points of the calibration curve (0.5 min), standardization of the peak area ratio with the concentration of each compound, and determination of the concentrations required to obtain sufficiently high signal intensities to construct reliable curves. The data set that made up the metabolite library was transferred to an Excel spreadsheet for storage in. csv format, including the following parameters: compound name, m/z of the molecule in the monoprotonated form, retention time, molecular formula, and main fragment ions.
Data processing with the TargetAnalysis software enabled wide- and narrow-range threshold values to be established for the analytical parameters mentioned above. These values were used to determine the detection score for each analyte. The analytical parameters outlined above were used to score and classify the identification of each analyte in the scorpion venoms.
Data Analysis
Pearson correlation analysis was used to elucidate the behaviors of the concentrations of the identified compounds, considering the three Tityus species and the data for the retention times, m/z values, and compound scores. A correlation matrix was constructed, with the coefficients obtained using the {rcorr} function in the “Hmisc” package? of RStudio v4.3.2 statistical software.? Subsequently, the data were fitted to a Gaussian model to identify any significant relationships between the variables. To assess the dispersion of the compounds isolated from the venoms of the Tityus species, principal component analysis (PCA) was performed using the “ade4” package. ?,? The “factominer”? and “factoextra”? packages were used to indicate explanatory and quantitative variables, respectively. Ellipses were plotted to show the separation of the different species for the identified compounds (95% confidence level). Species scores, representing the degree of correlation between the mean variables and the principal components, were added to represent each compound identified, for the distribution along the first two principal component axes. The graphical representation was obtained using the “ggplot2” package.?
Results and Discussion
The low molecular weight (LMW) standard compounds were separated using reversed-phase ion-pair chromatography (RPIP-HPLC), with ion-pair formation facilitated by the addition of heptafluorobutyric acid (HFBA) to both mobile phases (A and B). This modification significantly improved the retention of the analytes on the C18 column, consequently enhancing the chromatographic resolution. The formation of ion-pair compounds with HFBA under reversed-phase conditions enabled satisfactory chromatographic separation of the analytes, especially considering the compositional complexity of scorpion venom as an analytical matrix.
Fifty-five LMW standards were analyzed using RPIP-HPLC coupled with an ESI-microTOF-QIII mass spectrometer. Table provides comprehensive analytical data, including the high-resolution molecular masses of quasi-molecular ions, retention times, chemical formulas, compound names, and diagnostic fragment ions (m/z1, m/z2, and m/z3).
1: Standard Compounds Used to Build the Library of Low Molecular Mass Compounds, with Their Respective m/z Values as [M + H]+ Ions, Their Retention Time (Rt), Molecular Formula Obtained through HRMS, and Their Main Fragment-Ions Used in Their Identification (m/z 1, m/z 2, and m/z 3)
The standard compounds were quantified using a standardized calibration routine, as shown in the Supporting Information (Supplementary Figures S1–S16). The calibration curves exhibited excellent linearity, with R-values approaching 1.000, confirming the reliability and reproducibility of the method.
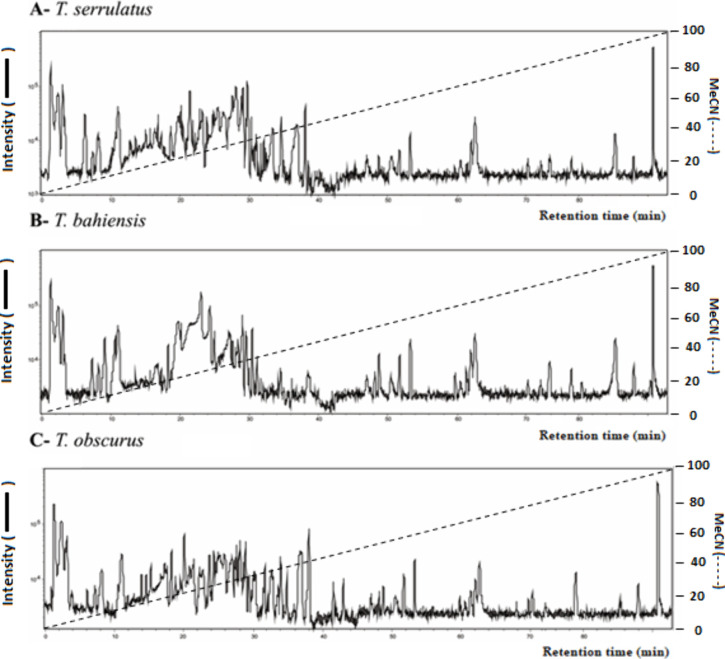
Following method standardization, venom samples from the three Tityus species (T. serrulatus, T. bahiensis, and T. obscurus) were fractionated and analyzed under optimized conditions. The total ion chromatograms (TICs) for each species are shown in Figure.
Total Ion Chromatogram (TIC) of the extracts containing the low molecular mass compounds from scorpion venoms, under RP-LCMS using a XBridge BEH130 C18 column (2,1 × 100 mm; 3,5 μm), with a gradient from the mobile phases (A) [H2O containing 0.1% (v/v) HFBA] to (B) [MeCN containing 0.1% (v/v) HFBA] in 90 min, at a flow rate of 0.2 mL/min under at 38 °C. The TICs above correspond to (A) T. serrulatus, (B) T. bahiensis, and (C) T. obscurus.
Tables–? show the analytical parameters, including the retention times for the venom samples and standards, mass errors, identification scores, peak areas, and estimated concentrations. These data provide a comprehensive overview of the LMW compound compositions of the venom samples.
2: Main Low Molecular Weight Compounds Identified in the Venom of Tityus serrulatus by LC-ESI-microTOF-QIII [WATERS XBridgeBEH130 C 18 column (2.1 × 10 mm, 3.5 μm)] from the Comparison with the Metabolite Library Using the Standard Chromatographic Conditions [Containing HFBA 0.1% (v/v)]
3: Main Low Molecular Weight Compounds Identified in the Venom of Tityus bahiensis by LC-ESI-microTOF-QIII [WATERS XBridgeBEH130 C 18 column (2.1 × 10 mm, 3.5 μm)] from the Comparison with the Metabolite Library Using the Standard Chromatographic Conditions [Containing HFBA 0.1% (v/v)]
4: Main Low Molecular Weight Compounds Identified in the Venom of Tityus obscurus by LC-ESI-microTOF-QIII [WATERS XBridgeBEH130 C 18 column (2.1 × 10 mm, 3.5 μm)] from the Comparison with the Metabolite Library Using the Standard Chromatographic Conditions [Containing HFBA 0.1% (v/v)]
To develop the data processing method using the TargetAnalysis software, it was necessary to establish both wide-range and narrow-range threshold values for the parameters mentioned above. These values were used to assign the scores for the detection and identification of the compounds, enabling classification of the parameters. Tables–? show the following analytical parameters for the compounds: experimental retention times for the metabolites in the scorpion venoms, retention times for the standards, differences in retention times between the venom compounds and standards, m/z of the compounds detected as [M + H]^+^ ions for the venoms and standards, errors of the molecular masses, identification scores, peak areas of the compounds, and concentrations of the compounds detected in the scorpion venoms. All the compounds assigned to the venoms were reliably identified, presenting scores from ++ to ++++.
A library of LMW organic compounds was constructed using 55 standard reference compounds. From this library, 45 compounds were reliably identified in the venoms of the Tityus scorpions. Among the compounds identified, 20 were amino acids, 4 were organic acids, 12 were biogenic amines, 6 were nitrogenated bases and/or their derivatives, 2 were alkaloids, and 1 was an amphetamine. The global distribution profile of these compounds across the venoms of the three scorpion species provided insights into the biochemical diversity and specificity of the venom compositions (Table).
5: Summary of Compounds Detected, Identified and Quantified in the Venom of Each Scorpion Species
The distribution of LMW toxins among the three Tityus species revealed both significant overlap and species-specific profiles. As shown in the Venn diagram (Figure), the three species shared 23 compounds, including key amino acids (glutamic, aspartic, and kainic acids), biogenic amines (dopamine and epinephrine), and nitrogenated bases (cytosine and guanosine).
Venn diagram of the low molecular mass compounds detected in the venoms from the scorpions T. serrulatus, T. bahiensis and T. obscurus.
Notably, four compounds were shared between T. serrulatus and T. obscurus (GABA, spermidine, octopamine, and aspartic acid). Another four compounds were common to T. bahiensis and T. obscurus (tyrosine, maleic acid, serotonin, and 2-phenylethylamine). However, no compound was found to be shared between T. serrulatus and T. bahiensis. Unique compounds were identified for each species, with T. serrulatus presenting betaine, cadaverine, and adenine, while T. bahiensis presented glutamine, trypargine, adenosine, and guanine, and T. obscurus presented asparagine, hydroxyphenylacetic acid, hydroxyindoleacetic acid, 1–3 diaminopropane, histamine, putrescine, and thymine.
Several compounds reported previously in other animal venoms, such as taurine, serine, and AMP-related derivatives, were not reliably identified in the Tityus venoms, suggesting unique venom profiles and potential evolutionary specialization. These findings highlighted the conserved biochemical elements and distinct molecular signatures of the scorpion species.
The PCA results provided insights into the distribution of venom compounds among the Tityus species. The first principal component (PC1), accounting for 60.3% of the variance, was the primary axis determining compound differentiation among the species. The second principal component (PC2) explained 25.5% of the variance, contributing to the structure of the compound distribution (Figure). The compound retention time, m/z, and score were significantly correlated (Gaussian model: df = 3; F = 4.122; p = 0.0078), suggesting that these parameters were important in determining compound variability. However, the compound concentrations showed weak correlations with these parameters, implying that concentration alone did not strongly influence compound classification.
Principal component analysis (PCA) and spatial distribution of proteomics analysis of venom from scorpion species. Plots analyzed Tityus species based on the concentrations, retention time, m/z and score of compounds identified. The species close to each other contain a similar composition of substances, while species with the biggest distance differ in substances identified. The ellipses were described with a 95% confidence level of the sampled venoms, and the arrows show the quantitative parameters identified along the first two principal component axes. RT: Retention Time; m / z: mass-to-charge ratio; S: Score; and C: Concentration.
The species T. bahiensis and T. serrulatus showed greater similarity in venom compound distribution, as indicated by the clustering of the points, reflecting shared chemical features of the venom compositions. T. obscurus appeared to be more distinct, with the corresponding data points distributed differently across the PCA component plot. The larger elliptical spread for T. serrulatus could be attributed to variations in the compound concentrations, as suggested by the influence of the concentration vector. This variability could have been due to intraspecific differences in venom composition, as well as the effects of environmental and physiological factors on production of the compounds.
The LMW organic compounds found in scorpion venom are metabolites secreted into the lumen of the venom gland, becoming exometabolites that do not act in the classical metabolic steady-state. However, these compounds remain active, playing an important role in the envenomation mechanism, with various pharmacological and physiological actions.
Amino acids found in the venoms of the three scorpion species included glutamic acid, aspartic acid, histidine, arginine, phenylalanine, alanine, glycine, glutamine, isoleucine, leucine, lysine, methionine, asparagine, tyrosine, threonine, tryptophan, proline, and valine. These amino acids are essential components of proteins, although some are also neurotransmitters, such as aspartic acid, glycine, and glutamic acid. ?,? In addition to being a proteinogenic amino acid, glutamic acid is the most abundant and important excitatory neurotransmitter in the central nervous system (CNS).? This amino acid was observed in the venoms of the three scorpion species studied in the present work.
Gamma-aminobutyric acid (GABA), which was detected in the venoms of T. serrulatus and T. obscurus, is a nonproteinogenic amino acid present at high concentrations in the CNS, where it functions as an inhibitory neurotransmitter and blocks CNS signals. GABA performs a series of functions in neuronal biochemistry and postsynaptic regulation.? A decrease in GABAergic neurotransmission in the CNS contributes to hyperexcitability.? GABA activates the enzymatic processes of transamination and decarboxylation in the Krebs cycle, being synthesized from glutamate by the l-glutamic acid decarboxylase enzyme, with vitamin B6 as cofactor. This process converts the primary excitatory neurotransmitter (glutamate) into one of the main inhibitory neurotransmitters.?
Kainic acid was detected in the venoms of all three scorpion species studied. Kainic acid is an agonist of the kainate subtype of ionotropic glutamate receptors, which directly gate ion channels and are generally excitatory. Studies using experimental models have shown that the administration of kainic acid can result in convulsions,? changes in rodent behavior,? oxidative stress,? glial activation,? production of inflammatory mediators,? endoplasmic reticulum stress, mitochondrial dysfunction, and selective neuronal degeneration in the rodent brain.?
Betaine (N,N,N-trimethylglycine), present in the T. serrulatus venom, is an essential intracellular osmolyte that regulates cell volume by counterbalancing changes in extracellular tonicity and stabilizing macromolecules against various physiological disturbances. ?,? Betaine also plays a key role in the metabolism of methyl radicals, providing cells with a supply of methyl groups. ?,? Betaine can be obtained from the diet or be produced in the body by the oxidation of choline.?
The polyamines found in the scorpion venoms included diaminopropane, putrescine, spermidine, cadaverine, and spermine. These compounds are widely distributed in animal tissues, where they play fundamental roles in the action of hormones, control of cell division, and synthesis of macromolecules.? Polyamines have also been reported to influence cellular excitability and neurotransmission by interacting with potassium ion channels and AMPA- and NMDA-dependent glutamate receptors. ?,?
Serotonin was identified in the T. bahiensis and T. obscurus venoms. This compound does not generally appear to alter the toxicity of venoms, but can contribute to the occurrence of pain and inflammation.?
Dopamine, which was identified in the venoms of all three scorpion species, is a monoamine neurotransmitter acting in the communication of messages between nerve cells in the brain and the rest of the body, leading to an increased heart rate that could assist rapid circulation of venom components in the body of the prey.?
Histamine, identified in the venoms of T. bahiensis and T. serrulatus, has been reported previously in other scorpion venoms. This pro-inflammatory compound has a vasodilatory effect on blood vessels, increasing vascular permeability and reducing blood pressure.?
Epinephrine, which was observed in the venoms of the three scorpion species, is a hormone that also acts as a CNS neurotransmitter. It participates in the sympathetic nervous system and is involved in “fight-or-flight” responses to emergency situations.?
Spermine has been reported in the venoms of spiders? and snakes, ?,? while putrescine, spermidine, and cadaverine have been observed in the venoms of four species of tarantulas.? Spermine causes nephrotoxicity,? breathing difficulty, hypotension, and diuresis.? Many of these effects are also exhibited by spermidine, but at higher concentrations of the compound. In mice, spermidine was reported to cause other toxicity symptoms including head and limb tremors, reduced muscle tone, paralysis of the hind limbs, decreased reflexes, hypothermia, and peripheral vasoconstriction,? while in rats, it led to histopathological renal changes.?
Octopamine was observed in the venoms of T. serrulatus and T. obscurus, while tyramine, an invertebrate neurotransmitter analogous to vertebrate adrenergic transmitters, was detected in the venoms of all three scorpion species. The decarboxylation of tyrosine produces these compounds, with tyramine being the biological precursor of octopamine. Both compounds are neurotransmitters that act by coupling to G-protein receptors. They can be stored at high concentrations in synaptic vesicles, presenting agonist activity on adrenergic receptors in mammals, and can cause sudden hypotension. Octopamine regulates several behaviors and sense organs in insects, enabling appropriate responses to external stimuli. This compound has attracted particular attention, because it is the only biogenic amine acting exclusively in invertebrates, so its receptors are promising targets for new insecticides.? Octopamine and tyramine in animal venoms may cause hyperexcitation in the prey, prior to paralysis.?
The venom of T. obscurus contained 1,3-diaminopropane, which is a compound that acts as a building block in the biosynthesis of acylpolyamine toxins of orb-web spiders.? However, no information is available regarding its physiological and pharmacological effects as a free metabolite in venom.
This is the first study to detect guanosine and cytosine in scorpion venom. Adenosine, which was previously reported in the venom of the scorpion Heterometrus laoticus, was reliably identified in the venom of T. bahiensis. Adenosine is known to be a potent inhibitor of platelet aggregation, consequently interfering in blood coagulation.? Previous work has evidenced the presence of compounds such as guanosine, inosine, and 2,4,6-trihydroxypurine in spider venoms.? Sulfated guanosine derivatives present in brown spider venoms have been found to act as potent neurotoxins.?
5-hydroxyindoleacetic and 4-hydroxyphenylacetic acids, which were detected in the T. obscurus venom, are used in venomous animals as chromophores in the biosynthesis of acylpolyamines.? No pharmacological roles are known for these compounds in animal venoms. Indoleacetic acid was observed in the venoms of the three Tityus species. In animal models, the administration of indoleacetic acid was found to cause an acute hypoglycemic response, ?,? accompanied by irritability, weakness, myotonia,? lassitude, and immobility.? Maleic acid was found in the venoms of T. bahiensis and T. obscurus. This is the first report of maleate in animal venom and it is not known whether this compound is active in scorpion envenomation incidents.
The detected LMW compounds included some beta-carboline alkaloids, such as hydroxytrypargine in the venoms of all three scorpion species and trypargine in the venom of T. bahiensis. These alkaloid toxins were previously observed in the venoms of the spiders Nephyla clavipes and Parawixia bistriata, ?,? acting as potent neurotoxins causing the death or paralysis of prey.
The amphetamine 2-phenylethylamine was identified in the venoms of T. bahiensis and T. obscurus. This compound can induce depression and anxiety, acting on the motor activity of animals, with changes in the frequency parameters of locomotion and body lifting.?
Venomous animals typically use their venoms to capture prey, deter predators, facilitate parasitism, and perform extra-oral digestion.? The richness of LMW compounds in animal venom provides many tools for these purposes. It is likely that natural selection has led to the use of these compounds as toxins in animal venoms, due to their capacity to affect neuronal systems. Some of them can induce pain or discomfort in predators (such as vertebrates), targeting monoaminergic systems, apparently for the purpose of defense.? Another potential function of LMW compounds in scorpion venoms is to accelerate the distribution of venom components throughout the body of the victim. For example, histamine and serotonin (which induces the release of histamine) cause vasodilation at the injection site, thereby facilitating the circulation of compounds throughout the victims.? Dopamine has been reported in Hymenoptera venoms, acting to increase the heart rate in the victims of stinging.? Several of the LMW organic compounds found in venoms can activate the sympathetic nervous system or adrenergic receptors in vertebrates, increasing the heart rate.
Another aspect to be considered is the purpose of compromising the ability of the prey to develop an effective escape response. The presence of many neurotransmitters and ion channel blockers may be related to the occurrence of intense physiological stress, which generally results in paralysis and/or death of the prey. Paradoxically, this is often achieved by activation of the sympathetic nervous system, such as in the catecholamine cascade and other actions involving epinephrine, often causing cardiac collapse, as demonstrated in the case of spider venoms.? The hyperexcitation induced by the venom immobilizes the prey until the occurrence of flaccid paralysis, characterized by weakness or reduced muscle tone. Hyperarousal in the prey is a common response to the venoms of various animals, such as spiders, scorpions, coelenterates, and some snails, associated with the presence of monoamines in the venoms. ?−? ? ? Hyperexcitation results in immediate immobilization of the prey, so that it remains within reach of the predator until flaccid paralysis begins. It has been found that dopamine in the venom of the Amulex compressa wasp induces excessive grooming in cockroaches, preventing their escape,? so an analogous mechanism may occur in the case of scorpion venom.
Conclusions
Generally venomous animals use their venoms as instruments for prey capture, and for deterring predators, facilitating parasitism, and to perform extra-oral digestion. The richness of LMW compounds in animal venoms certainly provide many tools for the above-mentioned purposes. Many of these compounds act individually at physiologic/pharmacological level, while others may interact with the proteins/peptides (specially neurotoxins and their receptors), potentiating their effects.
One of the reasons why natural selection chosen these compounds to act as toxins in animal venoms is probably related to their capacity of affecting neuronal systems, as well because some of them can produce pain or discomfort in predators (e.g., vertebrates), targeting monoaminergic systems, apparently with a defensive purpose. Another potential function for LMW compounds from scorpion venoms is to accelerate the distribution of venom components throughout the victim’s body. Some LMW organic compounds from venoms can activate the proteins of the sympathetic nervous system and the adrenergic receptors, thereby increasing heart rate in vertebrates.
Another point to be taken into consideration is the purpose of compromising the prey’s ability to organize an effective escape response. The presence of a large number of different types of neurotransmitters and ion channel blockers may be related to the occurrence of intense physiological stress, which generally results in paralysis and/or death of prey. Paradoxically, this is often accomplished with activation of the sympathetic nervous system, as in the catecholamine cascade and other actions involving epinephrine, often causing cardiac collapse. The hyperexcitation induced by the venoms immobilizes the prey until the occurrence of flaccid paralysis, which is characterized by weakness or reduced muscle tone. Hyperarousal is a common strategy perceived as responses to various types of animal venoms, produced by monoamines present in these venoms.
Scorpion venoms are extremely versatile tools, being effective against prey (insects) and in defense against vertebrate predators. These venoms have drastic effects in mammals, and an interesting fact to speculate is why the venoms evolved this way, since these animals normally target insects as prey. The action in mammals is possibly related to the effect caused by the synergistic actions at the pharmacological level, of the various biologically active LMW organic compounds present in the venoms, causing considerable physical discomfort in mammals’ victims of stings of these animals, with a powerful defensive effect. In addition to this, the high number of compounds acting at the level of the nervous and homeostatic systems, can cause an intense physiological stress that results in the paralysis or death of scorpions’ prey.
Supplementary Material
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Xia Z.He D.Wu Y.Kwok H. F.Cao Z.Scorpion venom peptides: Molecular diversity, structural characteristics, and therapeutic use from channelopathies to viral infections and cancers Pharmacol. Res.202319710697810.1016/j.phrs.2023.10697837923027 · doi ↗ · pubmed ↗
- 2Palma, M. S. Hymenoptera Venom Peptides. In Handbook of Biologically Active Peptides; Kastin, A. ; Ed.; Academic Press, San Diego, USA, 2013; pp 416–422.
- 3Pucca M. B.Cerni F. A.Pinheiro Junior E. L.Bordon K. d. C. F.Amorim F. G.Cordeiro F. A.Longhim H. T.Cremonez C. M.Oliveira G. H.Arantes E. C. Tityus serrulatus venom - A lethal cocktail Toxicon.201510827228410.1016/j.toxicon.2015.10.01526522893 · doi ↗ · pubmed ↗
- 4Klupczynska A.Plewa S.Derezinski P.łGarrett T. J.Rubio V. Y.Kokot Z. J.Matysiak J.Identification and quantification of honeybee venom constituents by multiplatform metabolomics Sci. Rep.2020102164510.1038/s 41598-020-78740-133303913 PMC 7729905 · doi ↗ · pubmed ↗
- 5Alvarado-Gonzalez C.Clement H.Ballinas-Casarrubias L.Escarcega-Avila A.Arenas-Sosa I.Lopez-Contreras K. S.Zamudio F.Corzo G.Espino-Solis G. P.Identification and Venom Characterization of Two Scorpions from the State of Chihuahua Mexico: Chihuahuanus coahuliae and Chihuahuanus crassimannus Toxins (Basel)202315741610.3390/toxins 1507041637505685 PMC 10467103 · doi ↗ · pubmed ↗
- 6Salabi F.Vazirianzadeh B.Baradaran M.Identification, classification, and characterization of alpha and beta subunits of LVP 1 protein from the venom gland of four Iranian scorpion species Sci. Rep.2023132227710.1038/s 41598-023-49556-638097679 PMC 10721645 · doi ↗ · pubmed ↗
- 7Bar-On B.On the form and bio-mechanics of venom-injection elements Acta Biomater.20198526327110.1016/j.actbio.2018.12.03030583109 · doi ↗ · pubmed ↗
- 8Nencioni A. L. A.Neto E. B.de Freitas L. A.Dorce V. A. C.Effects of Brazilian scorpion venoms on the central nervous system J. Venom. Anim. Toxins. Incl. Trop. Dis.201824310.1186/s 40409-018-0139-x 29410679 PMC 5781280 · doi ↗ · pubmed ↗